
Abstract
Low capacity runner (LCR) rats have been developed by divergent artificial selection for treadmill endurance capacity to explore an aerobic biology-disease connection. The beneficial effects of resveratrol supplementation have been demonstrated in endurance running. In this study it was examined whether 12 weeks of treadmill exercise training and/or resveratrol can retrieve the low running performance of the LCR and impact mitochondrial biogenesis and quality control. Resveratrol regressed running performance in trained LCR (p<0.05). Surprisingly, exercise and resveratrol treatments significantly decreased pAMPK/AMPK, SIRT1, SIRT4, forkhead transcription factor 1 (FOXO1) and mitochondrial transcription factor A (TFAM) levels in these animals (p<0.05). Mitochondrial fusion protein, HSP78 and polynucleotide phosphorylase were significantly induced in LCR-trained, LCR-resveratrol treated, LCR-trained and resveratol treated groups compared to LCR-controls. The data indicate that the AMPK-SIRT1-NAMPT-FOXO1 axis could be important to the limited aerobic endurance capacity of low running capacity rats. Resveratrol supplementation was not beneficial in terms of aerobic endurance performance, mitochondrial biogenesis, or quality control.
INTRODUCTION
Large-scale epidemiologic studies demonstrate a strong statistical association between low capacity for aerobic exercise and increased risk for development of complex diseases (DeMarco et al. 2012). Although an underlying mechanistic relationship has been hypothesized (aerobic hypothesis), the complexity of both aerobic metabolism and the putative disease conditions in humans makes the unraveling of cause and effect a challenge (Kivela et al. 2010). In 1996 Koch and Britton (Koch and Britton 2001) initiated a prospective test of the linkage between aerobic capacity and disease risk by applying large-scale artificial selective breeding in rats with widely varying genetic backgrounds to produce low and high strains that differ for intrinsic (i.e., untrained) aerobic endurance treadmill running capacity (Koch and Britton 2001). The hypothesis was that rats selectively bred as Low Capacity Runners (LCR) would display disease risks and rats bred as High Capacity Runners (HCR) would have positive health effects. Several studies report that the LCR present with negative health features including metabolic syndrome, reduced heart function, hepatic steatosis, disordered sleep, and diminished oxidative capacity in skeletal muscle (Wisloff et al. 2005; Thyfault et al. 2009; Kivela et al. 2010). In contrast, HCR demonstrate greater maximal oxygen consumption, insulin sensitivity and improved metabolic health. In accordance with the aerobic hypothesis, aged rats with low intrinsic aerobic capacity have diminished longevity and display a reduced ability for mitochondrial regeneration, decreased metabolic control in the heart, and reduced antioxidant status (Koch and Britton 2001). Therefore, this model is excellent to study how life-style interventions, such as exercise training or nutritional manipulation, could overcome or exacerbate the effects of genetics on physiological and biochemical processes that lead to health promotion (Kamei et al. 2004; Wisloff et al. 2005; Bowman et al. 2010).
Exercise training and resveratrol have been suggested for therapeutic potential in treatment of metabolic disorders. Both interventions increase oxidative metabolism in skeletal muscle by induction of a highly integrated molecular network which results in increased insulin sensitivity (Nunn et al. 2010), activity of AMP-activated protein kinase, (AMPK) peroxisome proliferator-activated receptor-γ coactivator 1α (PGC-1α), and mitochondrial content (Baur et al. 2006; Li et al. 2011). Recent findings reveal that knockdown of a mitochondrial located sirtuin, SIRT4, results in increased fatty acid oxidation, enhanced mitochondrial function, and higher AMPK levels in skeletal muscle (Nasrin et al. 2010), suggesting a primary role of this sirtuin in aerobic metabolism. Moreover, another member of the sirtuin family, the NAD+ dependent SIRT1, is also an important regulator of oxidative mitochondrial metabolism (Rodgers et al. 2008) by deacetylation of the peroxisome proliferator-activated receptor gamma co-activator 1 alpha (PGC-1α) (Nemoto et al. 2005). PGC-1α activates gene transcription factors important for mitochondrial biogenesis including nuclear respiratory factor 1 (NRF1) and mitochondrial transcription factor A (TFAM), which encode nuclear and mitochondrial DNA imprinted genes required for the production of mitochondrial proteins. Mitochondrial fusion and fission are important mechanisms for maintenance of the mitochondrial network and for quality control (Westermann 2010), and thus impact mitochondrial function (Otera and Mihara 2011). The quality control of mitochondrial proteins is supervised by Lon protease and HSP78, which prevent the accumulation of oxidized and dysfunctional proteins in mitochondria (Bota and Davies 2002; Rottgers et al. 2002; Ngo and Davies 2009). Recent studies have revealed that polynucleotide phosphorylase (PNPase) is involved in the stability of mitochondria (Chen et al. 2006) and is crucial for the import of nuclear coded RNAs into the mitochondrial matrix (Wang et al. 2010).
It is well known that similar exercise programs can result in different training responses for different subjects (Radak et al. 2013). However, the possible different responses to nutritional and pharmacological interventions, including resveratrol treatment, remain to be well characterized. Resveratrol has been shown to reduce plasma triglyceride concentrations, oxidative stress, and inflammation in humans (Zern et al. 2005), as well as to improve insulin sensitivity in rats (Zheng et al. 2012).
Therefore, in the present study, the LCR contrasting rat model system was utilized to test the influence of exercise training, and resveratrol singularly, and the combined effects of training and resveratrol, upon indices of health, including running performance, VO2max, and forearm gripping strength. We were particularly interested to further understand retrieval of the negative LCR clinical phenotype. Positive effects of exercise training and resveratrol treatment on the LCR group would mean that life-style interventions could compensate, in some degree, for the health treating effects of a poor genetic setup. Both physical exercise and resveratrol can modulate the generation and activity of reactive oxygen species (ROS), which are implicated in a variety of diseases, and signaling, and mitochondrial biogenesis (Radak et al. 2013). Therefore, as an additional aim, the relative density of mtDNA and activities of a number of factors that regulate mitochondrial biogenesis, quantity and quality control and differentiation from gastrocnemius muscle, were investigated for possible explanations.
METHODS
Animals
Artificial selective breeding, starting with a founder population of 186 genetically heterogeneous rats (N:NIH stock), was used to develop rat strains differing in inherent aerobic capacity. The procedure has been described in detail previously (Koch and Britton 2001). Briefly, endurance running capacity was assessed on a treadmill and the total distance run during a speed-ramped exercise test was used as a measure of maximal aerobic capacity. Rats with the lowest capacity were bred to produce the LCR strain. A subgroup of 24 male rats from generation 22 was phenotyped for intrinsic treadmill running capacity when 11 weeks old, at the University of Michigan (Ann Arbor, USA), and then shipped via air freight to Semmelweis University (Budapest, Hungary) for further study. Investigations were carried out according to the requirements of The Guiding Principles for Care and Use of Animals, EU, and approved by the ethics committee of Semmelweis University.
Exercise protocol and resveratrol treatment
Twenty four LCR male rats, aged 13 months, were equally assigned to control (LCR-C), trained (LCR-Tr), resveratrol treated control (LCR-Rsv), and trained resveratrol treated (LCR-TrRsv), groups. Control rats had access to a treadmill with an electrical stimulator, three times a week, for ten min. Trained rats were introduced to treadmill running for three days, then for the next two weeks the running speed was set to 10 m/min, on a 5% incline for 30 min. The treadmill was equipped with a high pressure air pipe and electric grid to stimulate running.
In the following week, maximal oxygen uptake (VO2max) was measured on a motor driven treadmill (Columbus Inst. Columbus, Ohio) with a gradually increasing intensity. VO2max was measured for each animal, using three criteria: (i) no change in VO2 when speed was increased, (ii) rats no longer kept their position on the treadmill, and (iii) respiratory quotient (RQ = VCO2/VO2) > 1. Based on the level of VO2max, a treadmill speed corresponding to 60% VO2max was determined and used for daily training for one hour, five times per week. VO2max was measured every second week and running speed was adjusted accordingly. The total training period lasted 12 weeks. In addition, the forelimb strength of the animals was assessed weekly by using a gripping test as described by Marton
Resveratrol supplementation (100 mg/kg, oral dose) (Smith et al. 2009) was started two weeks before habitual treadmill running was introduced to the animals, and four weeks before the actual training started, thereby lasting 16 weeks. The animals were sacrificed two days after the last exercise session to avoid the acute metabolic effects of the final run. The gastrocnemius muscle was dissected and homogenized in buffer (HB) containing 137 mM NaCl, 20mM Tris-HCl (pH 8.0), 2% NP 40, 10% glycerol and protease inhibitors.
ROS, and protein carbonyl content and antioxidant enzyme activities
Intracellular oxidant and redox-active iron levels (Kalyanaraman et al. 2012) were estimated using modifications of the dichlorodihydrofluorescein diacetate (H2DCFDA) staining method (Radak et al. 2004). The oxidative conversion of stable, nonfluorometric DCF-DA to highly fluorescent 2′7′-dichlorofluoorescein (DCF) was measured in the presence of esterases, as previously reported (Radak et al. 2004). This assay approximates levels of reactive species, such as superoxide radical, hydroxyl radical, and hydrogen peroxide. The method has been widely used in the literature but does have the problem of not being particularly specific, and results can be strongly affected by release of labile iron or copper (Kalyanaraman et al. 2012). Briefly, the H2DCFDA (Invitrogen-Molecular Probes #D399) was dissolved to a concentration of 12.5 mM in ethanol and kept at −80 °C in the dark. The solution was freshly diluted with potassium phosphate buffer to 125 μM before use. For fluorescence reactions, 96-well, black microplates were loaded with potassium phosphate buffer (pH 7.4) to a final concentration of 152 μM/well. Then eight μl diluted tissue homogenates and 40 μl 125 μM dye were added to achieve a final dye concentration of 25 μM. The change in fluorescence intensity was monitored every five minutes for 30 minutes with excitation and emission wavelengths set at 485 nm and 538 nm (Fluoroskan Ascent FL) respectively. Data obtained after 15 min were used. The fluorescence intensity unit was normalized with the protein content and expressed in relative unit production per minute. The protein carbonyl measurement was assessed as described earlier (Koltai et al. 2012).
Western blots
Ten to 50 micrograms of protein were electrophoresed on 8–12% v/v polyacrylamide SDS-PAGE gels. Proteins were electrotransfered onto PVDF membranes. The membranes were subsequently blocked and incubated at room temperature with antibodies (1:500 #sc-13067 Santa Cruz PGC-1 (H-300), 1:1400 #2532 Cell Signaling AMPKα, 1:500 #2535 Cell Signaling p-AMPKα (Thr172) (40H9), 1:1000 #sc-33771 Santa Cruz NRF-1 (H-300), 1:500 #sc-30963 Santa Cruz mtTFA (E-16) /TEAM/, 1:500 #sc-98900 Santa Cruz Fis1 (Fl-152), 1:10000 #sc-50330 Santa Cruz Mfn1 (H-65), 1:1000 #sc-99006 Santa Cruz PNPase (H-124), 1:200 #U7757 Sigma-Aldrich UCP3, 1:5000 #ab87253 Abcam CLPB /HSP78/, 1: 15000, #T6199 Sigma alpha-tubulin). The antibody for Lon protease was generated in our laboratory, as described previously (Merrill et al. 1997). After incubation with primary antibodies, membranes were washed in TBS-Tween-20 and incubated with HRP-conjugated secondary antibodies. After incubation with the secondary antibody, membranes were repeatedly washed. Membranes were incubated with an ECL Plus reagent (RPN 2132, Amersham) and protein bands were visualized on X-ray films. The bands were quantified by ImageJ software, and normalized to tubulin, which served as an internal control.
Assessment of SIRT1 activity
To measure SIRT1 deacetylase activity, a Cyclex SIRT1/Sir2 Deacetylsase Fluorometric Assay Kit (Cyclex, CY-1151) was used according to the established protocol including the separation of nuclear extract (Koltai et al. 2012). The purity of nuclear extract was checked by organelle specific antibodies as reported earlier (Radak et al. 2009).
To prepare nuclear fractions, the homogenate was centrifuged at 1000 ×
Measurement of mtDNA by PCR
The mtDNA content was quantified as the mtDNA to nuclear DNA (nDNA) ratio (mtDNA/nDNA). Total DNA was extracted (Fast DNA kit #6540-400 BIO 101 Systems Qbiogene) and quantified spectrophotometrically. The mtDNA content was measured by PCR (Rotor-Gene 6000, Corbett Research, Australia) using the following conditions: 94 °C for two min (initialdenaturation), 94 °C for 30 s, 60 °C for 30 s, 72 °C for 45 s (25 cycles), 72 °C for ten min (finalextension), and corrected by the simultaneous measurement of a single copy nuclear BDNF gene. Primers used for the analysis of mtDNA were R-CYTB-F (5′-CCC CAG AGG ATT AAA CTC CAA CGC A-3′), and R-CYTB-R (5′-GGG TGG GGT CAG GGG GT-3′). Primers used for the analysis of nDNA were R-BDNF-genom-exon-IV-F (5′- TTG GGA TGG GAA AGA TGG G −3′), and R-BDNF-genom-exon-IV-R (5′-CAG AGT AGG AGG GAA CAA GTG TGA C −3′). The mtDNA content was normalised to nDNA. Data are expressed as the mean of three measurements.
Statistical analyses
Because of the limited sample size and for the purpose of finding the appropriate statistical procedure, normality was tested on all dependent variables (Shapiro Wilk's W-test). It was found that few of the dependent variables demonstrated a normal distribution, namely: NRF1, Fis1, NAMPT, LonP, Mfn1, and VO2max. Therefore, non-parametric Kruskal-Wallis ANOVA analysis was used to test for differences among the dependent variables. The Mann-Whitney U-test was applied for post-hoc analyses. Significance level was set at p < 0.05.
RESULTS
Exercise performance
The body mass of the LCR-Tr group was significantly lower (p<0.001) (470±47g) than LCR-C (609±38g).
Exercise training significantly increased the level of running distance of LCR-Tr (p<0.01) compared to control and resveratrol treated rats (p<0.05) (Fig. 1A) and similar results were present for the levels of VO2max (Fig. 1B). The gripping strength of the upper limbs did not show significant differences between groups (Fig. 1C).

Running distance (A), VO2max (B), and gripping time (C) of LCR rats. Running distance (m) (A), maximal oxygen uptake (VO2max; ml/kg/min) (B) and gripping time (s) (C) for low capacity runner (LCR) rats were measured every second week across a 12 week exercise training period and during the two weeks of treadmill habituation. Control LCR (LCR-C), trained LCR (LCR-Tr), resveratrol treated control LCR (LCR-Rsv), trained resveratrol treated LCR (LCR-TrRsv) groups. Values are means ± SD for six animals per group *Significantly different from control group, + Significantly different from trained group, # Significantly different from resveratrol treated group, p < 0.05.
Effects of exercise and resveratrol on mitochondrial metabolic factors
AMPK activity is a known modulator of PGC-1α. Surprisingly, exercise and resveratrol treatments significantly decreased pAMPK/AMPK levels in LCR rats compared to control animals (p<0.05) (Fig. 2A). Interestingly, the changes in SIRT1 levels were very similar in pattern to AMPK activity although to a lesser degree (Fig. 2B). Nicotinamide phosphoribosyltransferase (NAMPT) was measured because it is involved in the biosynthesis of NAD+ and SIRT1 is dependent on the availability of NAD+. The NAMPT levels were decreased in both exercising groups (p<0.05) (Fig. 2C). Despite the decreasing activity of SIRT1, PGC-1α levels increased with exercise training (Fig. 2D). FOXO1 is an important transcription factor for regulation of gluconeogenesis and adipogenesis and in this model resveratrol alone and with training decreased FOXO1 levels (Fig 2E). Mitochondrial sirtuin, SIRT4, is related to fat metabolism and the data revealed that exercise training and resveratrol treatment decreased SIRT4 content (Fig 2F). mtDNA levels increased significantly with exercise and resveratrol (Fig 2G).

The effect of exercise and resveratrol on metabolic factors in skeletal muscle of LCR rats. The activity of AMPK, assessed by the pAMPK/AMPK ratio (A), which decreased with training and resveratrol treatment. The activity of SIRT1 was not changed by exercise or resveratrol (B). The protein levels of NAMPT (C), PGC-1a (D), FOXO1 (E) and SIRT4 (F) were determined by Western blot and densitometry in gastrocnemius muscle. G panel shows the mtDNA content. Control LCR (LCR-C), trained LCR (LCR-Tr), resveratrol treated control LCR (LCR-Rsv), trained resveratrol treated LCR (LCR-TrRsv) groups. Values are means ± SD for six animals per group *Significantly different from control group, + Significantly different from trained group, # Significantly different from resveratrol treated group, p < 0.05.
The effects of exercise and resveratrol on mitochondrial quantity and quality control
TFAM protein levels decreased in LCR-Tr and LCR-TrRsv rats compared to controls (p<0.05) (Fig. 3A). The concentration of NRF-1 was not affected by exercise training or resveratrol treatments (Fig. 3B). Mfn1 is a mediator of mitochondrial fusion and was significantly induced in LCR-Tr, LCR-Rsv, and LCR-TrRsv groups compared to LCR-C (Fig. 3C). The mitochondrial fission controlling protein, Fis1, was not affected by exercise training in any of the LCR groups (Fig 3D). Lon protease was measured to assess the level of protein degradation in mitochondria, which could be important for quality control. However, significant differences were not noted in all groups (Fig. 3E). HSP78 and PNPase were induced by both training and resveratrol (Fig. 3F, Fig. 3G).

Markers of quantity and quality control of mitochondria. Exercise and resveratrol altered the levels of TFAM (A) while the NRF1 concentration was unchanged (B). We evaluated mitochondrial fusion, Mfn1(C), and fission Fis1 (D) and quality control was assessed by Lon protease (E), HSP78 (F) and PNPase (G) levels. Control LCR (LCR-C), trained LCR (LCR-Tr), resveratrol treated control LCR (LCR-Rsv), trained resveratrol treated LCR (LCR-TrRsv) groups. Values are means ± SD for six animals per group *Significantly different from control group, + Significantly different from trained group, # Significantly different from resveratrol treated group, p < 0.05.
Oxidative stress markers
The levels of ROS, as measured by the fluorescent activity of H2DCFDA, increased with exercise training (Fig. 4A). The levels of carbonyl groups were assessed as potential markers of oxidative protein damage and the data clearly indicate decreased levels in LCR-TrRsv rats compared to controls (p<0.05) (Fig. 4B).
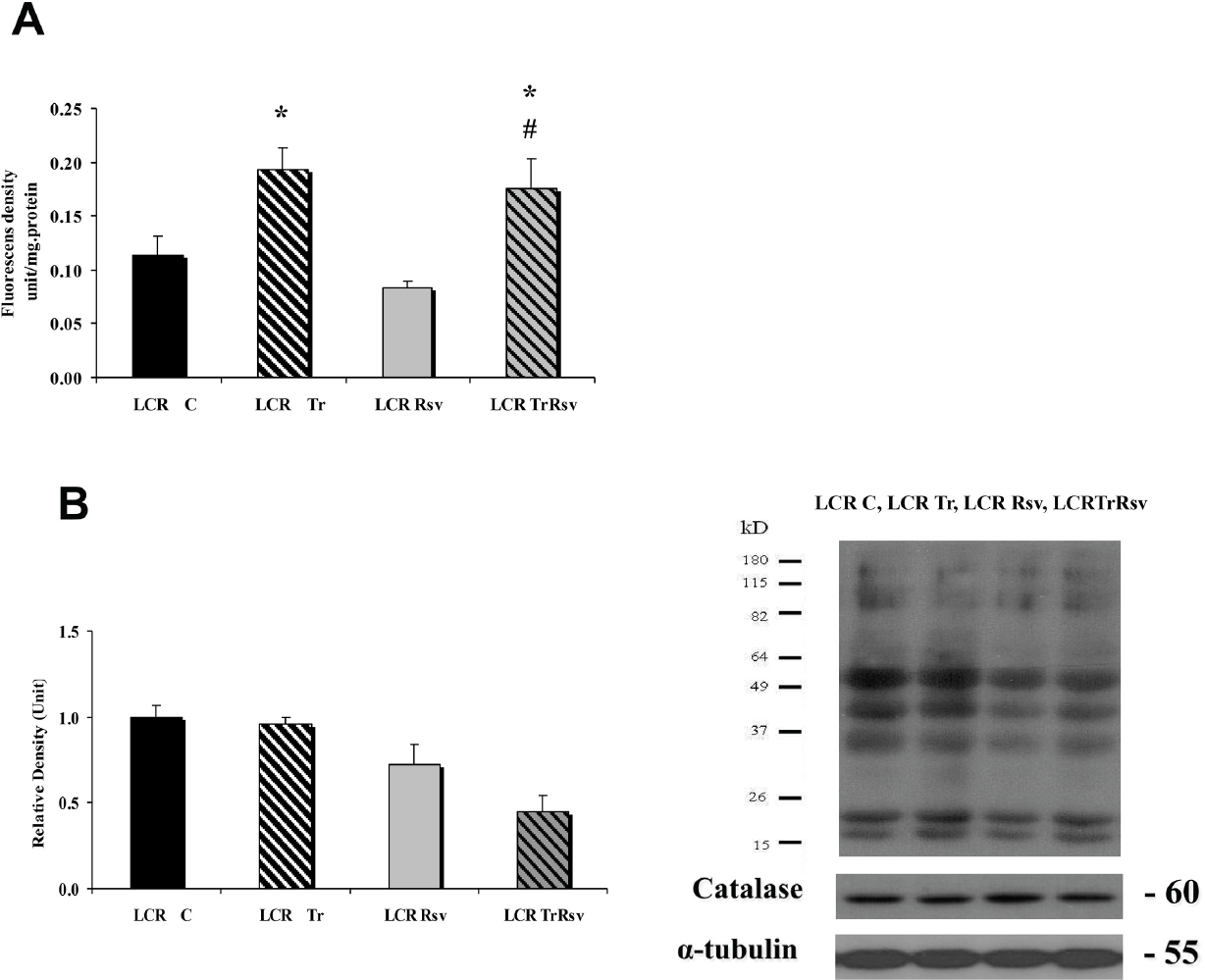
Oxidative stress markers. Rat gastrocnemius muscle was stained with dichlorodihydrofluorescein diacetate (H2DCFDA) to measure relative steady-state oxidant levels and redox-active iron release levels (both increase DCF fluorescence) as an estimate of levels of reactive oxygen species (ROS) (A). The oxidative damage of proteins was evaluated by protein carbonyl groups (B). Control LCR (LCR-C), trained LCR (LCR-Tr), resveratrol treated control LCR (LCR-Rsv), trained resveratrol treated LCR (LCR-TrRsv) groups. Values are means ± SD for six animals per group *Significantly different from control group, + Significantly different from trained group, # Significantly different from resveratrol treated group, p < 0.05.
DISCUSSION
Low running capacity rats have shown a wide range of physiological disorders (Wisloff et al. 2005; Thyfault et al. 2009; Kivela et al. 2010), which suggests that the increases in aerobic performance could be important to overcome some of these disorders. We have demonstrated in this study, that exercise training alone can increase VO2max of LCR rats. Since higher VO2max is associated with longer life-spans of LCR and HCR rats (Koch and Britton 2001) it can be suggested that the enhanced endurance ability would result in an increased life-span for LCR rats and reduce the metabolic disorders associated with the low endurance capacity of LCR rats.
In the present model resveratrol supplementation was not beneficial, although according to the general view, resveratrol supplementation promotes health. However, it must be noted that in some models resveratrol supplementation has been reported to act differently on obese prone and obese resistant rats (Louis et al. 2012). Therefore, the response to resveratrol treatment could be highly selective and could be based on metabolic status.
One of the striking observations of this study was that exercise training decreased AMPK activity in this rat model. AMPK acts as an energy sensor to cope with metabolic challenges and can interfere with fuel preference and availability (Kurth-Kraczek et al. 1999). Therefore, the exercise related decreased activity of AMPK could, at least in part, explain the limited running capacity of LCR rats.
It has been shown that exercise training increases NAMPT levels in skeletal muscle of wild rats (Koltai et al. 2010) and humans (Costford et al. 2010). NAMPT is important to NAD production which serves as a cofactor for sirtuins. Indeed, the activity of SIRT1 tends to decrease in a similar pattern to NAMPT and AMPK. AMPK has been shown to be linked to the activity of SIRT1, since AMPK can activate SIRT1 by increasing the levels of NAD+ (Canto et al. 2009) which, as a metabolic sensor, can control the expression of a number of genes involved in the metabolism of skeletal muscle (Canto et al. 2009). The AMPK associated decreased activity of SIRT1 could lead to increased acetylation of FOXO1, which is a target for SIRT1 deacetylase (Canto et al. 2009). Increased acetylation of FOXO1 attenuates the activity of this transcription factor (Nakae et al. 2012). The present data show decreased activity of AMPK and SIRT1 and decreased content of NAMPT and FOXO1 with exercise training. We strongly believe that this “unusual” response to exercise causes the limitation of aerobic capacity which limits the endurance capacity of low running capacity rats. This suggestion is based on the fact that wild rats respond to exercise training with increased AMPK-SIRT1-NAMPT (Costford et al. 2010; Koltai et al. 2010) and FOXO1 levels (Canto et al. 2009). In addition, other data from our laboratories on HCR rats suggests up-regulation of the AMPK-SIRT1-NAMPT-FOXO1 system (Hart et al. 2013).
The exact function of mitochondrial located SIRT4 is still in debate, but a recent study revealed that knockdown of SIRT4 by shRNA significantly increased fat metabolism in myotubes (Nasrin et al. 2010). This is an important finding and could indicate that lower levels of SIRT4 result in increased free fatty acid utilization, a key element for increased endurance capacity (Holloszy and Coyle 1984; Spina et al. 1996). Our present observations in trained and resveratrol supplemented rats suggests that the high body mass observed in control LCR rats could be due to impaired utilization of fat, which is enhanced by exercise training and resveratrol supplementation. Moreover, it is also known that SIRT4 is involved in the development of insulin resistance (Chen et al. 2010). Therefore, the down-regulating effects of exercise could be meaningful, especially in LCR rats, which suffer from this problem.
TFAM levels and the activity of SIRT1 changed in similar patterns with resveratrol and exercise intervention, suggesting a functional relationship between these proteins. TFAM is reported to increase the half life of and stabilize mtDNA, and thus increased levels of TFAM could have beneficial effects (Ikeuchi et al. 2005). However, the decreased levels of TFAM and mtDNA content with training, indicate an increased vulnerability of mtDNA.
Mitochondrial fission and fusion interact with each other. Fusion enables mitochondria to mix their contents within interconnected mitochondrial reticulum in order to minimize abnormalities. Fusion pathways have been shown to have beneficial roles in muscular atrophy (Romanello et al. 2010), endurance (Garnier et al. 2005), lifespan (Scheckhuber et al. 2007) and ROS production (Yu et al. 2006). In the present study, we have found that both exercise training and resveratrol enhance the levels of the main fusion protein, indicating beneficial affects of these interventions on the mitochondrial network.
Lon protease plays an important role in the quality control of mitochondrial proteins by degrading oxidatively modified proteins, and HSP78, an important mitochondria chaperone involved in LonP- associated degradation and quality control (Rottgers et al. 2002), and appears to be very well induced by exercise and resveratrol treatment in these LCR rats. Indeed, it has been suggested that long-term health conditions is accomplished by a complex network called vitagenes, a group of genes involved in preserving cellular homeostasis during stressful conditions (Calabrese et al. 2011; Calabrese et al. 2012). Vitagenes encode for heat shock proteins, sirtuins and Lon protease as well (Calabrese et al. 2006; Cornelius et al. 2013), and our currenet foundings suggest an improved level of quality control in the mitochondria with resveratrol and exercise training.
PNPase content was evaluated because it is important for the homeostasis of mitochondria (Chen et al. 2006) and can induce cellular senescence (Sarkar et al. 2005). Ablation of PNPase leads to a significant drop in enzymatic activities of respiratory complexes and decreases ATP production, leading to activation of AMPK phosphorylation (Chen et al. 2006). PNPase knockdown results in impaired mitochondrial membrane potential, which can readily affect fusion and fission (Chen et al. 2006). In the present study it was observed that exercise training increased PNPase content, suggesting that exercise played a role in the improvement of mitochondrial respiration.
In summary, the present data show that exercise intervention is an excellent tool to overcome genetic weaknesses by increasing endurance capacity. Exercise was able to overcome some of the weaknesses of mitochondrial systems through the activation of mitochondrial biogenesis. However, our data revealed that the AMPK-SIRT1-NAMPT-FOXO1 axis could be important for the limited aerobic endurance capacity of low running capacity rats. Resveratrol supplementation was not beneficial, in terms of aerobic endurance performance, mitochondrial biogenesis, or quality control.
Footnotes
ACKNOWLEDGEMENT
This work was supported by Hungarian grants from ETT 38388, OTKA (K75702) TAMOP-4.2.2/B-10/1-2010-0013 awarded to Z. Radák. The LCR rat model system was supported by grant R24 RR017718 from the National Center for Research Resources of the National Institutes of Health (to L.G.K. and S.L.B.), and National Institutes of Health grant RO1 DK077200 (to S.L.B.). The LCR model can be made available for collaborative study (contact: