
Abstract
Biosynthesis of steroid hormones in the cortex of the adrenal gland takes place in smooth endoplasmic reticulum and mitochondria and requires NADPH. Four enzymes produce NADPH: glucose-6-phosphate dehydrogenase (G6PD), the key regulatory enzyme of the pentose phosphate pathway, phosphogluconate dehydrogenase (PGD), the third enzyme of that pathway, malate dehydrogenase (MDH), and isocitrate dehydrogenase (ICDH). However, the contribution of each enzyme to NADPH production in the cortex of adrenal gland has not been established. Therefore, activity of G6PD, PGD, MDH, and ICDH was localized and quantified in rat adrenocortical tissue using metabolic mapping, image analysis, and electron microscopy. The four enzymes have similar localization patterns in adrenal gland with highest activities in the zona fasciculata of the cortex. G6PD activity was strongest, PGD, MDH, and ICDH activity was ∼60%, 15%, and 7% of G6PD activity, respectively. The Km value of G6PD for glucose-6-phosphate was two times higher than the Km value of PGD for phosphogluconate. As a consequence, virtual flux rates through G6PD and PGD are largely similar. It is concluded that G6PD and PGD provide the major part of NADPH in adrenocortical cells. Their activity is localized in the cytoplasm associated with free ribosomes and membranes of the smooth endoplasmic reticulum, indicating that NADPH-demanding processes related to biosynthesis of steroid hormones take place at these sites. Complete inhibition of G6PD by androsterones suggests that there is feedback regulation of steroid hormone biosynthesis via G6PD.
G
High G6PD activity has been found in adrenocortical cells of the adrenal gland (Rudolph and Klein 1964; Okano et al. 1965). High activity is related to NADPH production for the biosynthesis of steroids. This is also based on the localization of G6PD activity in the hyaloplasm associated with membranes of smooth endoplasmic reticulum (SER) where a number of steps of the steroid hormone biosynthesis take place in these cells (Bara 1979; Berchtold 1979; Ishibashi et al. 1999). However, other enzymes involved in the biosynthesis of steroid hormones are localized in mitochondria (Bara 1979; Berchtold 1979; Ishimura and Fujita 1997).
Adrenocorticotropic hormone (ACTH) has been shown to stimulate G6PD activity in rat adrenocortical cells (McKerns 1964; Criss and McKerns 1968a), whereas steroids inhibit G6PD activity in the adrenal cortex (Criss and McKerns 1968b).
So far, the contribution to NADPH production by phosphogluconate dehydrogenase (PGD), the third enzyme of the pentose phosphate pathway, and the two other NADPH-producing enzymes, malate dehydrogenase (MDH) and isocitrate dehydrogenase (ICDH), in adrenocortical cells is not known. Therefore, in the present study we investigated the kinetic properties of G6PD, PGD, MDH, and ICDH in rat adrenal gland with in situ techniques and image analysis. Effects of androsterone, an inhibitor of G6PD activity in a number of cell types (Shantz et al. 1989; Gordon et al. 1995), on G6PD activity in adrenal gland were studied as well. Moreover, G6PD and PGD activity was localized ultrastructurally using the ferricyanide method according to Frederiks and Vreeling-Sindelárová (2001).
Materials and Methods
Tissue Preparation
Adrenal glands were obtained from five male Wistar rats weighing 325-350 g after sacrifice by an overdose of phenobarbital or an incision in the vena cava. Principles of laboratory animal care were followed and, according to Dutch law, the Animal Welfare Committee of the Academic Medical Center approved the study. The adrenals were immediately embedded in 7% (w/v) gelatin in distilled water, frozen in liquid nitrogen, and stored at −80C up to maximally 1 week until used. Eight-μm-thick cryostat sections were cut at −25C on a motor-driven Bright cryostat (Huntington, UK) fitted with a rotary retracting microtome, picked up on glass slides, and stored at −25C until used (Van Noorden and Frederiks 1992).
Cell Isolation Procedure
To obtain a suspension of adrenal gland cells, the capsule of the adrenal gland was mechanically removed. Adrenal glands were then dissected into small pieces using scissors. Tissue fragments were incubated in a solution of 2 mg collagenase (Sigma; St Louis, MO) in 5 ml 100 mM phosphate buffer, pH 7.4, for 60 min at 37C. Cells were spun down by centrifugation for 5 min at 1000 × g at 4C, washed three times, and taken up in 0.5 ml 100 mM phosphate buffer, pH 7.4.
Demonstration of G6PD, PGD, MDH, and ICDH Activity
Cryostat sections of adrenal gland were allowed to dry at room temperature for 5 min and were then incubated for the demonstration of G6PD, PGD, MDH, and ICDH activity according to Van Noorden and Frederiks (1992). Incubation medium contained 18% polyvinyl alcohol (PVA, average molecular mass 70,000-100,000; Sigma) in 0.1 M phosphate buffer, pH 7.4, 0.32 mM 1-methoxyphenazine methosulfate (Serva; Heidelberg, Germany), 5 mM sodium azide, 5 mM MgCl2, 5 mM nitro BT (Sigma), and the respective substrate. For G6PD, PGD, MDH, and ICDH, the following substrate concentrations were used: 10 mM glucose-6-phosphate (G6P; Boehringer, Mannheim, Germany), 10 mM 6-phosphogluconate (PG; Boehringer), 100 mM L-malate (Serva), and 20 mM DL-isocitrate (Sigma), respectively. The media were freshly prepared just before incubation and nitro BT was added after being dissolved in a heated mixture of dimethylformamide and ethanol (final dilution of each solvent in the medium was 2% v/v). For the demonstration of G6PD, PGD, MDH, and ICDH activity, sections were incubated for 5 min, 5 min, 30 min, and 30 min at room temperature, respectively. Sections were then rinsed with hot phosphate buffer (0.1 M, pH 5.3, 65C) to remove the incubation medium and to immediately stop the reaction. It is essential to rinse in phosphate buffer, pH 5.3, at 65C to stop the reaction because G6PD shows activity at temperatures up to 60C, whereas it is scarcely present at pH 5.3 (Corpas et al. 1995). Afterwards, sections were embedded in glycerol jelly. Control reactions were performed in the absence of substrate and NADP+ for G6PD, MDH, and ICDH and in the absence of PG for PGD (Butcher and Van Noorden 1985). To test whether PGD activity was involved in the assay for the demonstration of G6PD activity, sections were incubated in the presence of both 10 mM G6P and 10 mM PG. The test minus control reaction of the latter assay was diminished with the test minus control reaction of PGD and was taken as a measure for the actual G6PD activity (Lawrence et al. 1988), showing that PGD activity was not involved in the assay of G6PD activity.
To investigate the possible involvement of hexose-6-phosphate dehydrogenase (H6PD) in the assay of G6PD activity, 10 mM galactose-6-phosphate (Boehringer) was used as a substrate instead of G6P (Kidder 1983).
Effects of different inhibitors on G6PD activity were tested by incubating in the presence of 10 mM epiandrosterone (Sigma), dehydroepiandrosterone (Sigma), and dehydroepiandrosterone sulfate (Sigma), all dissolved in dimethylformamide and 10 mM
Ultrastructural Localization of G6PD and PGD Activity in Isolated Cells
Cells isolated from adrenal gland were incubated for the ultrastructural localization of G6PD and PGD activity as previously described for liver parenchymal cells (Frederiks and Vreeling-Sindelárová 2001). It has been previously shown that unfixed cryostat sections can be used to demonstrate the activity of various types of enzymes (Schellens et al. 2003) but not that of oxidoreductases, including dehydrogenases. However, the activity of G6PD (Frederiks and Vreeling-Sindelárová 2001), xanthine oxidoreductase (Frederiks and Vreeling-Sindelárová 2002), and lactate dehydrogenase (Schellens et al. 2003) was localized ultrastructurally in isolated cells. Therefore, we decided to use isolated cells instead of cryostat sections of adrenal gland to demonstrate G6PD and PGD activity at the ultrastructural level. To protect G6PD and PGD activity in isolated cells against the permeabilization procedure, pelleted cells were incubated in 0.5 ml of a solution of 20 mM NADP+ (Boehringer) in 100 mM phosphate buffer, pH 7.4, for 10 min at 4C. After centrifugation at 1000 × g, 4C, for 5 min, cells were permeabilized by adding 5 ml of a solution containing 0.025% (v/v) glutaraldehyde (Merck) in 100 mM phosphate buffer, pH 7.4. Permeabilization was performed for 30 min at room temperature under continuous rotation of the tubes. The procedure was stopped by washing the cells three times for 1 min with 3 ml 100 mM phosphate buffer, pH 7.4, at 4C, followed by rapid centrifugation at 1000 × g. Cells were resuspended in 0.5 ml of the same buffer. Cell suspensions (0.1 ml) were added to the histochemical media (1 ml) for demonstration of G6PD or PGD activity. Incubation was carried out for 15 min at room temperature under continuous rotation. Incubation medium contained 6% PVA in 100 mM phosphate buffer, pH 7.4, 10 mM G6P or PG, 0.8 mM NADP+, 0.32 mM 1-methoxyphenazine methosulfate, 5 mM MgCl2, 5 mM sodium azide, 10 mM potassium ferricyanide, 30 mM sodium citrate, and 30 mM cupric sulfate. Control reactions were performed by omitting the substrate and NADP+ from the incubation medium. After incubation, the reaction was stopped by adding 5 ml 100 mM phosphate buffer, pH 7.4, at 4C. Cells were washed three times with the same buffer, spun down rapidly at 1000 × g, and finally resuspended in 1 ml buffer. For light microscopic inspection, 10 μl of the suspended cells was placed on glass slides. The suspension was air dried for 30 min at 37C and mounted in glycerol jelly. For electron microscopy, incubated cells were mixed with fixative containing 1% (w/v) glutaraldehyde and 4% (w/v) paraformaldehyde in 100 mM cacodylate buffer, pH 7.4, and fixation lasted for 60 min at 4C. Fixation was followed by centrifugation at 1000 × g at 4C. Cells were rinsed in 100 mM cacodylate buffer, pH 7.4, for 30 min, postfixed in 1% OsO4 (Drijfhout; Amsterdam, The Netherlands) in 100 mM cacodylate buffer, pH 7.4, for 60 min at 4C or with 1% OsO4 and 1.5% potassium ferrocyanide in 100 mM phosphate buffer for 2 hr at 4C and thoroughly rinsed with bidistilled water. Afterwards, samples were dehydrated and embedded in epoxy resin LX-112 (Ladd; Burlington, VT) according to standard procedures. Ultrathin sections (50- to 70-nm thick) were cut on an LKB Ultratome III ultramicrotome and studied with an EM-10c transmission electron microscope (Zeiss; Oberkochen, Germany).
Image Analysis
Formazan in adrenocortical cells of rat adrenal gland was measured by image analysis according to Chieco et al. (1994) using a Vanox-T photomicroscope (Olympus; Tokyo, Japan) with a ×2 objective (numerical aperture 0.08). Experiments were performed in triplicate, and two preparations were incubated per experiment. Sections were illuminated with white light from a stabilized power supply after filtering by infrared blocking filters (Jonker et al. 1997) and a monochromatic filter of the isobestic wavelength of nitro BT formazan (585 nm) (Van Noorden and Frederiks 1992). Images of the sections were captured using a CCD camera that was attached to a frame grabber (Scion image 1.59 for Mac; Scion, Frederick, MD) and a computer (8100; Apple Macintosh, Cupertino CA). Gray values were converted to absorbance values by using a set of neutral density filters (Jonker et al. 1997). Absorbance values were converted into amounts (micromoles) of substrate converted per minute per area (Van Noorden and Frederiks 1992). Absorbance values of control reactions were subtracted from test values to obtain specific activity (Butcher and Van Noorden 1985). Measurements were performed in the zona fasciculata of the cortex.
Statistics
Statistical processing of data was performed using Excel 97 (Microsoft; San Jose, CA) and SPSS 8.0 for Windows (SPSS; Chicago IL). Vmax and Km values were expressed as mean values ± SD. A paired Student's
Flux rates (Φ) of G6PD and PGD with their respective substrates were calculated using the formula:
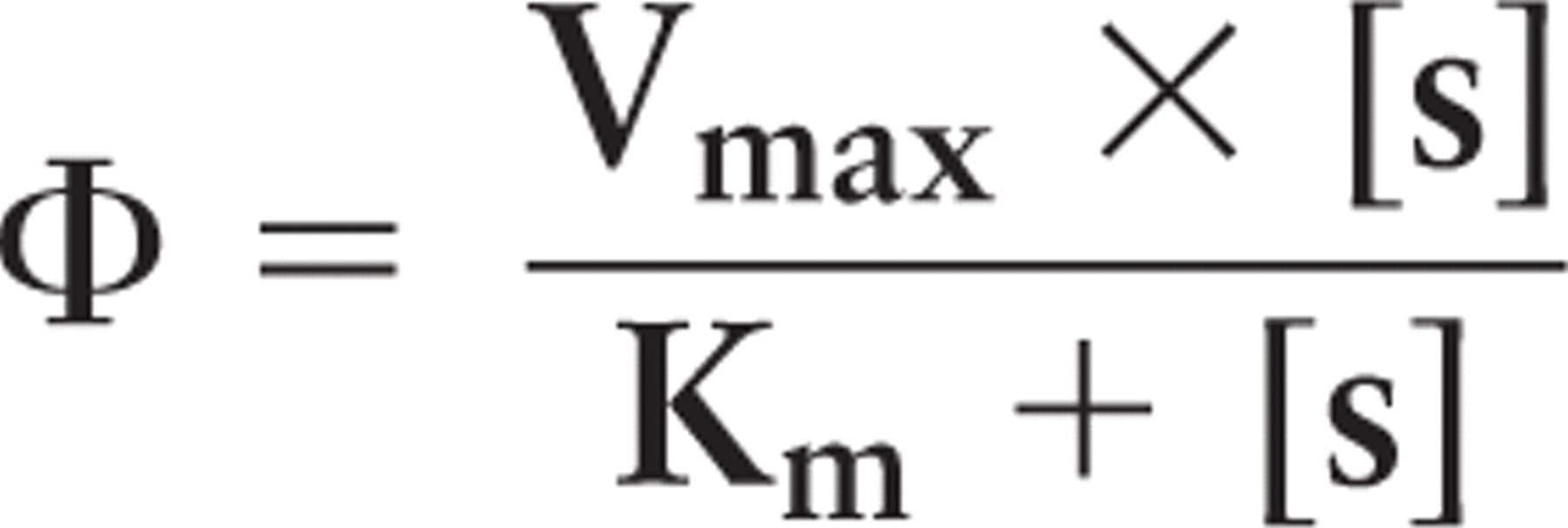
with an assumed physiological substrate concentration of 0.1 mM (Teutsch 1981).
Results
Localization of NADPH-producing Enzymes
G6PD, PGD, MDH, and ICDH activity in serial sections of adrenal gland showed similar distribution patterns. Enzyme activities were highest in cells of the zona fasciculata of the cortex of adrenal gland. A somewhat lower activity was found in cells of the zona glomerulosa and zona reticularis, whereas activities were scarcely present in cells of the medulla and the capsule of the gland (Figure 1). Only small amounts of final reaction product were formed in cortical cells of the gland after incubation in the absence of substrate and coenzyme.
To exclude the involvement of H6PD in the detection of G6PD activity, the incubation was performed in the presence of galactose-6-P instead of G6P. The amounts of final reaction product formed in the presence of galactose-6-P were similar to the amounts produced in the absence of substrate (data not shown). This suggests that H6PD was not interfering in the G6PD reaction.
Quantification of G6PD, PGD, MDH, and ICDH Activity
Epiandrosterone and dehydroepiandrosterone inhibited G6PD activity almost completely in the adrenal cortex (90% and 95%, respectively). Dehydroepiandrosterone sulfate inhibited G6PD activity by only 30%, possibly due to the poor solubility of this inhibitor in aqueous media.
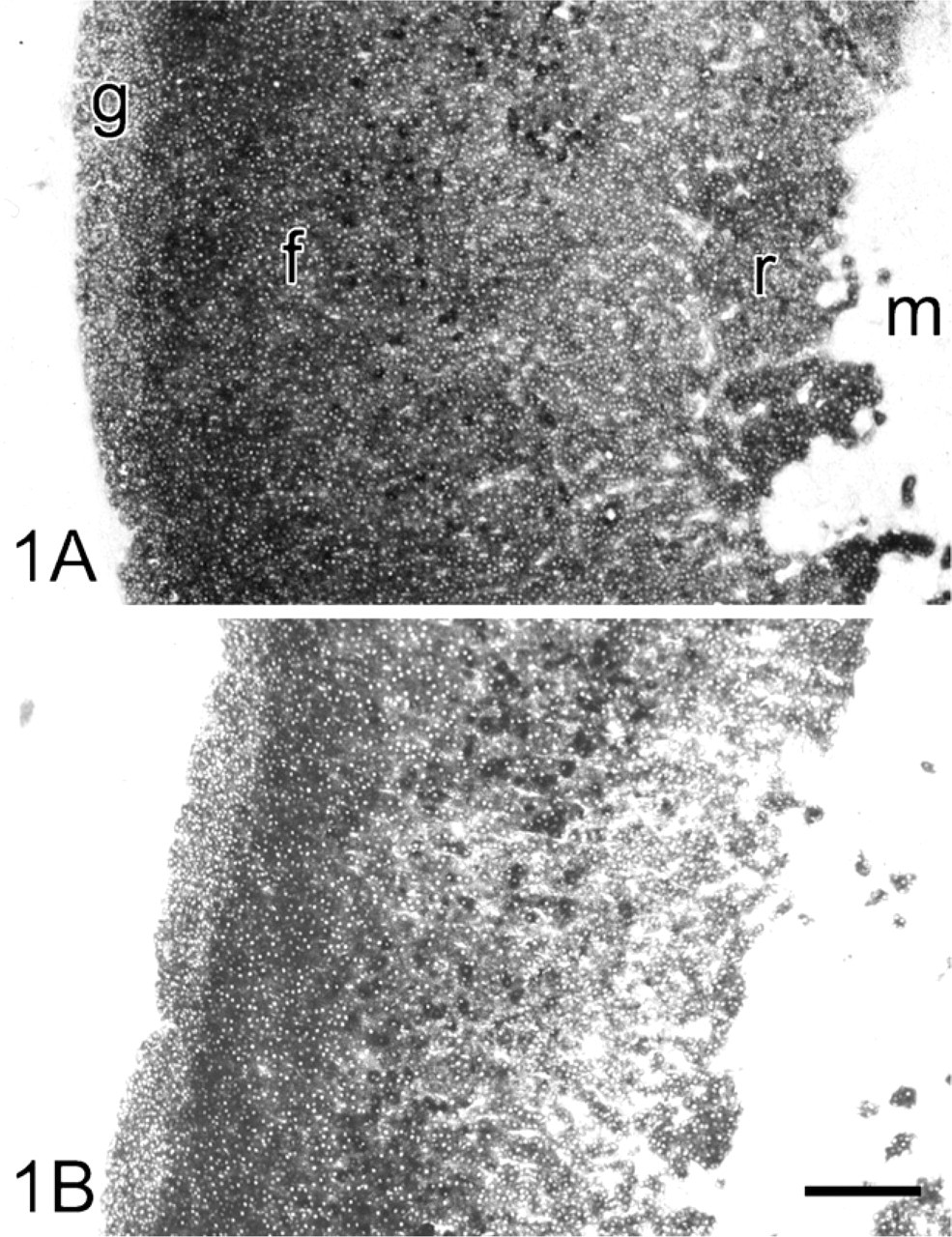
Light microscopy of cryostat sections of rat adrenal gland incubated for the demonstration of glucose-6-phosphate dehydrogenase (G6PD) activity (
Table 1 demonstrates that G6PD showed the highest activity in the zona fasciculata of adrenal glands. PGD was approximately half the G6PD activity, whereas the relative contribution of MDH and ICDH to NADPH synthesis was only 15% and 7%, respectively. Determination of Vmax and Km values of G6PD and PGD in the zona fasciculata showed that G6PD activity was ∼75% higher than PGD activity, whereas the Km value of G6PD for G6P was approximately two times higher than the Km value of PGD for PG (Table 2). As a consequence, imaginary fluxes calculated at an assumed physiological substrate concentration of 0.1 mM (Teutsch 1981) were largely similar through both enzymes. Metabolic fluxes are considered to be direct parameters of metabolic activity (Newsholme et al. 1980).
Activities of G6PD, PGD, MDH, and ICDH in the zona fasciculata of rat adrenal gland as determined quantitatively with the use of image analysis in cryostat sections

Values were determined in three glands and are expressed as percentage (mean ± SD) related to G6PD activity. Differences among all values were statistically significantly. G6PD, glucose-6-phosphate dehydrogenase; PDG, phosphogluconate dehydrogenase; MDH, malate dehydrogenase; ICDH, isocitrate dehydrogenase.
Ultrastructural Localization of G6PD and PGD Activity
Suspensions of cells from adrenal glands were incubated for the demonstration of G6PD and PGD activity with the ferricyanide method. At the electron microscopic level, cells were observed with lipid droplets, extensive networks of SER, and spherical mitochondria with tubular cristae typical for adrenocortical cells. These cells contained varying amounts of electron-dense precipitate, likely dependent on their localization in the cortex of the adrenal glands. Cells with the highest amounts of electron-dense precipitate were carefully studied. Electron-dense reaction product due to G6PD activity was present in the cytoplasm associated with membranes of SER and free ribosomes (Figure 2). Nuclei always contained some precipitate, but this was considered to be nonspecific because it was also observed after control incubations. Final reaction product was not observed after incubation in the absence of G6P and NADP+, except for nuclei. A similar distribution pattern of electron-dense reaction product was observed when cells were incubated for the demonstration of PGD activity (Figure 2D).
Discussion
The present study shows that four NADPH-producing enzymes (G6PD, PGD, MDH, and ICDH) have similar localization patterns in adrenal gland. Highest activities were found in the zona fasciculata in the cortex where glucocorticoids are synthesized, whereas medulla showed scarcely any activity of these enzymes (Figure 1). The two NADPH-producing enzymes of the phosphate pathway, G6PD and PGD, produce ∼85% of the total pool of NADPH. Both enzymes can produce roughly the same amount of NADPH at physiological substrate concentrations (Table 2). Therefore, it can be concluded that the pentose phosphate pathway is the major provider of NADPH for glucocorticoid synthesis in the zona fasciculata.
Vmax and Km values and imaginary fluxes (Φ) of G6PD and PGD in the zona fasciculata of rat adrenal gland as determined quantitatively with the use of image analysis in cryostat sections
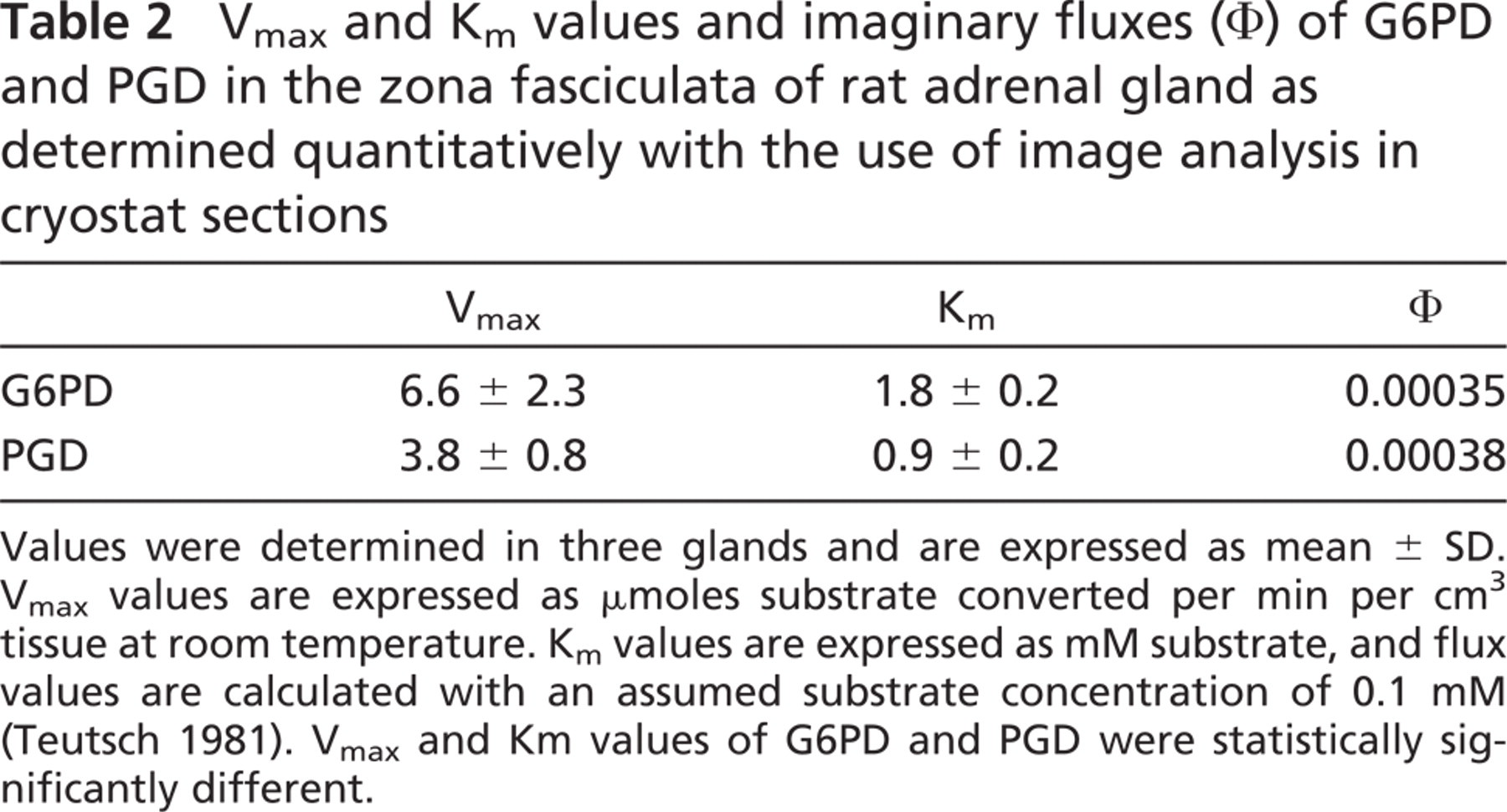
Values were determined in three glands and are expressed as mean ± SD. Vmax values are expressed as μmoles substrate converted per min per cm3 tissue at room temperature. Km values are expressed as mM substrate, and flux values are calculated with an assumed substrate concentration of 0.1 mM (Teutsch 1981). Vmax and Km values of G6PD and PGD were statistically significantly different.
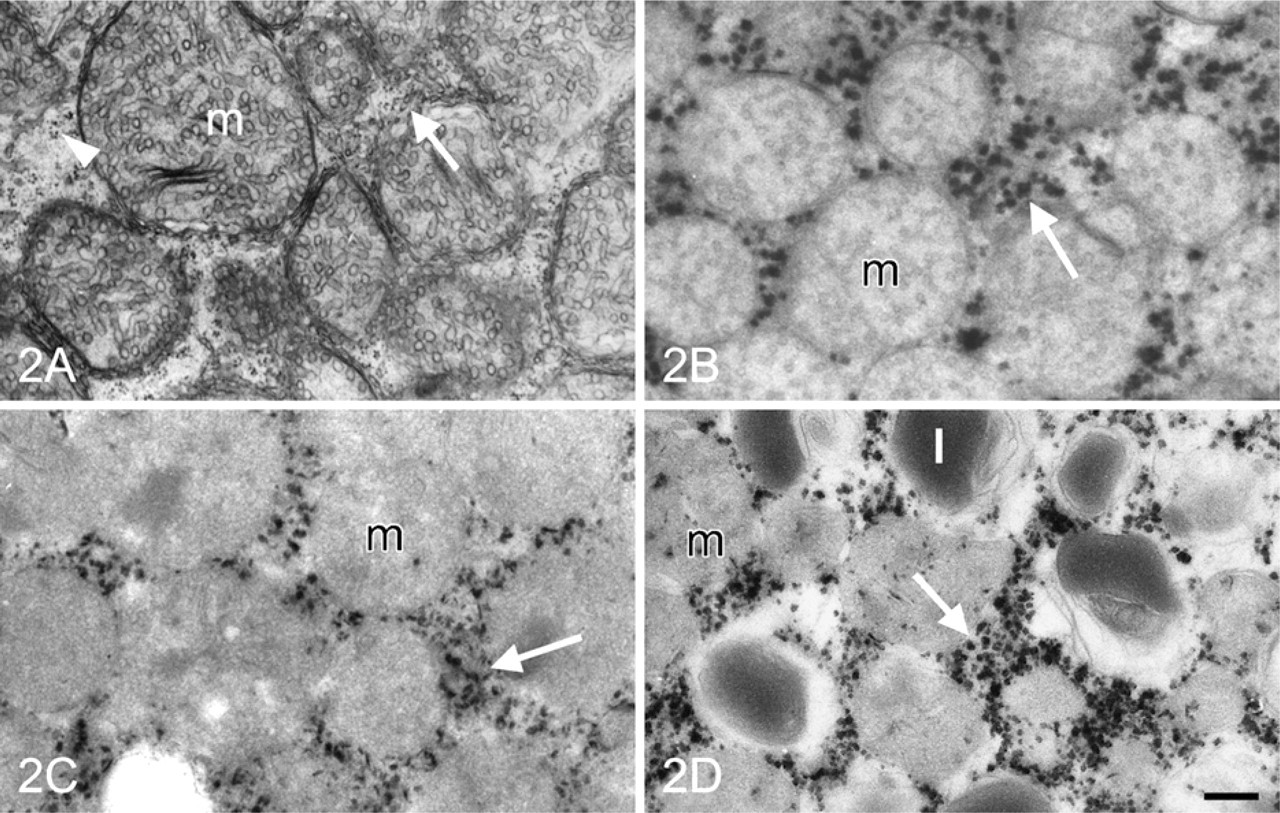
Electron micrographs of isolated cells of adrenal gland. (
Steroids produced in the zona fasciculata are mainly glucocorticoids, whereas zona glomerulosa and zona reticularis synthesize and secrete mineralocorticoids and androgens, respectively. In the present study we have shown that DHEA is a strong inhibitor of G6PD activity, whereas ACTH has been demonstrated to stimulate G6PD activity in the cortex (McKerns 1964; Criss and McKerns 1968a). This suggests that the demand for NADPH and indirectly for hormones produced by the cortex of the adrenal gland may be regulated by ACTH and DHEA via G6PD activity.
In the present study we also demonstrated that G6PD activity was localized in the cytoplasm, associated with SER and free ribosomes (Figure 2). This is in accordance with previous findings in adrenal gland (Berchtold 1979; Ishibashi et al. 1999), testis (Bara 1979), and intestine (Ninfali et al. 2001). As far as we know, this is the first study on the ultrastructural localization of PGD activity. The finding that G6PD and PGD activity have a similar distribution pattern in adrenocortical cells is not surprising. NADPH produced by G6PD and PGD at SER is used for the synthesis of sterols as a precursor for steroid hormones from acetyl CoA. Many enzymes involved in the biosynthesis of steroid hormones are localized at membranes of SER (Berchtold 1977; Bara 1979; Balasubramaniam et al. 1981; Ishimura and Fujita 1997), although certain enzymes are found at inner membranes of mitochondria (Berchtold 1977; Ishimura and Fujita 1997). These findings suggest that NADPH-demanding processes related with biosynthesis of steroid hormones occur at membranes of SER, whereas NADPH produced at free ribosomes is used for other purposes.
In conclusion, both G6PD and PGD, but not MDH and ICDH, have a high capacity to produce NADPH in SER for steroid biosynthesis in adrenocortical cells. This pathway is regulated by androsterones via G6PD.
Footnotes
Acknowledgements
The authors thank Mr. A. Maas and Mr. G. van Woerkom for obtaining adrenal glands, Mr. J. Peeterse and Mr. C. Gravemeijer for photographic work, and Mrs. T.M.S. Pierik for preparation of the manuscript.