
Abstract
Based on the cooperative virulence index (c.f.), the interaction effect between short-term heat pretreatment and avermectin on 2nd instar larvae of diamondback moth (DBM), Plutella xylostella (Linnaeus), was assessed. The results suggested that the interaction results between short-term heat pretreatment and avermectin on the tested insects varied with temperature level as well as its duration and avermectin concentration. Interaction between heat pretreatment at 30°C and avermectin mainly resulted in addition. Meanwhile, pretreatment at 35°C for 2 or 4 h could antagonize the toxicity of avermectin at lower concentrations, which indicated a hormetic effect occurred. The results indicate that cooperative virulence index (c.f.) may be adopted in hormetic effect assessment.
Keywords
INTRODUCTION
Hormesis, synonymous with other terms in other research fields (Calabrese 2008), is a dose-response phenomenon characterized by a low dose stimulation and a high dose inhibition, or vice versa depending on the endpoint measured. The hormetic dose response may be an inverted U-shape in the case of growth, longevity, or cognitive function or a J-shaped curve in the case of disease incidence (Calabrese and Baldwin 2001; 2003a; Calabrese 2005). Its underlying mechanism is overcompensation which occurs in the cell or tissue in response to low-dose chemicals, radiation, etc and results in a beneficial effect despite the fact it can be overcome at higher doses (Roberts 2001).
The hormesis phenomenon was first reported more than a century ago by Schulz (1887, 1888), who used yeast as his experimental model (Calabrese 1999). Up to now, it has been proven that hormetic dose-response relationships occur in males and females of numerous animal models in all principal age groups as well as across species displaying a broad range of differential susceptibilities to toxicants. The biological models are extensive, including plants, viruses, bacteria, fungi, insects, fish, birds, rodents, and primates, including humans. The spectrum of endpoints includes growth, longevity, numerous metabolic parameters, disease incidences (including cancer occurrence) and various performance endpoints such as cognitive functions, immune responses and many others (Calabrese and Baldwin 1998; 2003a; Calabrese and Blain 2005).
Quantitative features of the hormetic dose response indicate that the vast majority of cases display a maximum stimulatory response less than 2-fold, typically being 40%–60% greater than the control while the width of the stimulatory response is typically less than 100-fold, usually approximately 10-fold and the maximum stimulatory response is about 20%–25% of the traditional threshold dose for a toxic response (no observable adverse effect level, NOAEL) (Calabrese and Baldwin 1998; Calabrese 1999; 2004; Calabrese and Blain 2005).
The diamondback moth (DBM), Plutella xylostella (L.) (Lepidoptera; Plutellidae), is the major cosmopolitan pest of brassica and other crucifer crops all over the world. It distributes in areas of different climatic types including tropical, subtropical and temperate zones and can migrate everywhere (Chu 1986; Honda 1990; Honda et al. 1992; Chapman et al. 2002; Coulson et al. 2002). In tropical climates, DMB has 20 generations or more a year. As a result, the crop loss incurred by DBM may be as high as 90% (Verkerk and Wright 1996) and only few 4th instar larvae on a cabbage can make it unsaleable (Shelton et al. 1983; Maltais et al. 1998). The total annual cost for DBM control throughout the world surpasses one billion US dollars (Talekar and Shelton 1993; Roux et al. 2007). For decades, insecticide use has been the most important control targeting the 2nd and 3rd instar larvae. Presently, avermectin is widely used in DBM control both in- and outside China (Sayyed et al. 2004; Luo et al. 2008).
Temperature is the most important factor in relation to the occurrence and distribution of DBM. Prior studies suggest that, for growth and development of DBM, the most favorable temperature is 25°C and temperatures higher than 30°C or lower than 20°C are harmful (Ma and Chen 1993; Dan et al. 1995; Shirai, 2000; Liu et al. 2002).
High temperature and insecticides both can threaten the survival of DBM, but what is the interaction effect between them? Does short-term heat pretreatment always improve the toxicity of the subsequently administered insecticides to DBM or does the prior heat treatment have a hormetic effect on the tolerance of DBM to the subsequently administered insecticide? The purposes of this study were to answer these two questions. We did find that short-term high temperature pretreatment did not always increased the toxicity of avermectin, an important DBM control insecticide to the 2nd instar larvae of DBM, and short-term high temperature pretreatment did have a hormetic effect on the tolerance of the 2nd instar larvae of DBM to avermectin.
MATERIALS AND METHODS
Chemicals
Avermectin crude chemical (95.0% purity) was provided by Zhejiang Hisun Pharmaceutical Co., Ltd. TritonX-100 (a nonionic detergent) was bought at a local Fuzhou chemical market. Avermectin was diluted half serially into 4.50, 2.25, 1.13, 0.56 and 0.28 mg/L with distilled water containing 0.1% (vol:vol) Triton X-100 for bioassay.
Insect stock culture
Pupae of DBM were originally collected from the vegetable fields in a suburb of Fuzhou City, Fujian Province, People's Republic of China. The pupae were put into a plastic soft drink bottle of 1.5L with lots of small round holes (1mm dia) in the lower side and 4 bigger round holes (10mm dia) in the upper side of the wall and 20 couples placed in each. After pupation, the female and male adults mated and the eggs were laid to the inside wall. The hatched 1st instar larvae dropped through the holes onto the leaves of cabbage plants, Brassica oleracea L. var. capitata L, grown in the pots 20 cm under the bottle. The insectary temperature was set at 25±1°C, 60%–70% RH with a photoperiod of 12:12 (L:D). Adults were fed with a 10% sugar solution saturated in cotton placed in the bigger holes in the upper side of the bottle and the cotton was changed twice everyday. The insects were reared for more than 30 generations without exposure to any insecticides. For experiments, the same day-aged 2nd instar larvae (body length about 2mm) were used.
Short-term heat pretreatment
For heat pretreatment, the temperature was set at 30°C and 35°C, respectively. Ten insects were released on each disc in an individual petri plate (100mm dia) covered with wet filter paper of same size at the bottom. The heat pretreatment duration included 0h, 2h, 4h, 8h and 12h, respectively. The heat pretreatment was conducted in a digitized biochemistry incubator, produced by Hankang electronic Co, Ltd, Jintan City, Jiangsu Province, People's Republic of China. Soon after heat pretreatment, the petri plates were removed to 25±1°C, 60–70% RH with a photoperiod of 12:12 (L:D) for avermectin bioassay.
Bioassay
Cabbage leaf disc dip method of bioassay as described by Tabashnik et al. (1987) was adopted in the present studies. Cabbage leaves were first washed with distilled water and dried for about 1h at room temperature. Cabbage leaf discs (10 mm dia) were then cut with a metal punch and dipped into a test solution prepared with distilled water containing 0.1% TritonX-100 to facilitate uniform treatment with active ingredient for about 5s. For control, the leaf discs were dipped in distilled water containing 0.1% TritonX-100 without active ingredients for the same period. The leaf discs were placed slanting for about 2 minutes over a blotting paper in a tray to drain excess solution and then flattened to dry the test solution for about 2h at room temperature. Finally, 2 leaf discs were put into one petri plate to feed the insects. Each concentration had 3 replications and each replication contained 10 insects (1 petri plate). Larvae were allowed to feed on the treated leaf discs for 48h at 25°C before being checked for mortality (Mohan and Gujar, 2003). An insect was regarded as dead if it had no response to a gentle touch of tweezers.
Data analysis
Data obtained from the experiments were analyzed using analysis of variance (P<0.05) (Proc ANOVA; Tang and Feng 1997). Treatment means were compared by Tukey's F test, accepting significant differences at P=0.05 (Tang and Feng 1997). The mortality data were transformed by the arcsine square root prior to significance analysis (Southwood and Henderson 2000) and were averaged within replications for each treatment (Sokal and Rohlf 1995; Yin et al. 2008).
Concentration-mortality data were analyzed by probit analysis using DPS (Tang and Feng 1997). Mortality rates were corrected using Abbott's formula (Abbott, 1925) for each probit analysis.
The median lethal concentration (LC50) in terms of mg active ingredient/l was estimated by subjecting mortality data to the maximum likelihood program of probit analysis (Tang and Feng, 1997). This program has a provision for control mortality. Tukey's F test was also used to compare the differences among the LC50s.
Cooperative virulence index (c.f.) was calculated with the formula proposed by Mansour et al. (1966), which was, c.f.=(real mortality-theoretical mortality)/theoretical mortalityX100, where theoretical mortality=the corrected mortality caused by heat treatment alone + the corrected mortality caused by avermectin alone - the product between them. The interaction result was assessed as, ‘synergism’ when c.f. was greater than or equal to 20; ‘addition’ when c.f. was between −20 and 20; and ‘antagonism’ when c.f. was less than or equal to −20.
RESULTS AND ANALYSIS
Interaction between short-term heat pretreatment at 30°C and avermectin on 2nd instar larvae of DBM, Plutella xylostella (Linn).
Sole heat treatment did not increase the mortality of the tested insects for no significant differences in mortality were found among the treatments (Tr1, Tr2, Tr3 and Tr4 in Table 1) and control (P>0.05). For example, the mortality in the control was 10.00% but that in Tr4, where the tested insects were treated at 30°C for 12 h, was only 13.33% and the difference was not significant(P>0.05). Meanwhile, mortalities in Tr1 and Tr3 were a little lower than that in the control, despite the fact that the differences were insignificant.
Combined toxicity between short-term heat pretrement at 30°C and avermectin on 2nd instar larvae of DBM, Plutella xylostella (Linn.)

Note: Data followed with the same lower case letter are not significantly different (P=0.05, Tukey's F test). Same as below.
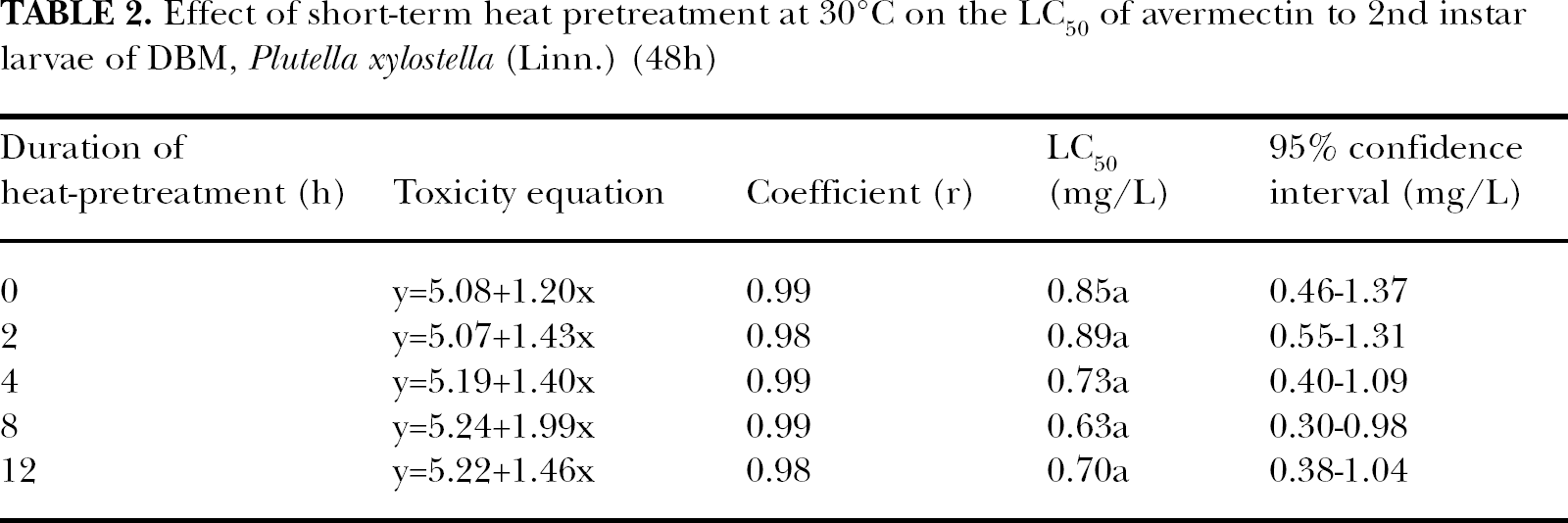
At higher avermectin concentrations, short-term heat pretreatment at 30°C seemed to increase its toxicity. For example, in the treatments with avermectin at 4.50 mg/L (Tr25, Tr26, Tr27, Tr28 and Tr29 in Table 1), the mortality was 82.33% in Tr25 (in which the insects experienced no prior heat exposure), but that increased to 93.00% in Tr 29 (in which the insects were pretreated at 30°C for 12h). However, the effect was not significant (P>0.05) and did not cause a significant change in the LC50 of avermectin to the tested insects (shown in Table 2).
Effect of short-term heat pretreatment at 30°C on the LC50 of avermectin to 2nd instar larvae of DBM, Plutella xylostella (Linn.) (48h)
The cooperative virulence index (c.f.) indicated that the interaction between short-term heat pretreatment at 30°C and avermectin mainly resulted in addition and the only antagonism was found in Tr6 (Table 3), where the heat pretreatment duration was 2 h and avermectin concentration was 0.28mg/L.
Assessment of interaction effect between short-term heat pretreatment at 30°C and avermectin on 2nd instar larvae of diamondback moth, Plutella xybstella (Linn)

Interaction between short-term heat pretreatment at 35°C and avermectin on the 2nd instar larvae of DBM, Plutella xylostella (Linn).
Similar to that at 30°C, heat treatment at 35°C alone did not significantly increase the mortality of the tested insects, either (P>0.05, Table 4). The mortality was 14.00% in control and 17.00% in Tr4 in which the insects were treated at 35°C for 12h. But in Tr2, Tr3 and Tr4, the mortality data were all a little lower than that in control.
Combined toxicity between short-term heat pretrement at 35°C and avermectin on 2nd instar larvae of DBM, Plutella xylostella (Linn.)
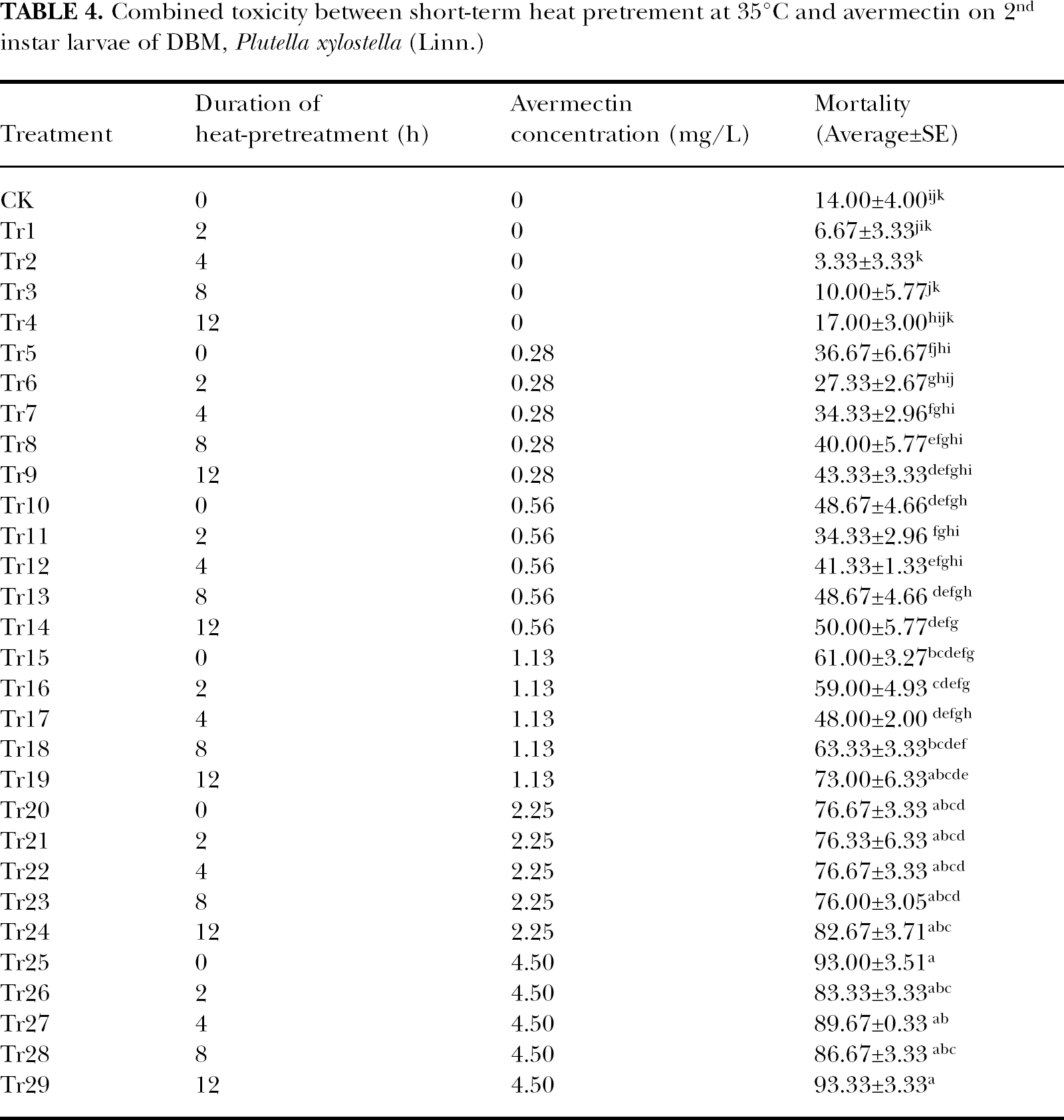
Compared with avermectin treatment only, short-term heat pretreatment at 35°C for 2or 4h could also decrease the mortality of the tested insects. For instance, when avermectin concentration was 1.13mg/L (Tr15, Tr16, Tr17, Tr18 and Tr19), the mortality in Tr15 (where the insects were only treated with avermectin) was 61.00%, but that in Tr16 and Tr17 (where the insects were pretreated at 35°C for 2 and 4h, respectively) was 59.00% and 48.00% respectively. But just as that at 30°C, the effect was not significant. However, when the duration of the heat pretreatment was extended to 8h or 12h, the mortality was increased. Also, take the concentration of avermectin at 1.13 mg/L as an example, when the duration of heat pretreatment was 8h (Tr18), the mortality was 63.33% and 73.00% when heat pretreatment duration was 12h (Tr19). Both were a little higher than that in Tr15 and again the differences were not significant (P>0.05). The effect of short-term heat pretreatment on the avermectin tolerance of the tested insects could also be seen in Table 5. Although, after heat pretreatment, the LC50 of avermectin to the tested insects fluctuated, no significant differences were found (P>0.05).
Effect of short-term heat pretreatment at 35°C on the LC50 of avermectin on the 2nd instar larvae of DBM, Plutella xylostella (Linn.) (48h)

As was indicated in Table 4, short-term heat pretreatment at 35°C could decrease the toxicity of avermectin by a small amount, and c.f between heat pretreatment for these 2 durations and avermectin were all below zero which suggested that the real mortality was lower than the theoretical one. In some treatments (Tr6, Tr11, Tr12 and Tr17), that even resulted in antagonism (Table 6). When the duration of heat pretreatment was extended to 8h (Tr8, Tr13, Tr18, Tr23 and Tr28), c.f. was below zero in Tr23 or Tr28, but above zero in the other 3 treatments. However, interaction in all these treatments all resulted in addition. Meanwhile, extending the heat pretreatment duration to 12h (Tr9, Tr14, Tr19, Tr24 and Tr29) and the c.f was above zero in all treatments except Tr14, where it was −1.27, indicating that most real mortality data were greater than the theoretical ones and that synergism even occurred in Tr19.
Assessment of interaction effect between short-term heat pretreatment at 35°C and avermectin on 2nd instar larvae of diamondback moth, Plutella xybstella (Linn)

DISCUSSION
Previous studies have proven that the most favorable temperature for growth and development of DBM is 25°C. Temperature higher than 30°C especially when it reaches 35°C or above is harmful, and even might cause the extinction of the laboratory population (Ma and Chen 1993; Dan et al. 1995; Shirai, 2000; Liu et al. 2002). For instance, the survival of the 2nd instar larvae is 88.77% reared at 25°C and 84.34% at 30°C, but only 54.17% at 35°C (Dan et al. 1995). But if the high temperature only lasts for a comparatively short period, the overcompensation is probable which means hormesis might occur. However this has not been proven before.
In this study, sole short-term heat treatment did not significantly change the mortality of the tested insects either at 30°C or 35°C (Table 1 and Table 4). But in some treatments, which were Tr3 in Table 1 and Tr3, Tr4, Tr5 in Table 4, heat treatment did decrease the mortality despite the fact that the differences were insignificant (P>0.05).
Of the known hormesis phenomena, about 80% of the maximum stimulatory or inhibitory responses were less than twice the control value and most were approximately 30%–60% greater than the control response (Calabrese 2004). This difference was always not significant by means of the often used statistical analysis method (Calabrese and Baldwin, 1998; Calabrese, 2004). In our work, compared with blank control and expressed in proportion, sole short-term heat treatment caused a decrease of mortality by 30.0%–66.7% at 30°C (Tr1 and Tr3 in Table 1) and 28.6% to 76.2% at 35°C (Tr1, Tr2 and Tr3 in Table 4). These are within the range of the hormetic effect, but more work still needs to be done to determine it.
Hormesis is an evolutionary natural selection process involving toxicological mechanisms as part of a strategy to enhance survival to low levels of stressor agents (Calabrese and Baldwin, 2001; Calabrese, 2008). Up to now, hormesis is found to have two types of roles. First, it helps the organism to recover from the damage caused by xenobiotic stressors (Calabrese and Baldwin 2003b). If the decrease of the mortality of the 2nd instar larvae of DBM shown in Table 1 and Table 4 is determined as a hormetic effect, it could be the result of the effect of the stressors. Second, it helps the organisms to display a heightened resistance to a subsequent and more massive exposure (Calabrese, 2008). Moreover, because for all low or modest level stresses, organisms have similar response mechanisms, hormesis can not only help the organisms to defend against the same experienced injury but also other unexperienced injuries. The latter is widely known as cross protection (Stebbing 1981; Calabrese and Baldwin 2003b). Cross protection has also been found in the interaction between short-term heat treatment and insecticides. For example, after treating 4th instar larvae of mosquitoes, Anopheles stephensi and Aedes aegypti normally, with high but sub lethal temperatures, cross-tolerance to propoxur (a carbamate insecticide) was conferred. The improvement of heat shock protein expression was suggested to be the underlying mechanism (Patil et al. 1996). The antagonism effect of the short-term heat pretreatment at 35°C on the toxicity of avermectin to 2nd instar larvae of DBM at some concentrations (Table 6) should be the result of cross-tolerance. But whether the underlying mechanism is also because of the more expression of heat shock protein still needs to be studied.
The hormetic effect may not be significant by means of the often used statistical analysis method (Calabrese and Baldwin, 1998; Calabrese, 2004) and it was not found in the present study. For although short-term heat pretreatment has changed the mortalities in some treatments, most were insignificant. This may also be the reason why no significant changes were found in the LC50s of avermectin to the tested insects (Table 2 and Table 5). A new assessment method for hormetic effect is necessary. Cooperative virulence index (c.f.) is widely used in the assessment of interaction effects between pesticides based on the resultant mortality data without taking the toxicity mechanisms into consideration. It seems reasonable to adopt it in the assessment of interaction effects between short-term heat pretreatment and insecticides. Just as the hormetic definition has indicated, the hormetic effect is dose dependent and nonlinear (Calabrese and Baldwin 2001; 2003a; Calabrese 2005). Also the hormetic effect varies with the level of the subsequent injuries, for it can be overcome by injury at higher levels (Roberts 2001). This is why the different assessment results were found in different treatments (Table 3 and Table 6). Integrate all these factors, the interaction results between short-term heat pretreatment at 35°C and avermectin shown in Table 6 may be explained as follows. When the duration of heat pretreatment was set at 2h or 4h, the hormesis effect was comparatively high, and would antagonize the toxicity of avermectin at some concentrations. In other treatments, although the interaction did not result in antagonism, heat pretreatment decreased the toxicity of avermectin for c.f was below zero except in Tr22. But when the duration was extended to 8 or 12h, hormesis became low or even the harmful effect came into being for c.f was above zero and in Tr19 the interaction even resulted in synergism.
The differences between the effect of short-term heat pretreatment at 30°C on the subsequent toxicities of avermectin at different concentrations and that of heat pretreatment at 35°C was probably because of the hormetic effect at 30°C was comparatively low and unstable and it could not agonize the toxicity of avermectin at most concentrations. This can be seen in Table 1 and Table 4. Expressed in proportion, heat pretreatment at 30°C could decrease the mortality by 30% when the duration was 2h (Tr1 in Table 1) and 67.7% when the duration was 8h (Tr3 in Table 1). However, when the temperature was set at 35°C, the decrease in mortality was 52.36%, 83.36% and 28.57% when the duration was 2, 4 and 8h, respectively (Tr1, Tr2 and Tr3 in Table 4). Other evidence is that 2nd instar larvae reared at 30°C for the whole stage had the similar survival as that at 25°C, but that at 35°C is significantly lower (Dan et al. 1995) which indicates that long-term 30°C is less harmful than 35°C.
Footnotes
ACKNOWLEDGEMENTS
This research work was supported by the Natural Science Foundations of Fujian Province (Project No. B0410015, 2004J010 and 2007F5021). Cordial thanks are given to Dr. Edward J. Calabrese (University of Massachusetts, USA) and Dr. Shusheng Liu (Zhejiang University, People's Republic of China) for their kindness of sending their related publications. Also, the authors deeply appreciate Mrs. Carol Gellrich and Dr. David Boyle in Canada and three anonymous reviewers for their valuable suggestions on this manuscript.