
Abstract
Delivery of drugs to the appropriate target cells would improve efficacy and reduce potential side effects. The nontoxic B-subunit of the intestinal pathogen-produced Shiga toxin (STxB) binds specifically to the glycosphingolipid Gb3, overex-pressed in membranes of certain tumor cells, and enters these cells through the retrograde pathway. Therefore, STxB binding to Gb3 receptors may be useful for cell-specific vectorization or imaging purposes. Here we labeled STxB with a fluorophore to evaluate its potential as an in vivo cell-specific targeting reagent in two different models of human colorectal carcinoma. Fluorescent STxB was administered systemically to xenografted nude mice, and its biodistribution was studied by optical imaging. The use of fluorescent STxB allowed the combination of the macroscopic observations with analyses at the cellular level using confocal microscopy. After administration, the fluorescent STxB was slowly eliminated by renal excretion. However, it accumulated in the tumor area. Furthermore, STxB was demonstrated to enter the Gb3-expressing tumoral cells, as well as the epithelial cells of the neovascularization and the monocytes and macrophages surrounding the xenografts.
Shiga toxin is a bacterial toxin produced by Shigella dysenteriae, with AB5 subunit structure 6 ; the subunits are noncovalently linked. The A subunit is responsible for toxicity: its ribosomal ribonucleic acid N-glycanase activity inhibits protein biosynthesis. The nontoxic B-subunit of Shiga toxin (STxB) is responsible for targeting the toxin to cells and cell entry. It binds specifically to glycosphingolipid globotriaosyl ceramide (Gb3 or CD77), which serves as a cellular toxin receptor. Glycosphingolipids are abundant in the plasma membrane of nearly all eukaryotic cells and are essential for survival, proliferation, and differentiation. 7 Aberrant glycosylation owing to tumorigenesis is associated with changes in glycosphingolipids. 8 In particular, B-cell lymphomas, 9 ovarian hyperplasias, 10 cell suspensions obtained from human breast tumors, 9 testicular seminomas, 11 and colorectal carcinomas 4 all overexpress Gb3. After its binding to the Gb3 receptors, STxB uses the retrograde transport to deliver the A subunit to the cytosol: STxB enters cells through clathrin-independent or -dependent endocytosis, reaches the Golgi apparatus, and is then transported to the endoplasmic reticulum, from where the catalytic A subunit is retrotranslocated to the cytosol.3,12,13
Here we show that the B-subunit of the Shiga toxin accumulates in two models of human colorectal carcinoma, after systemic in vivo administration. A fluorescence-tagged protein was administered to nude mice bearing xenografts overexpressing the Gb3 receptor and its distribution was studied by optical imaging. Macroscopic observations were then correlated with confocal microscopy analysis to identify the targeted cells and to prove the entry of the protein in the Gb3-expressing cells.
Materials and Methods
Purification of STxB from Bacteria and Labeling
A variant of STxB carries a cysteine in its terminal chain, allowing conjugation of the protein with diverse molecules. 14 STxB was purified from bacteria as previously described, 15 dialyzed against phosphate-buffered saline (PBS), and coupled to fluorophores (Amersham Biosciences, Piscataway, NJ) according to the instructions from the supplier.
Cell Culture
HT29 human colon cancer cells were maintained at 37°C in Dulbecco's Modified Eagle's Medium (with glutamine, high glucose, without pyruvate; Invitrogen, Cergy Pontoise, France), supplemented with 4 mM of glutamine (Gibco), 5 mM sodium pyruvate (Gibco), 10% fetal bovine serum (Gibco), and 1% of a penicillin-streptomycin-amphotericin mix (Invitrogen, Cergy Pontoise, France). HT29 cells were infected with the pTRIP-deltaU3-EF1alpha EGFP-lentiviral vector containing the eGFP gene (enhanced green fluorescent protein) under the control of the EF1 (elongation factor 1) promoter. 16
Animals and Xenograft Models
Swiss nu/nu male or female mice were bred in the animal facilities of the Institut Curie, (Paris, France), or of the Commissariat à l'Energie Atomique, Service Hospitalier Frédéric Joliot (Orsay, France). The animals were maintained under specified pathogen-free conditions. Their care and housing were in accordance with the institutional guidelines of the French Ethical Committee (Ministère de l'Agriculture, Paris, France) and under the supervision of authorized investigators.
The TC302 tumor specimen was obtained with consent from a colon cancer patient during surgical resection of the tumor. The tumor sample was established as a xenograft by subcutaneous implantation of a tumor fragment into the scapular area of nude mice and sequentially transplanted. TC302 fragments of 30 to 60 mm3 were grafted subcutaneously into the inter-scapular fat pad of 5- to 7-week-old Swiss nude mice, under tribromoethanol anesthesia.
HT29 xenografts were obtained by subcutaneously injecting 2.5 × 106 green fluorescent protein (GFP)-expressing HT29 cells into nude mice, under isoflurane anesthesia.
Fluorescence Imaging
Biodistribution of fluorescent STxB in mice xenografted with GFP-expressing HT29 cells and TC302 tumor fragments was studied by fluorescence imaging. When the tumors reached approximately 250 mm3, the mice were injected with 50 μg of STxB-Cy5 (1.67 mg/kg). A PhotonImager optical camera (Biospace Lab) or an IVIS-Xenogen 100 system (Caliper Lifesciences) was used for optical imaging.
Acquisition parameters for the PhotonImager optical camera were as follows: excitation filter, 650 nm; emission filter, 715 nm; light intensity, 100%; aperture, 3.3; background correction filter, 520 nm. Fluorescent signal intensities were corrected for background and quantified using Photovision+ software (Biospace Lab).
Acquisition parameters for the IVIS-Xenogen 100 system (Caliper Lifesciences) were as follows: excitation filter, 615 to 665 nm; emission filter, 695 to 770 nm; background correction filter, 580 to 610 nm. The images were collected and processed using the Xenogen Living Image software (Caliper Lifesciences).
Immunohistochemical Analyses
Nude mice xenografted with GFP-expressing HT29 cells were injected intravenously with 100 μg of STxB-Cy5 (3.34 mg/kg). Tumors were collected 8 and 48 hours postinjection and incubated overnight at 4°C in an aqueous solution containing 4% of paraformaldehyde, 10 mM PBS, 0.1 mM CaCl2, and 1.0 mM MgCl2. Tumors were then incubated for 24 hours at 4°C in an aqueous solution containing 4% paraformaldehyde, 20% sucrose, 10 mM PBS, 0.1 mM CaCl2, and 1.0 mM MgCl2; frozen in isopentane; and stored at −80°C. Tumor sections (5.0 μm) were cut with a CM3050 cryostat (Leica). Tumor sections were stained with hematoxylin and eosin or incubated with monoclonal antibodies specific for CD34 (1 hour; 1:200 dilution; MEC 14.7: sc-18917; Santa Cruz Biotechnology) or CD11b (1 hour; 1:1,000 dilution; MCA711; Serptec). Secondary antibodies were goat antirabbit IgG coupled to Alexa594 (30 minutes; 1:1,000; A11007; Molecular Probes), and 4,6-diamidino-2-phenylindole (DAPI) was used to stain nuclei. To label the Gb3 sphingolipid, cryosections were incubated ex vivo for 30 minutes with STxB conjugated to the fluorophore Cy3, at a final concentration of 10 μg/mL in PBS containing 0.2% bovine serum albumin. Sections were observed under a confocal microscope (SP2, Leica; objective: x63) equipped for epifluorescence.
Results
We used optical imaging to study the accumulation of fluorescently labeled STxB in TC302 xenografts after administration by various routes (Figure 1). After oral administration of STxB-Cy5 to TC302-xenografted nude mice, no STxB fluorescence was detected in the tumors. In contrast, after intraperitoneal injection of STxB-Cy5, fluorescence was observed in the xenograft from 5 hours to 5 days postinjection; the STxB was slowly eliminated by renal excretion. After intravenous injection, the STxB-Cy5 similarly reached xenografted tumors, but the fluorescent signal was higher and persisted longer (up to 7 days). We therefore chose the intravenous route for subsequent experiments.

Influence of the route of injection on STxB-Cy5 biodistribution in TC302 xenografted nude mice. Nude mice bearing TC302 primary human tumors were given 50 μg (1.7 mg/kg) of STxB-Cy5 either per os (PO; A), intraperitoneally (IP; B), or intravenously (IV; C). Fluorescence was observed using an optical camera 5 hours and 2, 4, 5, and 7 days after STxB administration. The red arrows indicate the position of the xenografts. D, Fluorescence signal intensities (n = 4 for each routes of delivery) were corrected for background values and quantified using the Xenogen Living Image software (Caliper Lifesciences) according to the manufacturer's recommendations.
The targeting of Gb3-positive tumors by STxB was also studied in nude mice xenografted with HT29 cells, a human colorectal carcinoma cell line overexpressing the Gb3. The HT29 cells were stably transfected with the GFP reporter gene before implantation into the mice. Fluorescent STxB was injected intravenously into the xenografted mice. Fluorescence was rapidly accumulated in the HT29 xenografts, and its elimination from the body was slow (Figure 2). Injection of fluorescent STxB in non-tumor-bearing mice showed that the biodistribution of the protein was the same as in the tumor-bearing mice (data not shown). STxB-Cy5 was eliminated mainly by the renal system. Fluorescence was also observed in the spleen, the liver, and, to a lesser extent, the intestine (Figure 3). The tumor to muscle fluorescence ratio was 2.8 by 8 hours after the injection (Figure 2) and increased to 4.1 after 2 days, where it remained until day 4; thereafter, the ratio declined but was still 2.3 9 days postinjection.

Biodistribution of STxB-Cy5 in nude mice xenografted with GFP-expressing HT29 cells after intravenous administration. A, Bright field and fluorescence images of a representative xenografted nude mouse observed 8 hours and 4 days after intravenous injection of 1.7 mg/kg STxB-Cy5. The white arrows indicate the position of the xenografts. B, Tumor to muscle ratio. Fluorescence signal intensities in the xenograft and in the muscles of the mice (n = 6) were corrected for background values and quantified using the Photovision+ software (Biospace Lab).

Histologic analysis of organs displaying fluorescence after STxB-Cy5 injection (red); 3.34 mg/kg of STxB-Cy5 were injected in non-tumor-bearing nude mice. Eight hours postinjection, kidneys (A), liver (B), intestine (C), and spleen (D) were analyzed by confocal microscopy (Leica SP2; objective: x63). Tumor-bearing nude mice also received 3.34 mg/kg of STxB-Cy5, and tumors 8 hours postinjection (E) were analyzed by confocal microscopy (Leica SP2; objective: x63). F, G, H, and I: Kidneys, liver, intestine and spleen of a control mouse, noninjected with STxB-Cy5. Nuclear staining with DAPI in blue.
Fluorescent STxB and GFP expression by cancerous cells allowed analysis of tumoral cell targeting at the cellular level by using confocal microscopy. Mice were sacrificed 8 and 48 hours after intravenous injection of STxB, and the tumors were collected. To compare the presence of fluorescent STxB and that of Gb3, tumor sections were stained ex vivo with STxB-Cy3 and observed by confocal microscopy (Figure 4). Despite a heterogeneous expression of Gb3 on HT29 tumor cells, the staining revealed good colocalization of STxB-Cy5 and Gb3. STxB accumulated inside GFP-positive HT29 cells after both 8 and 48 hours (Figure 5).
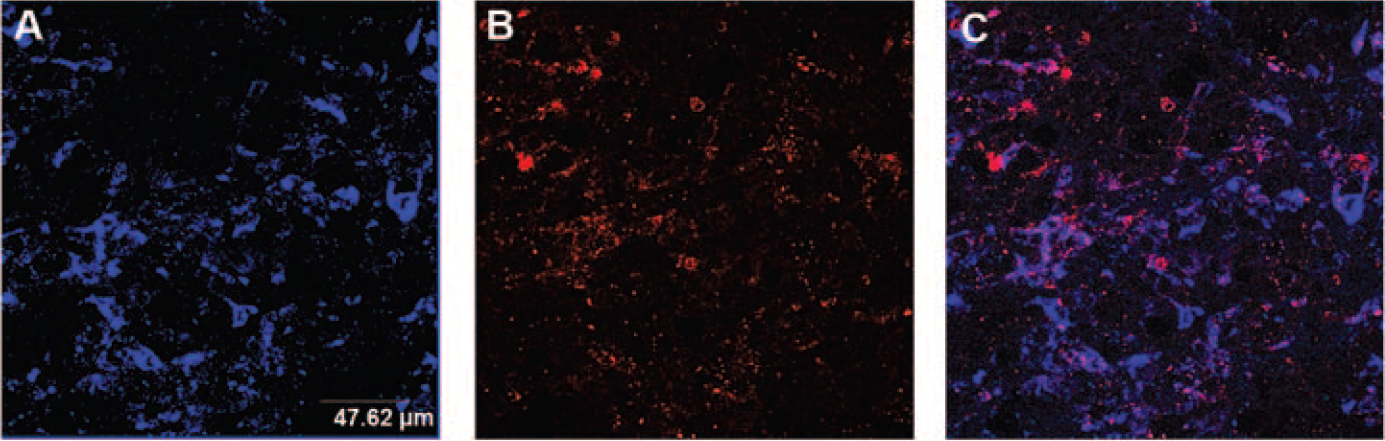
STxB accumulation in the xenograft coincides with Gb3 expression. Forty-eight hours after STxB-Cy5 intravenous injection in xenografted nude mice, tumors were collected and frozen and sections were made. Tumor sections were labelled for Gb3 by ex vivo incubation with Cy3-labeled STxB and were observed by confocal microscopy (Leica SP2; objective: x63). A, Injected STxB-Cy5; B, Gb3-positive cells; C, overlay.

Targeting of HT29 cells by STxB. Forty-eight hours after STxB-Cy5 intravenous injection in xenografted nude mice, tumors were collected and frozen and sections were made. Tumor sections were observed by confocal microscopy (Leica SP2; objective: x63). A, Injected STxB-Cy5; B, GFP-expressing HT29 cells; C, overlay.
Moreover, accumulation of STxB was also observed in GFP-negative cells. To identify these targeted cells accurately, tumor sections were labeled with anti-CD11b antibody to detect infiltrating macrophages (Figure 6) and with anti-CD34 antibody to reveal epithelial cells of new blood vessels (Figure 7). The anti-CD11b antibody (also known as Mac-1, CR3, or Integrin αM, Itgam) binds to a transmembrane glycoprotein found on granulocytes, monocytes, natural killer cells, and subsets of T and B cells. This staining revealed the presence of an area of inflammation surrounding the xenografts and substantial accumulation of the STxB-Cy5 in CD11b-positive cells (see Figure 6); tumors collected 8 and 48 hours after STxB-Cy5 injection showed similar patterns. The anti-CD34 antibody is specific for a heavily glycosylated transmembrane glycoprotein expressed on hematopoietic progenitor and vascular endothelial cells. This staining showed that the epithelial cells of the new blood vessels also accumulated STxB (see Figure 7, A-C). Most of the HT29 cells labeled with STxB-Cy5 were located near blood vessels (see Figure 7, A-C). However, dual labeling for CD34 and Gb3 showed that the strength of Gb3 expression by HT29 cells was not correlated with the proximity to blood vessels (see Figure 7, D-F), highlighting the role of the diffusion of STxB for explaining the heterogeneity of the protein accumulation.

Targeting of inflammatory regions by STxB. Forty-eight hours after STxB-Cy5 intravenous injection in xenografted nude mice, tumors were collected and frozen and sections were made. HT29 xenograft sections were labeled for macrophages with anti-CD11b antibodies and observed by confocal microscopy (Leica SP2; objective: x63). A, Injected STxB-Cy5; B, CD11b-positive cells; C, comparison of CD11b-expressing cells and accumulation of STxB-Cy5; D, GFP-expressing HT29 cells.

Targeting of blood neovessels by STxB. Forty-eight hours after STxB-Cy5 intravenous injection in xenografted nude mice, tumors were collected and frozen and sections were made. HT29 xenograft sections were labeled for neovessels with anti-CD34 antibodies and observed by confocal microscopy (Leica SP2; objective: x63). A, Injected STxB-Cy5; B, CD34-positive cells; C, overlay of A and B. Tumors of xenografted mice noninjected with STxB-Cy5 were also collected. Tumor sections were labeled for neovessels with anti-CD34 antibodies and for Gb3-positive regions by ex vivo incubation with Cy3-labeled STxB. D, Gb3-positive cells; E, CD34-positive cells; F, overlay of D and E.
Discussion
STxB has been used to study retrograde transport, and the mechanisms governing this pathway are now better known.2,17 STxB specifically enters Gb3-positive cells, and this sphingolipid is abundant on a large number of cancerous cells. Therefore, STxB is a potentially useful agent for drug vectoring, although there have been only a few detailed analyses of in vivo targeting of Gb3-overexpressing cells by STxB and its biodistribution after systemic administration. To assess the value of STxB as an in vivo vectorization agent, we used two models of colorectal carcinoma. Colorectal carcinomas—one of the most frequent tumors in Europe and North America—often overexpress Gb3. 5 The biodistribution of the STxB has been analyzed by positron emission tomography (PET) in a transgenic mouse model of spontaneous digestive tumorigenesis 4 : following per os or intravenous administration, STxB accumulated in intestinal tumor regions overexpressing Gb3. This PET study also showed that the radioactivity was eliminated through the urinary tract, as shown by high levels of radioactivity in the bladder and kidneys. Uptake was also observed in the spleen, lungs, and, to a lesser extent, the liver. In our xenograft models, STxB accumulation was also observed in Gb3-expressing tumor regions, confirming that cancerous cells are targeted in vivo after systemic injection. We also report that the STxB persisted for several days in the circulation and particularly in the xenografts. We observed that the main route of elimination of the protein was the urinary pathway, confirming the previous PET study. We also observed an accumulation of fluorescence in the liver and the intestine. In the PET study, no radioactivity could be detected in the abdominal region of wild-type mice, and only a slight labeling was observed in the liver. These differences may be due to the different methods of labeling or to the different species of mice. The presence of fluorescence in the kidneys, spleen, and liver confirms that it will be of primary importance to study the possible adverse effects on these organs during the development of new therapeutic strategies using STxB.
We also used fluorescently labeled STxB to compare macroscopic optical imaging patterns and the cellular distribution by confocal microscopy. Bartlett and colleagues showed that two compounds displaying similar tumor accumulation, as assessed by macroscopic analysis (by PET in their study), may have different intracellular and extracellular distributions in a tumor. 18 The use of a fluorescently labeled probe allows analysis of the tumor area at the cellular level to determine which types of cells are really targeted by the probe, if this targeting is receptor specific, and if the probe enters in the targeted cells. Confocal microscopy with tumor sections confirmed that STxB accumulated in HT29 cells expressing its receptor Gb3, demonstrating the efficacy of STxB as a vector for targeted delivery to Gb3-expressing tumor cells. STxB staining of the xenograft was heterogeneous, and there are two explanations: (1) accumulation of STxB in HT29 cells was enhanced by the proximity of blood vessels, and (2) Gb3 expression by HT29 cells was not homogeneous throughout the xenograft. The epithelial cells of the new blood vessels also accumulated STxB. These in vivo findings are consistent with previous ex vivo observations of fluorescent Shiga toxin binding to the neovasculature of frozen tumor sections.10,19,20 This specificity for neovasculature offers interesting possibilities for the STxB vector, including the delivery of antiangiogenic drugs, even to tumors that do not express Gb3. CD11b-positive cells (infiltrating macrophages, natural killer cells, and B-cell subpopulations) also accumulated STxB, offering opportunities for in vivo delivery of immunotherapeutic agents. Indeed, Vingert and colleagues demonstrated that Shiga toxin B subunit can target antigen to dendritic cells in vivo and can elicit antitumor immunity. 21
In conclusion, using two models of human colorectal carcinoma, we showed that STxB can target cells associated with Gb3-expressing tumors after a systemic injection. Furthermore, the protein entered Gb3-expressing cells, including the cancerous cells, as well as monocytes and macrophages and epithelial cells of the new blood vessels. These results illustrate the great potential of this protein as a vector for targeted antitumoral therapy.