
Abstract
The nervous system is of fundamental importance in the adaptive (hormesis) responses of organisms to all types of stress, including environmental “toxins”. Phytochemicals present in vegetables and fruits are believed to reduce the risk of several major diseases including cardiovascular disease, cancers and neurodegenerative disorders. Although antioxidant properties have been suggested as the basis of health benefits of phytochemicals, emerging findings suggest a quite different mechanism of action. Many phytochemicals normally function as toxins that protect the plants against insects and other damaging organisms. However, at the relatively low doses consumed by humans and other mammals these same “toxic” phytochemicals activate adaptive cellular stress response pathways that can protect the cells against a variety of adverse conditions. Recent findings have elucidated hormetic mechanisms of action of phytochemicals (e.g., resveratrol, curcumin, sulforaphanes and catechins) using cell culture and animal models of neurological disorders. Examples of hormesis pathways activated by phytochemicals include the transcription factor Nrf-2 which activates genes controlled by the antioxidant response element, and histone deacetylases of the sirtuin family and FOXO transcription factors. Such hormetic pathways stimulate the production of antioxidant enzymes, protein chaperones and neurotrophic factors. In several cases neurohormetic phytochemicals have been shown to suppress the disease process in animal models relevant to neurodegenerative disorders such as Alzheimer's and Parkinson's diseaess, and can also improve outcome following a stroke. We are currently screening a panel of biopesticides in order to establish hormetic doses, neuroprotective efficacy, mechanisms of action and therapeutic potential as dietary supplements.
HORMESIS AND ADAPTIVE STRESS RESPONSES
The term hormesis has been widely used in the toxicology field where it is defined as an adaptive response characterized by biphasic dose responses of generally similar quantitative features with respect to amplitude and range of the stimulatory response that are either directly induced or the result of compensatory biological processes following an initial disruption in homeostasis. In the context of responses of cells and organisms to environmental factors, hormesis refers to a process in which exposure to a low dose of a chemical agent or environmental factor that is damaging at higher doses induces a beneficial effect on the cell or organism. In biomedical sciences the terms “adaptive stress response” or “pre-conditioning” are often used instead of hormesis. At the cellular and molecular level hormesis involves activation of adaptive stress response pathways that result in increased expression of (for example) heat-shock proteins, antioxidant enzymes and anti-apoptotic proteins. In the present article we consider only the adaptive stress response aspect of hormesis in neurons—“neurohormesis”. For a consideration of the broader uses of the term hormesis we refer the reader to the recent article by Calabrese et al. (2007).
WHAT IS THE FUNCTION OF TOXINS PRESENT IN PLANTS?
The answer to this question is that plants evolved the ability to produce toxic substances and concentrate them in vulnerable regions (the skin, seeds and leaves) in order to dissuade insects and other organisms from eating and killing the plant. More than 100 such phyochemicals involved in plant defense have been identified (Trewavas and Stewart, 2003). At high concentrations they can be toxic (carcinogenic, neurotoxic and/or cardiotoxic) to mammals; however, in the amounts normally consumed, the concentration of the phytochemicals in the body is considerably below the toxic level. Phytochemical biopesticides include many different types of molecules—thiocyanates, polyphenolic compounds, nitriles and benzoxazinone, among others (Hansen et al., 2005). Additional specific examples include: alkaloids such as caffeine, strychnine and nicotine; flavonoids such as myricetin and rotenone; terpenoids such as camphor and farnesol; glucosinolates such as 2-pentylethyl isothiocyanate; indoles such as indole-3-acetonitrile; coumarins such as xanthotoxin; and phenylpropanols such as myristicin and eugenol. The fact that many of these phytochemicals can have beneficial effects on health is best established in studies of carcinogenesis where the phytochemicals can prevent the development of cancers at low doses, but are carcinogenic at high doses (Calabrese, 2005). However, neurons are the cells that are the “first line” responders to many phytochemicals. The importance of neurons in responses of organisms to potentially toxic phytochemicals is obvious when considering chemicals such as capsaicin (the chemical responsible for the “hotness” of hot peppers) or allicin in garlic which elicit an immediate stress response in sensory neurons in the mouth and associated structures. Similarly, psychoactive phytochemicals such as those in marijuana and peyote act primarily on neurons. However, neurons may also be particularly sensitive to phytochemicals absorbed through the gut including curcumin, sulforaphane and resveratrol (see below). We have therefore begun to investigate the molecular mechanisms underlying hormetic responses of neurons to phytochemicals.
SUBTOXIC DOSES OF PHYTOCHEMICALS CAN PROTECT NEURONS AGAINST INJURY AND DISEASE
As is the case with major diseases of other organ systems (cardiovascular disease, type 2 diabetes and cancers), data from epidemiological studies of human populations suggest that phytochemicals in fruits and vegetables can protect the nervous system against disease. For example, people who consume higher than average amounts of vegetables and fruits may be at reduced risk for Alzheimer's disease (Commenges et al., 2000; Staehelin, 2005). Because of their beneficial effects on the cerebral vasculature, phytochemicals may also reduce the risk of stroke (Flight and Clifton, 2006). Numerous studies of cell culture and animal models have demonstrated the efficacy of dietary supplementation with specific fruits or vegetables, or of extracts or specific chemical components of the fruits and vegetables. For example, dietary supplementation with blueberries: protected dopaminergic neurons against dysfunction and degeneration in a rat model of Parkinson's disease (Stromberg et al., 2005); improved learning and memory without affecting amyloid pathology in a mouse model of Alzheimer's disease (Joseph et al., 2003); and reduced brain damage and improved functional outcome in a rat model of stroke (Wang et al., 2005a). Drinking pomegranate juice reduced the amount of amyloid and improved behavioral deficits in a mouse model of Alzheimer's disease (Hartman et al., 2006), whereas apple juice concentrate prevented age-related impairment of cognitive function in mice (Tchantchou et al., 2005). Moderate consumption of red wine reduced amyloid pathology in a mouse model of Alzheimer's disease (Wang et al., 2006).
Considerable effort has been aimed at identifying specific molecules responsible for the health benefits conferred by plants. Many phytochemicals have been shown to exert neuroprotective actions in animal and cell culture models of neurological disorders, but in most cases their mechanism of action is unknown. Rather than attempting to review the now extensive literature on this topic, I will present examples of four different phytochemicals (sulforaphane, resveratrol, curcumin and the cannabinoid THC) which considerable evidence suggests have neuroprotective properties that likely involve a hormetic mechanism of action.
Sulforaphane is an isothiocyanate present in high amounts in broccoli (Fig. 1), Brussels sprouts and other cruciferous vegetables (Fahey et al., 1997). Several studies have reported neuroprotective effects of sulforaphane in animal models of both acute and chronic neurodegenerative conditions. In a rodent model of stroke sulforaphane administration reduced the amount of brain damage (Zhao et al., 2006). In a model of traumatic brain injury, sulforaphane treatment decreased brain edema (Zhao et al., 2005). Supforaphane also protected retinal pigment epithelial cells against photooxidative damage (Gao and Talalay, 2004). In a model of Parkinson's disease, sulforaphane protected dopaminergic neurons (Han et al., 2007).
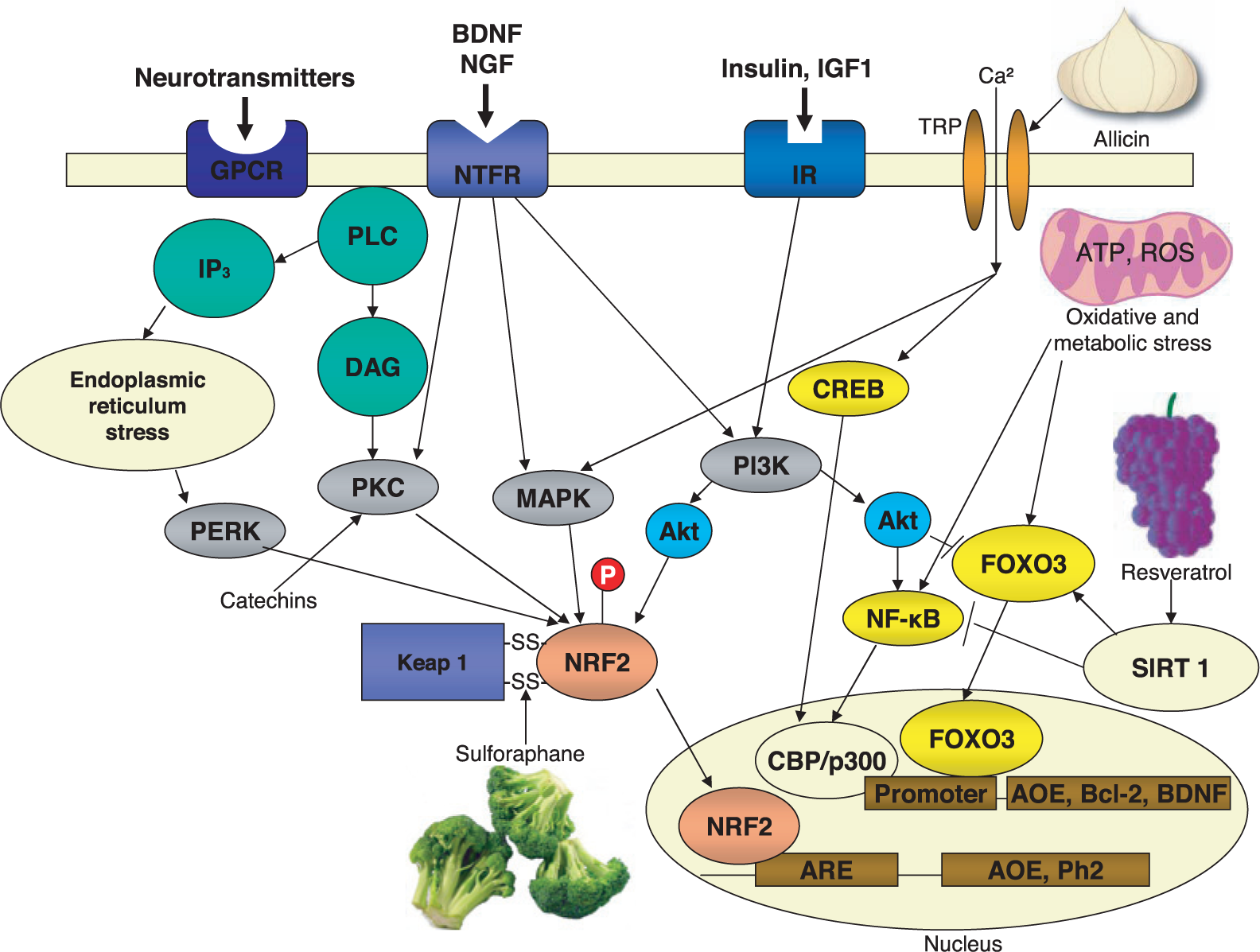
Examples of the molecular mechanisms by which phytochemicals activate hormetic stress response pathways. Neurons normally receive signals from multiple extracellular signals that activate adaptive cellular signaling pathways. For example, many neurotransmitters activate GTP-binding protein coupled receptors (GPCR), neurotrophic factors activate specific receptors (NTFR) and insulin and insulin-like growth factor 1 (IGF1) activate their receptors (IR). The receptors in turn activate kinase cascades including those that activate protein kinase C (PKC), mitogen-activated protein kinases (MAPK) and phosphatidylinositol-3-kinase (PI3K). Stress on the endoplasmic reticulum stress activates the PKR-like endoplasmic reticulum kinase (PERK). Downstream of PKC, MAPK, PI3K and PERK is the nuclear factor erythroid 2-related factor 2 (Nrf-2), a transcription factor that translocates to the nucleus and binds to the antioxidant response element (ARE) in genes that encode cytoprotective proteins such as antioxidant enzymes (AOE) and phase-2 proteins. Sulforaphane, a phytochemical present in relatively high amounts in broccoli, interacts with cysteine residues critical for the sulfhydryl-mediated interaction of the protein Keap with Nrf-2, thus freeing Nrf-2 so that it can translocate to the nucleus. Resveratrol, present in red grapes and wine, can activate a different hormetic pathway. Resveratrol interacts with the SIRT1 histone deacetylase which activates the transcription factor FOXO3 resulting in induction of genes encoding AOE, anti-apoptotic proteins such as Bcl-2 and neurotrophic factors such as BDNF. Other phytochemicals, including allicin from garlic and capsaicin from hot peppers, can activate specific ionotropic transient receptor potential (TRP) receptors in the plasma membrane resulting in Ca2+ influx; Ca2+, in turn, activates neuroprotective kinase cascades and the trancription factor CREB (cyclic AMP response element binding protein); CREB induces the expression of BDNF, an inter-cellular mediator of neurhormesis. Some phytochemicals may activate the transcription factor NF-κB, which is known to mediate adaptive cellular stress responses by inducing the expression of the AOE Mn-superoxide dismutase. BDNF, brain-derived neurotrophic factor; CBP, CREB-binding protein; DAG, diacylglycerol; IP3, inositol triphosphate; NGF, nerve growth factor; PLC, phospholipase C. This figure is modified from Mattson and Cheng, 2006.
Resveratrol (5-[(E)-2-(4-hydroxyphenyl)-ethenyl]benzene-1,3-diol) is a phytophenol present in high amounts in red grapes and wine. It has been suggested that resveratrol is largely responsible for the cardiovascular health benefits of red wine (Bradamante et al., 2004). Although there is as yet no direct evidence that dietary supplementation with resveratrol is effective in protecting against or treating neurological disease, an increasing number of studies in animal models have demonstrated neuroprotective effects of resveratrol. Administration of resveratrol to rats reduced ischemic damage to the brain in a model of stroke (Huang et al., 2001) and also protected spinal cord neurons against ischemic injury (Kaplan et al., 2005). Resveratrol can protect cultured neurons against nitric oxide-mediated oxidative stress-induced death (Bastianetto et al., 2000). Similarly, resveratrol protected dopaminergic neurons in midbrain slice cultures against metabolic and oxidative insults, a model relevant to Parkinson's disease (Okawara et al., 2007). Huntington's disease is caused by polyglutamine repeat expansions in the huntingtin protein resulting in aggregation and neurotoxicity of huntingtin. Resveratrol protected cells against the toxicity of mutant huntingtin in worm and cell culture models (Parker et al., 2005). In models relevant to Alzheimer's disease, resveratrol protected neuronal cells from being killed by amyloid β-peptide (Jang and Surh, 2003) and promoted the clearance of amyloid β-peptide from cultured cells (Marambaud et al., 2005).
Curcumin, the key component of curry spice, is a phenolic compound isolated from the roots of Curcuma longa (Joe et al., 2004). Dietary supplementation with curcumin protected rats against synaptic dysfunction and cognitive deficits in a model of traumatic brain injury (Wu et al., 2006). In a transgenic mouse model of Alzheimer's disease, dietary supplementation with curcumin (160–5000 ppm) decreased the accumulation of amyloid β-peptide, and markers of oxidative stress and inflammation in the cerebral cortex (Lim et al., 2001). Curcumin also exhibits anti-depressant-like effects in animal models of depression such as the forced swim test (Xu et al., 2005). Curcumin treatment was also effective in reducing neuronal apoptosis and improving functional outcome in an animal model of stroke (Wang et al., 2005b). Curcumin can directly protect cultured neurons against death induced by oxidative insults (Scapagnini et al., 2006).
THC (delta(9)-tetrahydrocannabinol) is the major psychoactive component of marijuana (Tanda and Goldberg, 2003). Beneficial effects of THC have been reported in studies of animal models of several neurodegenerative disorders. For example, THC attenuated the disease process and prolonged survival in a mouse model of amyotrophic lateral sclerosis (Raman et al., 2004). THC protected cultured neuronal cells (Chen et al., 2005a; Gilbert et al., 2007) and retinal neurons (El-Remessy et al., 2003) against excitotoxic damage. In addition, administration of THC to rats protected dopaminergic neurons against death induced by 6-hydroxydopamine in a model of Parkinson's disease (Lastres-Becker et al., 2005).
EXAMPLES OF HORMESIS PATHWAYS ACTIVATED BY NEUROPROTECTIVE PHYTOCHEMICALS
The conventional wisdom is that it is the direct antioxidant (free radical scavenging) activity of phytochemicals that confers their health benefits (Prior, 2003). In many cases, (resveratrol, lycopene and ginkgo biloba, for example) the chemicals do indeed possess antioxidant activity. However, these phytochemicals are only effective as antioxidants when they are present in supra-micromolar concentrations. Therefore, it is essentially impossible for humans to ingest the fruits and vegetables that contain these phyochemicals in amounts sufficient to achieve micromolar concentrations of the phytochemicals in cells in the body. Indeed, where examined, there is a biphasic dose-response relationship for many phytochemicals (low dose beneficial effects and high dose toxic effects), arguing against an exclusive antioxidant mechanism of action. Moreover, data from epidemiological studies and clinical trials have failed to demonstrate benefits of dietary supplementation with antioxidants such as vitamins E and C (Williams and Fisher, 2005). As described below, a more compelling case can be made for a hormetic mechanism of action of health-promoting phytochemicals.
Several phytochemicals have been shown to increase the resistance of neurons to various stressors by activating specific signal transduction pathways and transcription factors. For example, sulforaphane, curcumin and allicin can each activate the transcription factor Nrf-2 resulting in induction of genes regulated by the antioxidant response element (ARE) including antioxidant enzymes and phase-2 proteins (Dinkova-Kostova et al., 2002; Balogun et al., 2003; Chen et al., 2004). Apparently, sulforaphane and curcumin disrupt the interaction of Keap1 with Nrf-2 in the cytoplasm, which triggers the translocation of Nrf-2 to the nucleus (Fig. 1). Other findings suggest that sulforaphane may not cause Keap1 and NRF2 to dissociate, but instead sulforaphane inhibits the ubiquitin ligase activity of Keap1 which targets NRF2 for degradation (Zhang and Hannink, 2003). The gene encoding heme oxygenase-1 (HO-1), a redox stress-inducible protein known to protect cells against various types of stress, is one Nrf-2 target that likely contributes to the neuroprotective actions of sulforaphane and curcumin. While sulforaphane may have direct effects on neurons, recent findings suggest that it may also benefit neurons indirectly by activating the Nrf-2 pathway in astrocytes adjacent to neurons (Kraft et al., 2004). The latter is an example of transcellular hormesis in which one type of cell (astrocyte) responds to a stress by sending signals to adjacent cells (neurons) that protect them against stress.
The sirtuin–FOXO pathway, which is strongly linked to stress resistance and longevity, can be activated by resveratrol (Yang et al., 2007). Resveratrol may directly bind to the histone deacetylase SIRT1 resulting in the deacetylation and consequent activation of FOXO3 (Frescas et al., 2005). Activation of SIRT1 has been shown to increase neuronal survival via a FOXO3-mediated mechanism, at least under some conditions (Brunet et al., 2004). FOXO transcription factors may also mediate, in part, the hormetic effects of neurotrophic factors induced by stress including insulin-like growth factor-1 (IGF-1) and nerve growth factor (NGF) (Gan et al., 2005).
Another pathway that is a target for phytochemicals, and that likely mediates hormetic responses of neurons to a range of environmental stressors, involves activation of receptors for the excitatory neurotransmitter glutamate (Mattson, 2003). Low levels of activation of glutamate receptors can enhance synaptic plasticity and protect neurons against dysfunction and degeneration, in part by inducing the expression of neurotrophic factors (Marini et al., 2007). Experiments using cultured neurons and animal models have shown that activation of glutamate receptors by phytochemicals such as kainic acid can (at low concentrations) induce an adaptive cellular stress response that protects neurons against more severe and otherwise lethal conditions (Plamondon et al., 1999; Soriano et al., 2006). Activation of the glutamate receptors results in calcium influx, and calcium then activates kinases (protein kinase C and MAP kinases, for example) and transcription factors (CREB and NF-κB, for example) which induce the expression of cytoprotective proteins (anti-apoptotic proteins, antioxidant enzymes and calcium-binding proteins, for example) (Mabuchi et al., 2001; Maher, 2001; Mattson and Meffert, 2006). In addition, several phytochemicals that are well-known for their actions on the nervous system, may exert hormetic effects by activating transient receptor potential (TRP) calcium channels in the nerve cell membrane. Capsaicin (a noxious chemical in hot peppers), allicin (a pungent agent in garlic) and THC all activate TRP channels (Jordt et al., 2004; Macpherson et al., 2005). Cannabinoids are also known to activate specific G protein-coupled receptors (CB1 and CB2), and stimulation of those receptors may promote neuronal survival by stimulating Ca2+ influx and activating phosphatidylinositol-3-phosphate (PI3) kinase and mitogen-activated protein (MAP) kinases (Howlett, 2005; van der Stelt and Di Marzo, 2005). A possibly related neuroprotective mechanism of action of THC involves reactive oxygen species production and p38 kinase phosphorylation (Chen et al., 2005b).
Mitochondria play a critical role in supporting the survival and function of neurons by generating ATP required for the activities of various enzymes including ion-motive ATPases and kinases. However, mitochondria also produce high amounts of oxygen free radicals which can damage neurons. Some hormetic phytochemicals may target mitochondrial systems involved in energy and free radical metabolism. It is known that moderate levels of mitochondrial uncoupling can be neuroprotective (Chan et al., 2006; Liu et al., 2006), and that chemicals that induce uncoupling can, at low doses, exert hormetic effects on neurons (Mattiasson et al., 2003). Several neurohormetic phytochemicals have been shown to stabilize mitochondrial function under stress conditions including resveratrol (Lagouge et al., 2006). Resveratrol administration counteracted adverse effects of a high energy diet in mice by increasing mitochondrial number and improving insulin sensitivity (Baur et al., 2006). Allicin treatment increased the resistance of macrophages to apoptosis by stabilizing mitochondrial membranes (Cho et al., 2006).
PERSPECTIVE AND FUTURE DIRECTIONS
The efforts of Calabrese and colleagues (Calabrese and Baldwin, 2002; Calabrese, 2005) have revealed the widespread presence of hormetic dose-response curves in the fields of biology and toxicology. In essentially all types of cells and organisms studied so far, low doses of toxic agents can induce beneficial effects, presumably by activating evolutionarily conserved adaptive biological stress responses. Biomedical research has established hormetic effects of a variety of environmental factors, and has begun to elucidate the underlying cellular and molecular mechanisms. For example, the well-known health benefits of regular exercise, dietary energy restriction and cognitive stimulation are mediated by hormesis (Mattson et al., 2002; Martin et al., 2006; Vaynman and Gomez-Pinilla, 2006). The signal transduction pathways that mediate the neurohormetic effects of phytochemicals appear to overlap considerably with the pathways that mediate hormetic effects of more generalized types of stress (heat, metabolic and oxidative stress, for example) (Mattson et al., 2004; Arumugam et al., 2006; Ji et al., 2006). Such pathways result in the expression of genes that encode proteins that protect molecules in the cells against damage and dysfunction. Examples include: heat-shock proteins that bind to other proteins and thereby protect the proteins from damage (Lee et al., 1999); mitochondrial uncoupling proteins that decrease the production of free radicals (Liu et al., 2006); and growth factors that serve as inter-cellular mediators of adaptive stress responses (Lee et al., 2002; Maswood et al., 2004).
Knowledge of the range of phytochemicals that can activate hormetic responses in cells and organisms, and their mechanisms of action is limited. There is a need to develop high-throughput biochemical and cellular assays to identify and characterize neurohormetic phytochemicals. Detailed dose-response analysis and elucidation of the underlying molecular mechanisms of phytochemical-induced hormesis are essential for extending research to preclinical and clinical studies. There are several attractive features of phytochemical neurohormesis research in regards to applications to health care. For example, because they are natural products many phytochemicals can be readily moved to use in humans. On the other hand, there is considerable potential for the application of medicinal chemistry to phytochemicals with the aim of developing analogs with “better” properties (increased selectivity for a particular pathway, increased solubility, etc.). The coming years and decades promise to be exciting times in the emerging field of phytochemical hormesis.
Footnotes
ACKNOWLEDGMENTS
This work was supported by the Intramural Research Program of the National Institute on Aging.