
Abstract
In utero injection of a retroviral vector into the embryonic telencephalon aided by ultrasound backscatter microscopy permits introduction of a gene of interest at an early stage of development. In this study, we compared the tissue distribution of gene expression in adult mice injected with retroviral vectors at different embryonic ages in utero. Following ultrasound image-guided gene delivery (UIGD) into the embryonic telencephalon, adult mice were subjected to whole-body luciferase imaging and immunohistochemical analysis at 6 weeks and 1 year postinjection. Luciferase activity was observed in a wide range of tissues in animals injected at embryonic age 9.5 (E9.5), whereas animals injected at E10.5 showed brain-localized reporter gene expression. These results suggest that mouse embryonic brain creates a closed and impermeable structure around E10. Therefore, by injecting a transgene before or after E10, transgene expression can be manipulated to be local or systemic. Our results also provide information that widens the applicability of UIGD beyond neuroscience studies.
GENETICALLY MODIFIED transgenic or knockout mice are used as tools to analyze mammalian gene functions in vivo. However, the generation of these mice is expensive, technically demanding, labor intensive, and time consuming. In addition, even if a tissue- or cell type–specific promoter is used to induce localized genetic changes instead of gross alterations in the entire body, 1 these models are not suitable to study cell-autonomous effects of genes.
To overcome these drawbacks, in utero injection was developed as an alternative method to produce animal models.2,3 This technique allows us to quickly and easily study the effects of overexpression or knockdown of a gene of interest in a wild-type genetic background.3,4 In particular, in utero injection of a retroviral vector aided by ultrasound backscatter microscopy (UBM) permits introduction of the gene at a far earlier stage of development than injection by the naked eye.2,5 Since ultrasound image-guided gene delivery (UIGD) was developed based on injection into the embryonic telencephalon, it has been applied mostly to uncover gene functions in the embryonic forebrain area. 6
In this study, we employed a bioluminescence imaging technique to determine whether the UIGD technique can be applied to gain-of-function experiments in the brain and other types of tissue and found that we can control the extent of the gene-transferred area up to the whole-body level by varying the embryonic day of injection. Our results provide information that widens the applicability of UIGD beyond neurodevelopmental studies.
Materials and Methods
Retroviral Vector Production and Transduction
The retroviral vector used in this study contains the murine stem cell virus (MSCV) long terminal repeat (LTR) to drive gene expression and an internal ribosome entry sequence (IRES) to allow bicistronic expression of green fluorescent protein (GFP) and firefly luciferase as reporter genes. 4 The method of retroviral vector production has been described previously. 7 Briefly, the retroviral construct was transfected into 293T cells with gag-pol (pCA-gag-pol) and env-expressing vector (VSV-G) using Lipofectamine-PLUS (Invitrogen, Carlsbad, CA). Supernatant was collected 48 hours after transfection, filtered through a 0.45 μm filter, and frozen at 80°C until used. Concentrated viral stocks were prepared by ultracentrifugation at 25,000 rpm for 90 minutes at 4°C in an SW32 rotor (Beckman-Coulter, Fullerton, CA). Pellets were resuspended in 50 μL of phosphate-buffered saline (PBS) at 4°C for about 12 hour, and virus aliquots were stored at −80°C.
In Vivo Injection into the Ventricle of the Embryonic Brain
All animal procedures were approved by the Institutional Review Board and conducted in the Laboratory Animal Research Center of Sungkyunkwan University. Timed pregnant CD-1 mice (Orient-Bio, Gyeonggi-do, Korea) were used for viral injections, and embryos were considered 0.5 days old when a vaginal plug was detected in the morning. Virus delivery into the telencephalic ventricle was performed at E9.5 or E10.5 using the UBM system as described previously. 6 Briefly, pregnant mice were anesthetized with Zoletil 50 (Virbac, Carros, France) and xylazine (Rompun, Bayer Korea, Korea). The uterus was exteriorized, and the fetuses were scanned using an ultrasound biomicroscopic imaging system (Vevo660, VisualSonics, Toronto, ON). Ultracentrifuge-concentrated viruses containing polybrene (final concentration 80 μg/mL) were injected into the telencephalic ventricle of E9.5 or E10.5 embryos.
Imaging of Luciferase Activity in Vivo
Mice were anesthetized with Zoletil 50 and xylazine. An aqueous solution of D-luciferin substrate (150 mg/kg) (Molecular Imaging Products, Ann Arbor, MI) was injected into the peritoneal cavity 10 minutes before imaging. Images were collected for 30 or 40 seconds using the IVIS 100 imaging system (Xenogen, Alameda, CA).
Immunohistochemistry
Standard immunostaining procedures were used for visualization of reporter gene expression in adult mice. Briefly, tissues from adult mice were fixed in 4% paraformaldehyde and cryosectioned. Sections were washed in PBS and then blocked for 1 hour with PBS containing 1% fetal bovine serum and 0.2% Triton X-100. Sections were incubated overnight at 4°C in anti-GFP primary antibody (Invitrogen) diluted in blocking solution, washed three times in PBS and 0.1% Tween 20, and incubated for 1 hour at room temperature with antirabbit IgG secondary antibody conjugated with Alexa Fluor 488 (green, Invitrogen) diluted in blocking solution. Images were acquired using a Zeiss (Oberkochen, Germany) LSM 510 confocal microscope with an Axiovert 100M system.
Results
In Vivo Bioluminescence Imaging of Animals Injected with Retroviral Vectors at Different Embryonic Ages
To compare the effect of injection day on transgene expression pattern, we performed UIGD into the embryonic brain at E9.5 or E10.5 as previously described (Figure 1).3,8 The retroviral vector used in this study expresses GFP and luciferase bicistronically under control of the MSCV LTR. 4 For visualization of gene expression, bioluminescence in vivo imaging was conducted in mice at 6 weeks and 1 year postinjection.

Ultrasound image-guided delivery into the embryonic forebrain ventricle. A, Schematic of the experimental setup (modified from Slevin JC et al 14 ). Representative real-time ultrasound backscatter microscopic images of E9.5 (B) and E10.5 (C) embryos. n = injection needle; v = ventricle. Scale bar = 1 mm.
The tissue distribution of transgene expression was clearly different between the injected mice. In all animals (10 of 10) injected at E9.5, luciferase activity was observed all over the body (Figure 2), whereas animals injected at E10.5 (6 of 6) consistently showed reporter gene expression restricted to the brain. E9.5 samples produced stronger bioluminescent signals than E10.5 ones throughout the animals, even in the brain. Indefinable luciferase activity was also observed in the tail and hind legs of animals injected at E10.5. Luciferase expression remained stable in both animal groups up to a year of age (data not shown). These results suggest that the 1-day difference between E9.5 and E10.5 results in a remarkable change in the diffusion of retroviral vector particles.

Comparison of transgene expression patterns between mice injected at E9.5 and E10.5. The same titer of retroviral vector expressing luciferase and green fluorescent protein as a bicistronic message was injected into the ventricle of the embryonic brain. Animals were analyzed by in vivo luciferase imaging at 6 weeks. Images are representative of 10 E9.5 and 6 E10.5 animals. The scales to the right of the images describe the color map for the luminescent signal.
Tissue Distribution of Luciferase Activity
To rule out the possibility that the widespread bioluminescence signals in animals injected at E9.5 may be emitted not from internal organs but from the skin and that fur could have reduced bioluminescent signals in animals injected at E10.5, the major organs (eg, liver, heart, lung, eyes, spleen, intestine, blood cells, spinal cord, and brain) were harvested from the 1-year-old animals and luciferase activities in each tissue were measured. In all organs from mice injected at E9.5, reporter activity was readily detected (Figure 3A), whereas exclusive signals in the brain were observed in animals injected at E10.5 (Figure 3B). Reporter gene expression in the deep layer cells of these tissues was further confirmed by immunohistochemistry using an anti-GFP antibody. Consistent with the results from luciferase imaging, GFP immunostaining was detected in all tissues from the E9.5 animals tested in this study (Figure 4). These results indicate that UIGD at E9.5 confers widespread transgene expression and UIGD at E10.5 very restricted expression in the brain.
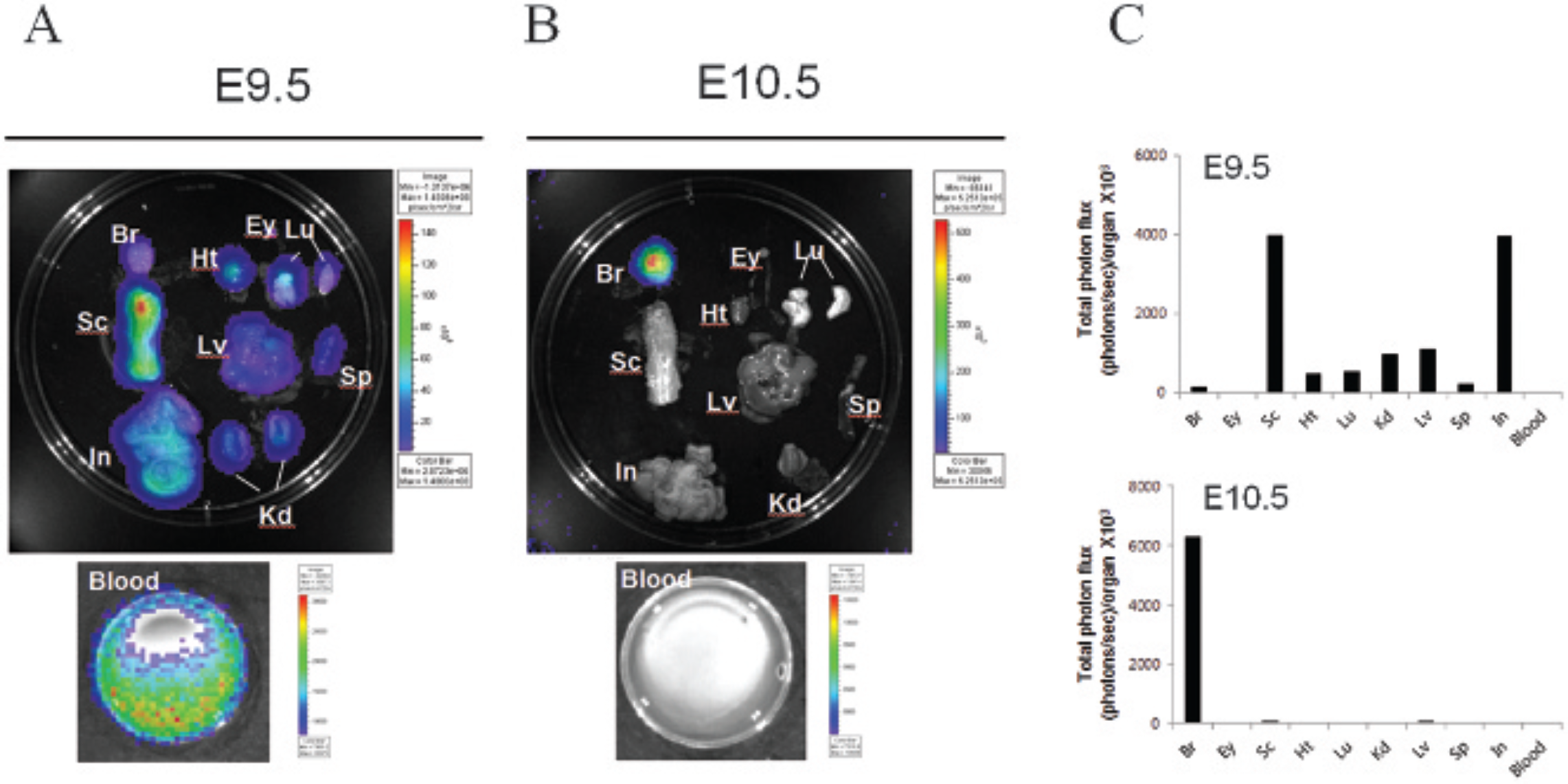
Transgene expression in the internal organs of mice injected with retrovirus at different embryonic ages. A retroviral vector expressing luciferase and green fluorescent protein was injected into the ventricle of E9.5 (A) or E10.5 (B) embryonic brains. Mice were injected intraperitoneally with D-luciferin at 1 year of age, and various organs were dissected and subjected to luciferase imaging. C, Bioluminescence from each organ of E9.5 and E10.5 animals is represented as the total photon flux/organ (photons/s). The scales to the right of the images describe the color map for the luminescent signal. Br = brain; Ey = eye; Ht = heart; In = intestine; Kd = kidney; Lu = lung; Lv = liver; Sc = spinal cord; Sp = spleen.

Immunohistochemical analysis of intraventricularly injected animals. Key internal organs, including the brain, spinal cord, liver, and kidney, were harvested from 1-year-old mice injected with retrovirus, and immunostaining using anti–green fluorescent protein antibody was performed on comparable sections from mice injected at E9.5 and E10.5. Scale bar = 100 μm.
Discussion
The embryonic telencephalon is an ideal target for the introduction of an exogenous gene of interest because it is composed of relatively large fluid-containing chambers forming the brain ventricular system. By using UBM, this tiny space (less than 0.5 mm in diameter at E9) can be targeted for injection in utero.5,9 For this reason, the UIGD technique has typically been used for gene function studies during embryonic brain development.
In the embryonic developmental process, two factors can be considered to be important for the extent of viral vector spread in the embryonic body after intraventricular injection: the open or closed state of the brain and the permeability of blood vessels. Early in development, the edges of the neural folds begin to move toward each other to create a hollow canal, the neural tube; the anterior portion of the neural tube becomes the brain; and the remainder becomes the spinal cord.
At around E9, the mouse brain and spinal cord are still open to amniotic fluid and then become closed structures.10,11 Also, blood vessels, which are highly permeable in the early telencephalon, become completely impermeable late in mouse development.12,13 These observations can explain why broader gene expression is obtained the earlier we inject. Our study suggests that the mouse embryonic brain appears to complete the formation of a closed and impermeable structure at E10.
For neuroscience research, injection at a later stage than E10 is highly recommended for brain-specific gene delivery because viral particles would be restricted to the brain ventricle region. However, if it is clear that transgene expression in other tissue does not produce problems in the interpretation of the results, injection at earlier than the E10 stage is also worth considering because more robust gene expression can be obtained in the brain.
Interestingly, E9.5 samples produced stronger bioluminescent signals than E10.5 samples. The E9.5 embryo is far smaller than the E10.5 embryo. Assuming that the same number of cells were exposed to viral vectors (same multiplicity of infection [MOI]), infection at an earlier stage of development would produce much stronger signals at the adult stage. It is also likely that the smaller ventricular space at E9.5 is advantageous in achieving a higher MOI than that at E10.5. In addition, downregulation of the cellular receptor for the retroviral envelope or production of a viral entry-inhibiting factor in cerebrospinal fluid might cause the weaker signals in E10.5 mice.
In summary, we show that the extent of viral vector diffusion can be controlled by choosing an appropriate embryonic day for injection and that UIGD can induce widespread and long-term transgene expression as well as localized brain-specific expression. Our results illuminate the usefulness of this means of quick and easy gene delivery compared to conventional transgenic techniques.
Footnotes
Financial disclosure of authors: This work was supported by Basic Science Research Programs through the National Research Foundation of Korea funded by the Ministry of Education, Science and Technology (#2012R1A1A2003185 and #2012R1A1A2008018).
Financial disclosure of reviewers: None reported.