
Abstract
CCN2/connective tissue growth factor (CCN2/CTGF) is a critical signaling modulator of mesenchymal tissue development. This study investigated the localization and expression of CCN2/CTGF as a factor supporting angiogenesis and chondrogenesis during development of secondary ossification centers in the mouse tibial epiphysis. Formation of the secondary ossification center was initiated by cartilage canal formation and blood vessel invasion at 7 days of age, and onset of ossification was observed at 14 days. In situ hybridization showed that CCN2/CTGF mRNA was distinctively expressed in the region of the cartilage canal and capsule-attached marginal tissues at 7 days of age, and distinct expression was also observed in proliferating chondrocytes around the marrow space at 14 days of age. Immunostaining showed that CCN2/CTGF was distributed broadly around the expressed cells located in the central region of the epiphysis, where the chondrocytes become hypertrophic and the cartilage canal enters into the hypertrophic mass. Furthermore, an overlapping distribution of metalloproteinase (MMP)9 and CCN2/CTGF was found in the secondary ossification center. These findings suggest that the CCN2/CTGF is involved in establishing epiphyseal vascularization and remodeling, which eventually determines the secondary ossification center in the developing epiphysial cartilage.
Keywords
CCN2/
Previously, a cDNA specific in a human chondrosarcoma-derived cell line, HCS-2/8 (Takigawa et al. 1989), was cloned by differential display PCR. After the identification of the cDNA clone as
The secondary ossification center is a feasible target for the study of the integrated function of CCN2/CTGF in mesenchymal tissues, because its formation involves various types of CCN2/CTGF producing/target cells (i.e., chondrocytes, osteoblasts, vascular endothelial cells) that support epiphyseal vascularization and endochondral ossification process. The formation of the secondary ossification center begins morphologically with cartilage canal formation, which is associated with peripheral vascular proliferation that invaginates from the perichondrium. This process provides a physical and biological background to support and nourish the joints that are exposed to mechanical stress immediately after birth, whereas the primary ossification center is engaged in longitudinal bone growth. Therefore, these two ossification centers play distinct roles in skeletal development through similar, but different, biological processes. Notably, subsequent ossification, which is characterized by disintegration of hypertrophic chondrocyte and formation of bone trabeculae accompanied by vascular invasion, takes place adjacent to preexisting cartilage canals (Karaplis 2002). A previous study presented evidence that CCN2/CTGF was a central growth factor in endochondral ossification, acting in paracrine and matricrine manners in the growth cartilage. However, the functional significance and role of CCN2/CTGF during earlier events, such as the formation of the secondary ossification center, are not known yet.
This study investigated the temporal and spatial gene expression and localization of CCN2/CTGF during the formation of the secondary ossification center and determined that CCN2/CTGF collaborates with other molecules in establishing epiphyseal vascularization. These data suggest that CCN2/CTGF is therefore involved in establishing epiphyseal vascularization, as well as determining and developing the secondary ossification center in the mouse tibia.
Materials and Methods
Tissue Preparation
To prepare samples for immunostaining and in situ hybridization, 20 mice at 1, 7, 14, 21, and 28 days postnatal were killed. Knee joints with the surrounding soft tissues were dissected and fixed with 4% paraformaldehyde in 0.1 M PBS (pH 7.4), decalcified with 10% EDTA, and embedded in paraffin according to an established procedure. Longitudinal sections of 7.0 μm thickness were cut on a microtome and mounted on slide glasses. The sections were stored at 4C until subsequent use. The Animal Committee of Okayama University Dental School approved all of the procedures.
Immunostaining
Two different antibodies were used as primary antibodies: One was a polyclonal anti-CCN2/CTGF serum that was raised in rabbits by immunization with a synthetic peptide of CCN2/CTGF, as described previously (Kubota et al. 2001). The other is a rabbit polyclonal antibody against mouse MMP9, which was commercially available (AB19047; Chemicon International, Temecula, CA). The paraffin sections were soaked in xylene to remove the paraffin and dehydrated in a graded series of ethanol (100% to 70%). After deparaffinization, the sections were rinsed in tap water. Endogenous peroxidase activity was quenched by immersion in 3% hydrogen peroxide in methanol for 30 min. After being washed in 0.1 M Tris-buffered saline (TBS; pH 7.4) for 5 min, the sections were incubated with 10% normal serum of the same species that produced the primary antibody for 1 hr at room temperature to eliminate nonspecific binding. Thereafter, the sections were incubated overnight with a 1:100 dilution of each primary antibody at 4C. After a wash in TBS with 0.1% Tween 20, the sections were incubated with a peroxidase-conjugated anti-goat IgG as a secondary antibody (PI-1000; Vector, Burlingame, CA) for 60 min at room temperature and washed in TBS. Finally, color development was performed using 3,3′-diaminobenzidine tetrachloride (Dojindo; Tokyo, Japan). The sections were also counterstained with hematoxylin and mounted. Controls samples were processed with the omission of the primary antibody or incubation with a normal rabbit serum (Dako; Tokyo, Japan) instead.
Confocal Laser-scanning Microscopy
The sections were also scanned for fluorescence using Radiance 2100K2 model (Bio-Rad Japan; Tokyo, Japan) to detect emission by nuclear staining (Sytox Green) or by antibody staining (Alexa Fluor). Sections were deparaffinized in xylene and rehydrated through graded ethanol to water, blocked in a blocking buffer (5% non-fat milk in TBS), and incubated overnight with a 1:100 dilution of primary anti-CCN2/CTGF antibody at 4C, and subsequently with a 1:800 dilution of goat anti-rabbit secondary antibody (IgG) conjugated to Alexa Fluor 568 (Molecular Probes; Eugene, OR) for 40 min at room temperature, after which samples were counterstained in Sytox Green (Molecular Probes) for 20 min at room temperature and washed in TBS. Confocal laser-scanning microscopy was performed using an argon/krypton laser (excitation wavelength, 488 and 568 nm). The images were enhanced using the Adobe Photoshop software program (San Jose, CA).
Preparation of Probes for In Situ Hybridization
For the generation of sense and antisense mouse CCN2/CTGF probes, a CCN2/CTGF cDNA (10kbp) that included the full length of the open reading frame, which was obtained from restriction enzymatic digestion and the fragment purification of pcDNA3.1(−)-ctgf (Kubota et al. 2000), was subcloned into a pGEM-3Zf plasmid (Promega; Madison, WI). For the generation of sense and antisense mouse collagen type X (α1) probes, a cDNA of mouse collagen type X α1 (640 bp), which was obtained from restriction enzymatic digestion and fragment purification of a PCR product, was subcloned into pGEM-7Zf(+) plasmid (Promega). Using these plasmids as templates, digoxigenin (DIG)-labeled sense and antisense riboprobes were synthesized with T7 or SP6 polymerase and DIG-11-UTP using a DIG-RNA labeling kit (Roche Molecular Systems; Mannheim, Germany). The synthesized probes were processed into pieces measuring ∼150 bp in length by hydrolysis for use in in situ hybridization.
Histological features and the localization of CCN2/connective tissue growth factor (CTGF) mRNA and protein in the tibial epiphysis at 1 day of age. Serial sections were stained with hematoxylin and eosin (
In Situ Hybridization
In situ hybridization was performed, using the DIG-labeled sense and antisense CCN2/CTGF riboprobes as previously described (Nakanishi et al. 1997). In brief, deparaffinized and rehydrated sections were incubated with 10 μg/ml proteinase K for 15 min at 37C. The sections were hybridized for 16 hr at 50C. The hybridization solution contained 50% deionized formamide, 1× Denhardt solution, 10% dextran sulfate, 0.25% SDS, 1 mM EDTA, 600 mM NaCl, and 1 μg/ml labeled probe. Hybridization was performed in a humidified chamber. After hybridization, the sections were washed and immunoreacted with a diluted anti-digoxigenin Fab fragment conjugated with alkaline phosphatase (Roche) at room temperature for 1 hr. The sections were stained with nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolyphosphate at room temperature. Under the conditions applied, the control hybridization incubated with the sense probe showed no detectable signals.
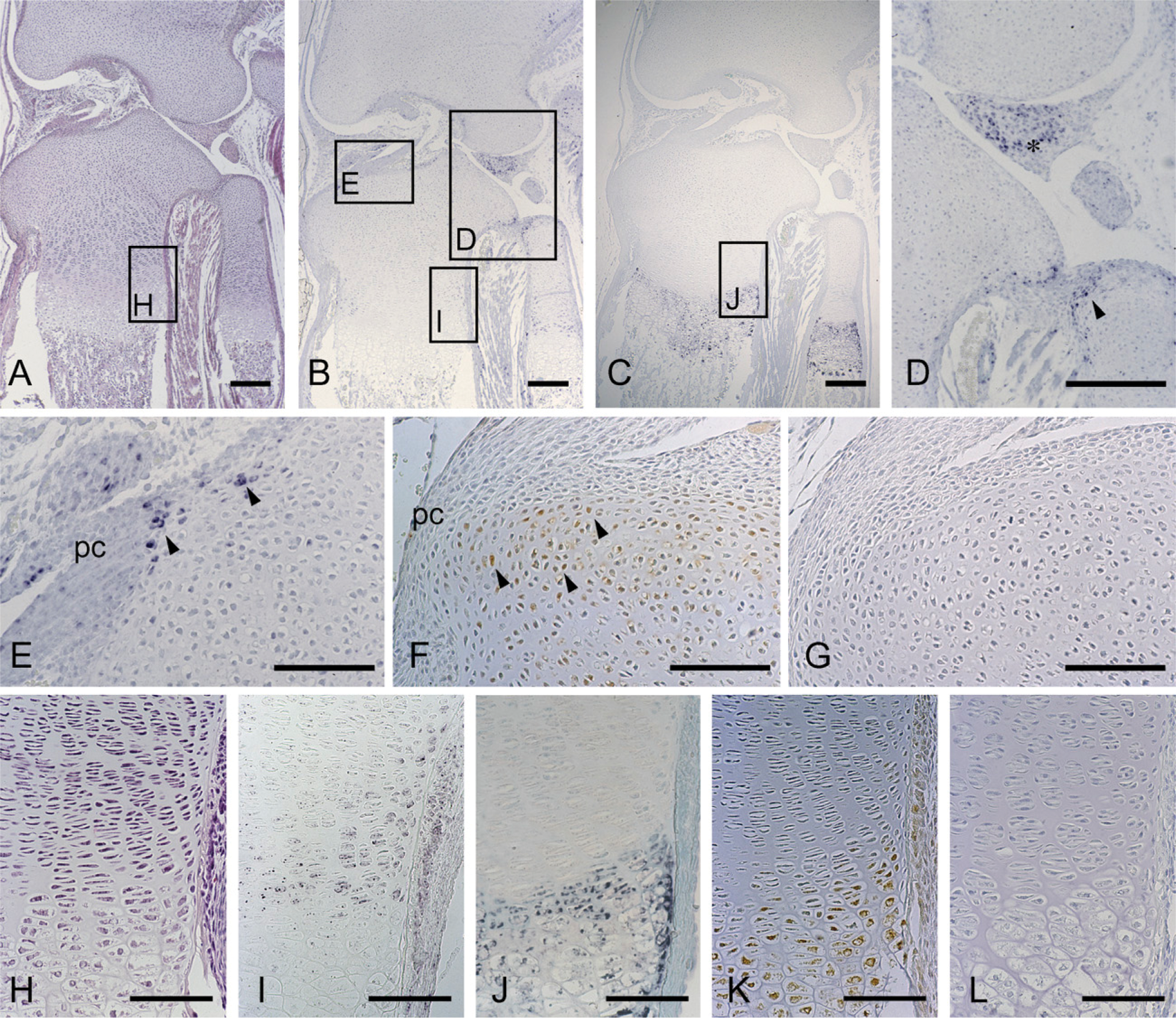
Histological features and the localization of CCN2/CTGF mRNA and protein in the tibial epiphysis at 7 days of age; sections facing toward the canal blind end. Serial sections were stained with hematoxylin and eosin (
Results
Developmental Time Course of the Secondary Ossification Center in Mouse Tibiae
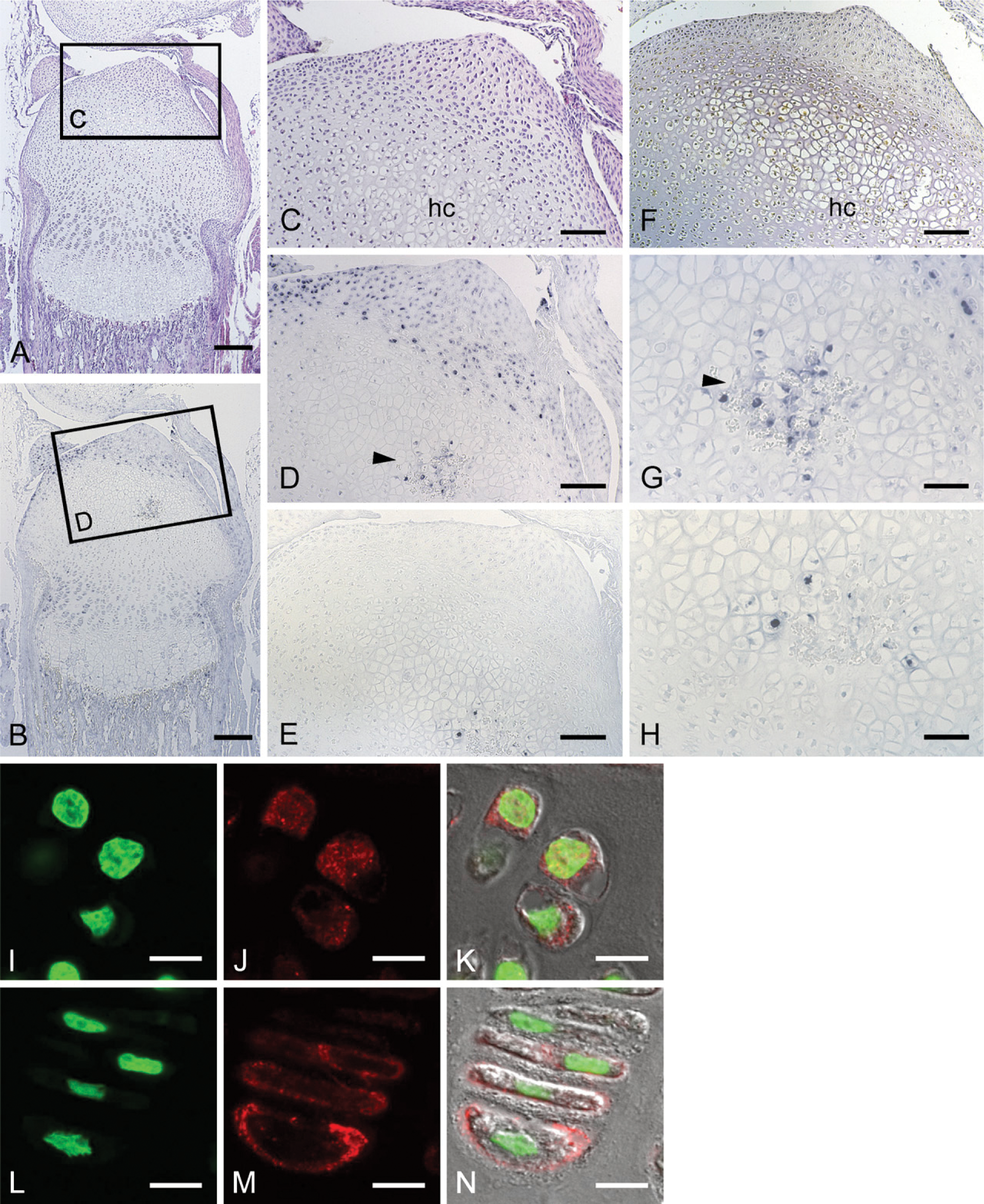
Initially, the secondary ossification center formation process was monitored before the study of stage-specific distribution of CCN2/CTGF, after a postnatal time course. At 1 day, the developing tibial epiphysis was mostly composed of resting cartilage (Figure 1A), and a cartilage canal was not yet present. However, in the superficial layer, chondrocytes adjacent to the capsule attachment tended to be proliferating. At 7 days of age, the chondrocytes in the central region of the epiphysis became hypertrophic, expressing type X collagen (Figure 2E). At the same time, the cartilage canal formed by an invagination from the perichondrium was observed with the growth of the epiphysis in the horizontal direction (Figure 3A). At this stage, fibrovascular proliferation started from the perichondrium close to the capsule attachment. The canal space was continuously lined by endothelial cells and was filled with a number of immature vasculatures (Figure 3D). Hypertrophic chondrocytes and resorption cartilage appeared around the cartilage canal and were particularly prominent near the canal blind end. At 14 days of age, the marrow space increased in size with the resorption of the hypertrophic chondrocytes, and a secondary ossification center was observed with the formation of thin bone trabeculae (Figure 4A). Along the boundary of the articular cartilage and secondary ossification center, an immature but polarized growth plate-like structure with continuously aligned chondrocytes was observed, but no such structure was observed at the metaphysial side of the marrow space (Figure 4B). Consistent with this histological feature, chondrocytes expressing type X collagen were observed only along the articular side of the ossification center (Figure 4J). Finally, at 28 days of age, the ossification center expanded, and the walls of marrow cavity were formed by thick bone trabeculae (Figure 5A). Groups of hypertrophic chondrocytes in the central region of the epiphysis were mostly replaced by bone marrow except at the marginal region of both lateral sides. At this stage, the formation of articular cartilage and the epiphyseal plate can be eventually clearly distinguished.
Expression of ccn2/ctgf and Distribution of CCN2/CTGF During Cartilage Canal Invasion
The spatiotemporal patterns of CCN2/CTGF expression were examined during the formation of the ossification center by in situ hybridization. Initially (1 day of age),
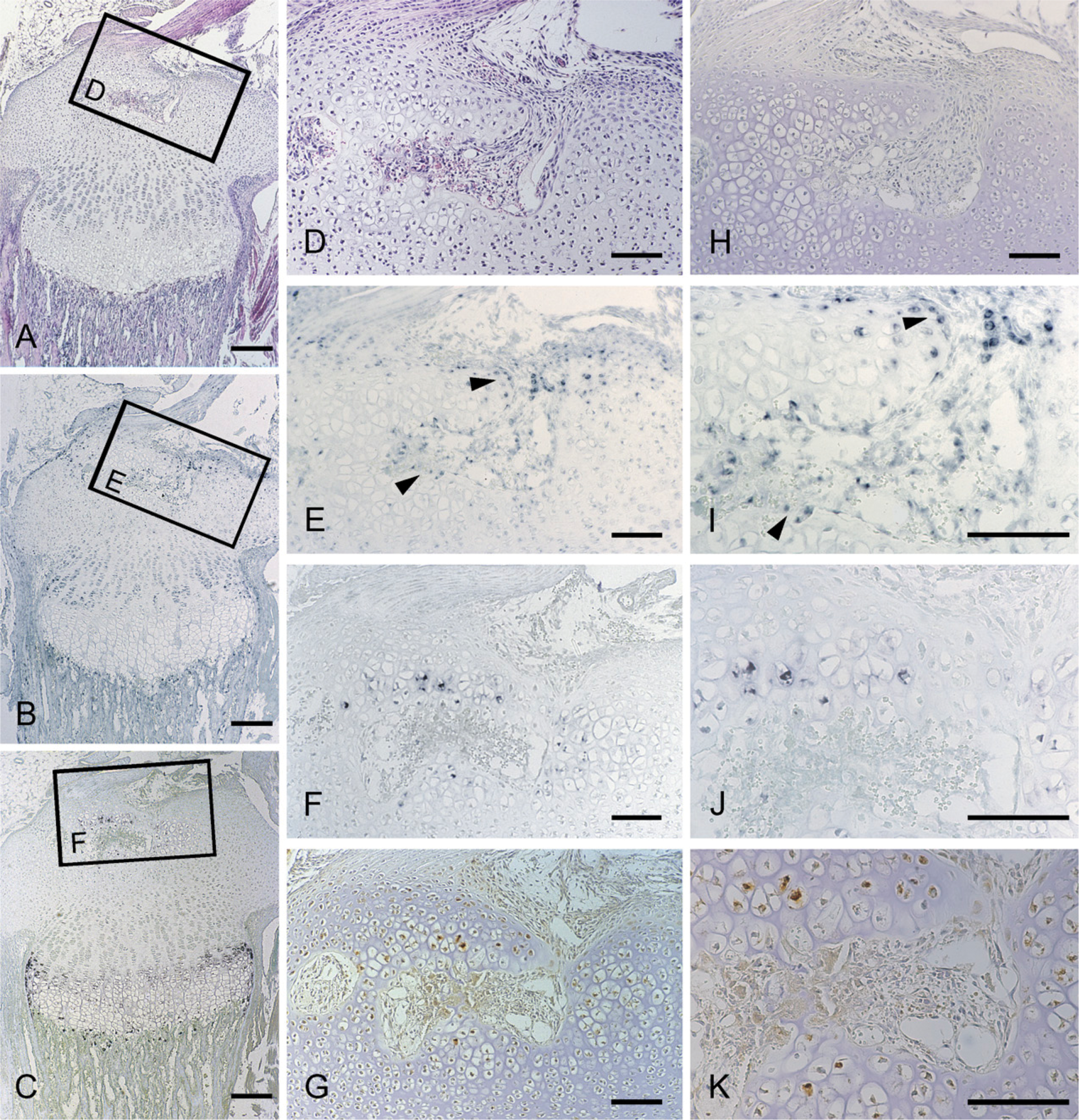
The histological features and localization of CCN2/CTGF mRNA and protein in the tibial epiphysis at 7 days of age; cross-section of the cartilage canal. Serial sections were stained with hematoxylin and eosin (
The histological features and localization of CCN2/CTGF mRNA and protein in the tibial epiphysis at 14 days of age. Serial sections were stained with hematoxylin and eosin (
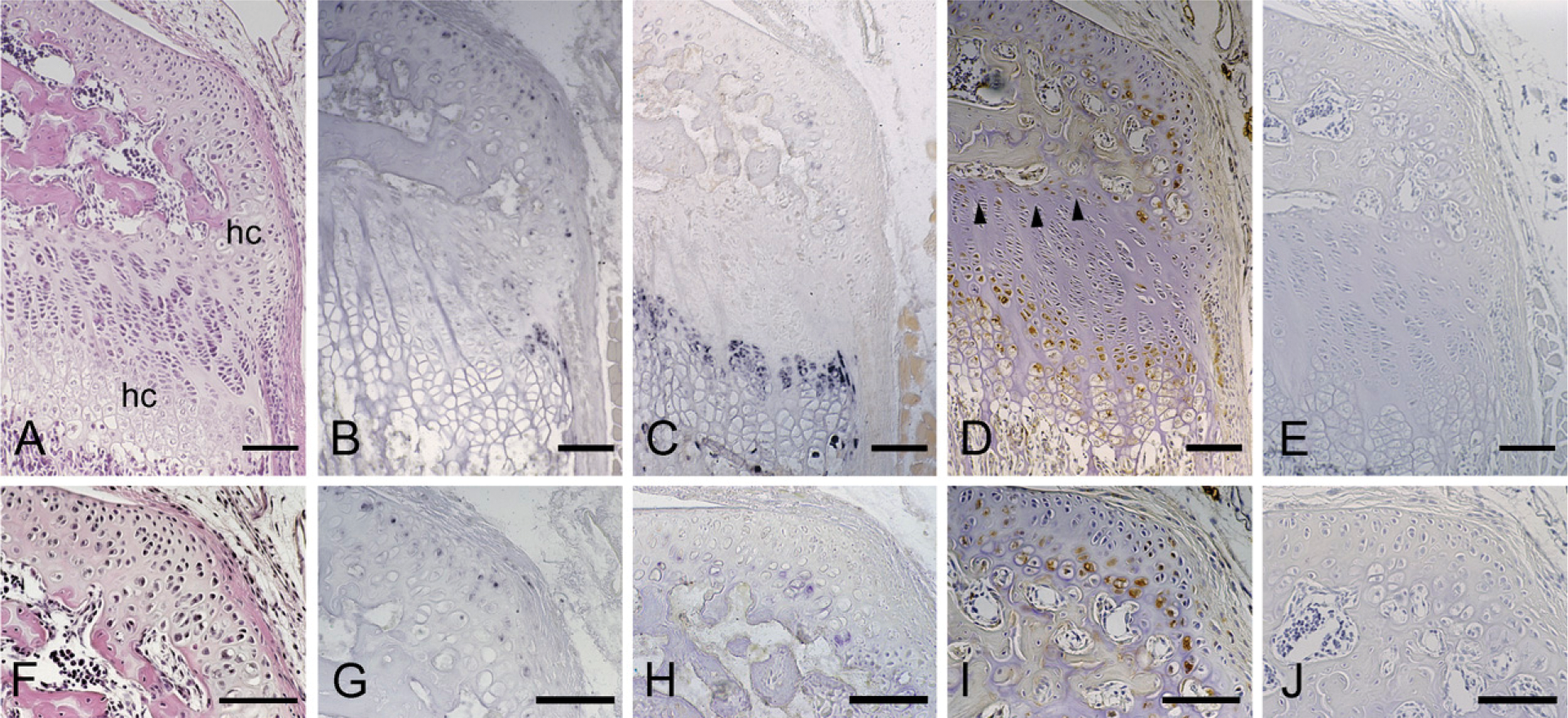
The histological features and localization of CCN2/CTGF mRNA and protein in the tibial epiphysis at 28 days of age. Serial sections were stained with hematoxylin and eosin (
Expression of ccn2/ctgf and Distribution of CCN2/CTGF During Development of the Secondary Ossification Center
After the formation of the cartilage canal, a strong
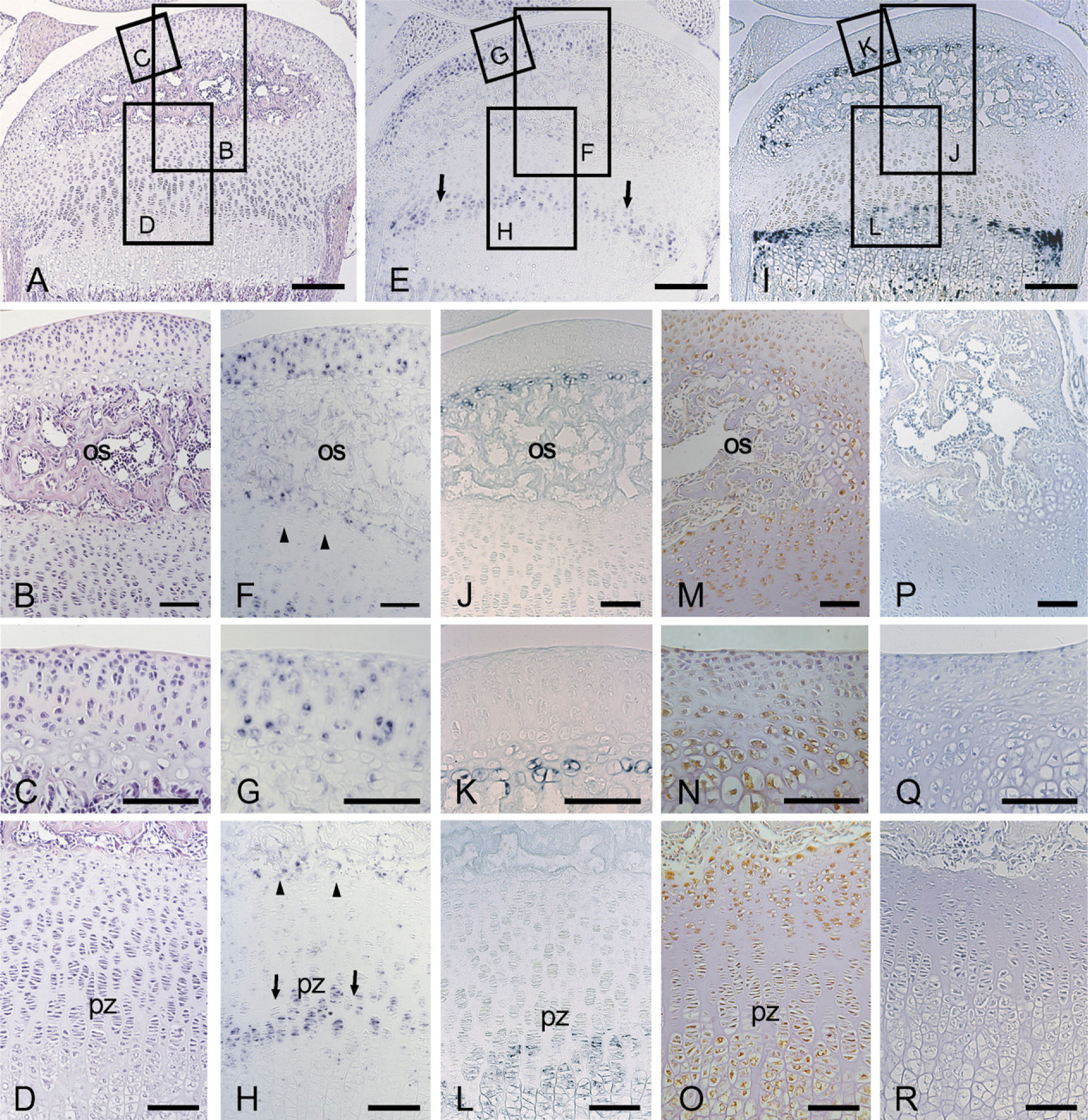
Production of MMP9 in the Cartilage Canal and Secondary Ossification Center in Early Stages of Secondary Ossification Center Formation
Because MMP9 is known to be critically involved in endochondral ossification (Vu et al. 1998), the distribution of MMP9 molecules during the formation of the ossification center was also examined. Positive immunostaining of MMP9 was clearly observed during the formation of the cartilage canal (Figure 6A) and an enlargement of the marrow space (Figure 6D). The MMP9 protein was strongly detected at the blind end of the canal and the wall of the forming marrow space. Furthermore, the MMP9 protein was also detected in the hypertrophic chondrocytes adjacent to the cartilage canal, where CCN2/CTGF protein was detected as well.
Discussion
This study clarified that CCN2/CTGF exhibits highly dynamic patterns of expression that are temporally and spatially restricted during postnatal formation of the secondary ossification center in mouse long bones. These expression patterns suggest that the factor is engaged in establishing epiphyseal vascularization and remodeling of the secondary ossification center. These experiments showed that the CCN2/CTGF protein was present at three sites: the surface of the epiphysis, proliferating and hypertrophic layers in the growth plate and secondary ossification center, and articular cartilage. At these sites, relatively immature chondrocytes expressing
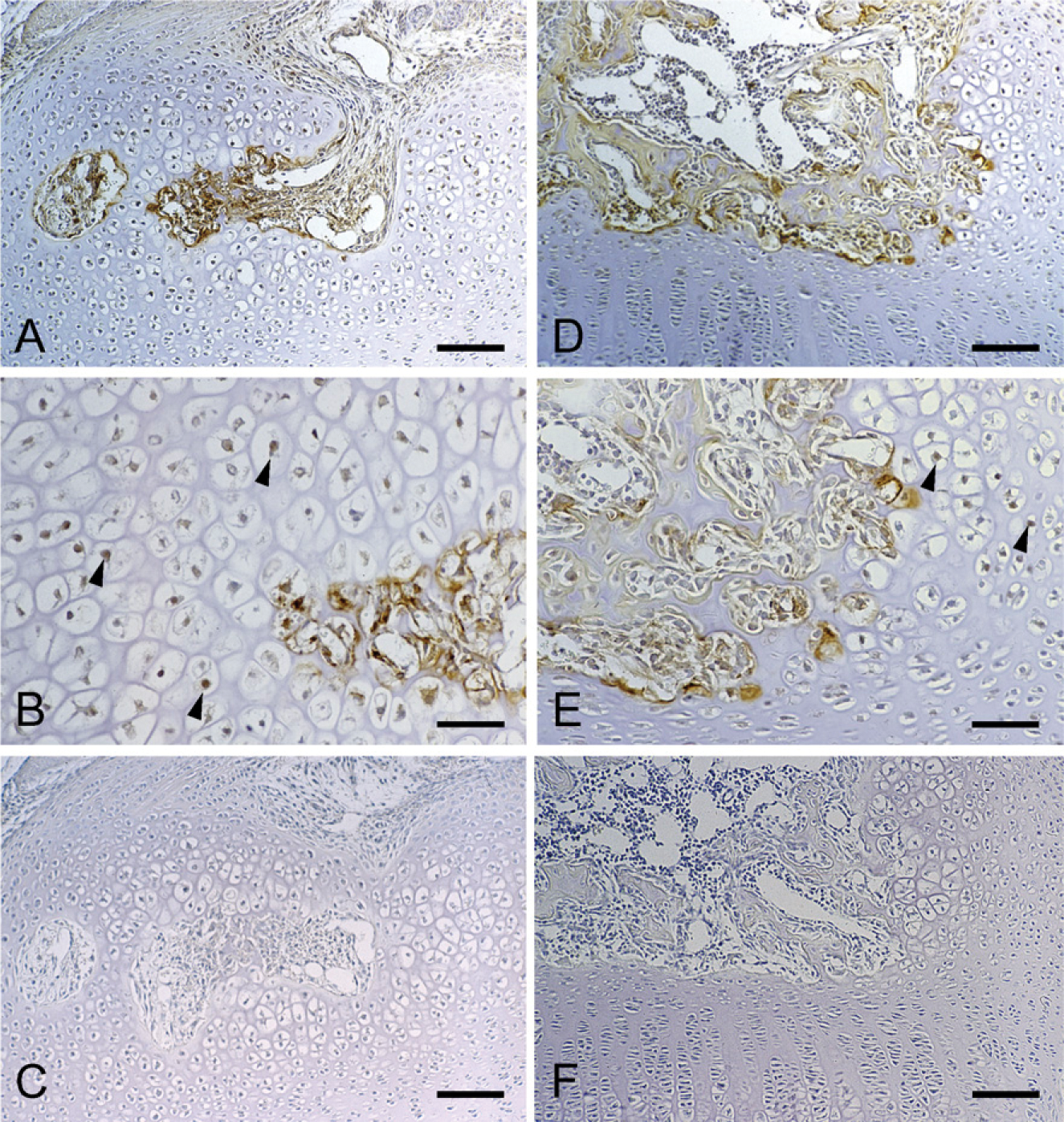
Immunohistochemical staining of MMP9 in the tibial epiphysis at 7 days of age (
The expression pattern of
There is significant evidence indicating that CCN2/CTGF plays a role in the regulation of the endothelial cell function and angiogenesis. CCN2/CTGF regulates the production and/or activity or other angiogenic molecules, such as vascular endothelial growth factor (Inoki et al. 2002; Kondo et al. 2006), or possibly, basic fibroblast growth factor (Brigstock 2002). Moreover, CCN2/CTGF is intrinsically active in in vivo assays for angiogenic activity (Babic et al. 1999; Shimo et al. 1999). In fact, CCN2/CTGF knockout mice exhibited vascular defects during endochondral bone formation (Ivkovic et al. 2003).
It should be noted that CCN2/CTGF may also affect ECM stability or integrity. CCN2/CTGF increases the expression of an MMP in vascular endothelial cells (Kondo et al. 2002). The MMPs are extracellular endopeptidases that regulate cell growth, migration, and ECM remodeling. In addition, the MMPs also have been implicated in a number of physiological and pathological processes including normal and tumor angiogenesis (Bergers et al. 2000; Engsig et al. 2000; Fang et al. 2000). These data together suggest that CCN2/CTGF can drive the balance of proteinases and their respective endogenous inhibitors toward increased proteolysis of the ECM, which eventually promotes vascular endothelial cell migration. This study showed the overlapping distribution of MMP9 and CCN2/CTGF during the formation of the ossification center, which suggested that CCN2/CTGF was produced in the perichondrium and cartilage around the canal and reached their target by diffusion to provoke MMP9 production. In addition, Davoli et al. (2001) previously reported the histozymographic reactions and immunolocalization of active MMP9 during the development of microcirculation of the secondary ossification center in the rat humeral head. Three immunoreactive sites were identified: the walls on the blind end of cartilage canal, segments of the wall around forming marrow space, and along the edges of the wall containing groups of hypertrophic chondrocytes. In this study, MMP9 was observed not only these sites but also in hypertrophic chondrocytes around cartilage canal and marrow space in the central portion of the epiphysis. This broader distribution of reactiveness might be because of the characteristic of the primary antibody that recognizes both the pro-form and active form of mouse MMP9 (the 95-to 105-kDa gelatinase). Supported by the in vivo and in vitro findings above, it is reasonable to hypothesize that MMPs, including MMP9, are playing major roles as cartilage excavators during the formation of the ossification center.
After serial changes during the development of the ossification center,
In conclusion, CCN2/CTGF first accumulated at the center of mouse tibial epiphysis on formation of the cartilage canal. Thereafter, it was distributed at the locations toward the secondary ossification center development. There was also an overlapping distribution of MMP-9 and CCN2/CTGF in the secondary ossification center. These findings suggest that CCN2/CTGF is an important factor for establishing epiphyseal vascularization in determining the secondary ossification center formation, together with MMP9.
Footnotes
Acknowledgments
This work was supported in part by the Grant-in-Aid for Young Scientists (B) (to MO), The Ministry of Education, Culture, Sports, Science and Technology, Japan, and the Grant-in-Aid for Scientific Research (S) (to MT) and (C) (to SK) from the Japan Society for the Promotion of Sciences.
The authors thank Drs. Takashi Nishida and Kumiko Nawachi and Harumi Kawaki for valuable comments and suggestions.