
Abstract
Glucose-6-phosphate dehydrogenase (G6PD) deficiency is a X-chromosomally transmitted disorder of the erythrocyte that affects 400 million people worldwide. Diagnosis of heterozygously-deficient women is complicated: as a result of lyonization, these women have a normal and a G6PD-deficient population of erythrocytes. The cytochemical assay is the only reliable assay to discriminate between heterozygously-deficient women and non-deficient women or homozygously-deficient women. G6PD deficiency is mainly found in areas where malaria is or has been endemic. In these areas, malaria is treated with drugs that can cause (severe) hemolysis in G6PD-deficient individuals. A cheap and reliable test is necessary for diagnosing the deficiency to prevent hemolytic disorders when treating malaria. In this review, it is concluded that the use of two different tests for diagnosing men and women is the ideal approach to detect G6PD deficiency. The fluorescent spot test is inexpensive and easy to perform but only reliable for discriminating hemizygous G6PD-deficient men from non-deficient men. For women, the cytochemical assay is recommended. However, this assay is more expensive and difficult to perform and should be simplified into a kit for use in developing countries.
Keywords
G6PD, the key enzyme in the oxidative pentose phosphate pathway, converts nicotinamide adenine dinucleotide phosphate (NADP+) into its reduced form, NADPH. NADPH is essential for protection against oxidative stress in erythrocytes. G6PD deficiency causes increased susceptibility of erythrocytes to H2O2 and other reactive oxygen species that can lead to hemolytic anemia, favism (especially in G6PD Mediterranea), chronic non-spherocytic hemolysis, spontaneous abortions (Toncheva and Tzoneva 1985), and neonatal hyperbilirubinemia resulting in neonatal kernicterus (Beutler 1994). Numerous drugs and chemicals, such as primaquine and dapsone (DeGowin et al. 1966; Clyde 1981; Wolf et al. 2002; Mason et al. 2007), in-gestion of fava beans, and stress (for example, infection) can induce hemolytic anemia in G6PD-deficient individuals (Table 1).
G6PD deficiency is transmitted X-chromosomally, and the deficiency can be detected reliably in homozygous women and hemizygous males with a number of tests. Diagnosing G6PD deficiency in heterozygous women is difficult, and a large part of this group is missed (Reclos et al. 2000; Zaffanello et al. 2004).
G6PD deficiency can cause great problems, especially in malaria treatment. Dapsone, used in combination therapy for the treatment of
Drugs and chemicals that have been reported to cause hemolysis in G6PD-deficient individualsa

Three tests to detect G6PD deficiency are: the fluorescent spot test (Tan and Whitehead 1969); the spectrophotometric assay, or so-called enzyme activity assay (Beutler 1966); and the cytochemical assay (Van Noorden et al. 1982; Van Noorden and Vogels 1985). The fluorescent spot test and the spectrophotometric assay are inexpensive and easy to perform, but only moderately reliable for detection of G6PD deficiency in heterozygous women. On the other hand, the cytochemical assay is reliable for detection of G6PD deficiency in heterozygous women (Table 2; Wolf et al. 1987; Tagarelli et al. 2006), but this is a relatively complicated test that requires a number of steps. In this review, the value of these tests is critically assessed, and recommendations are given with respect to detection of G6PD deficiency in relation to malaria.
Genetics of G6PD
The gene that codes for G6PD is located in a cluster of genes on the distal long arm of the X chromosome (locus q28). Genes for color vision (Filosa et al. 1993), hemophilia A (Boyer and Graham 1965), and the fragile X-syndrome (Oberle et al. 1987) are also located here. The G6PD gene is 18 kb long and contains 13 exons and 12 introns, of which the length varies between 12 bp and 236 bp (Martini et al. 1986). The sequence of the entire gene is known (Chen et al. 1991). Over 450 G6PD variants have been differentiated on the basis of enzyme kinetics, physicochemical characteristics, and other parameters (Luzzatto and Battistuzzi 1985; Chen et al. 1991). Almost 300 variants have been acknowledged by the World Health Organization (World Health Organization 1967).
G6PD deficiency is transmitted X-chromosomally. Females can thus be homozygously deficient or heterozygously deficient, whereas males are hemizygously deficient. Heterozygously-deficient women have a mixed population of erythrocytes, owing to random inactivation of one of the two X chromosomes, known as lyonization. One of the erythrocyte populations is G6PD deficient; the other has normal G6PD function (Figure 1; Lyon 1961; Davidson et al. 1963).
The DNA sequence of 140 known mutations has been established (Beutler and Vulliamy 2002). Most mutations are point mutations and small deletions that cause structural defects in the enzyme. Large deletions or expression defects are not known in the gene or its promoter, except one. One mutation causes a frame shift. The lack of severe mutations indicates that total G6PD deficiency is lethal. In most cases, mutations cause instability of the enzyme or altered activity, usually by decreased affinity of G6PD for its substrates, NADP+ or glucose-6-phosphate (G6P) (Luzzatto 2006). Mutations are transmitted from parent to child, although a few spontaneous mutations have been described (Roos et al. 1999).
Enzymology of G6PD
In inactive form, G6PD is a monomer of 515 amino acids and has a molecular mass of over 59 kDa (Rattazzi 1968). One of its substrates, NADP+, is required for activation of the enzyme. Activation occurs by the formation of a dimer or a tetramer that contains tightly bound NADP+ (Wrigley et al. 1972).
G6PD catalyses the first step and controls the oxidative pentose phosphate pathway (Figure 2). In this pathway, NADP+ is converted into NADPH, and G6P is converted into a pentose sugar, ribulose-5-phosphate, precursor of DNA, RNA, and ATP. Ribulose-5-phosphate can be shuttled into glycolysis via the anaerobic part of the pentose phosphate pathway that is controlled by transketolase (Turner 2000). NADPH is the most important reducing agent in the cytoplasm of cells (Koehler and Van Noorden 2003), including erythrocytes, and is essential for biosynthesis and detoxification reactions.
Sensitivity and specificity of the fluorescent spot test, the spectrophotometric assay, and the cytochemical assay for determination of G6PD activity in hemizygous males, homozygous females, and heterozygous femalesa

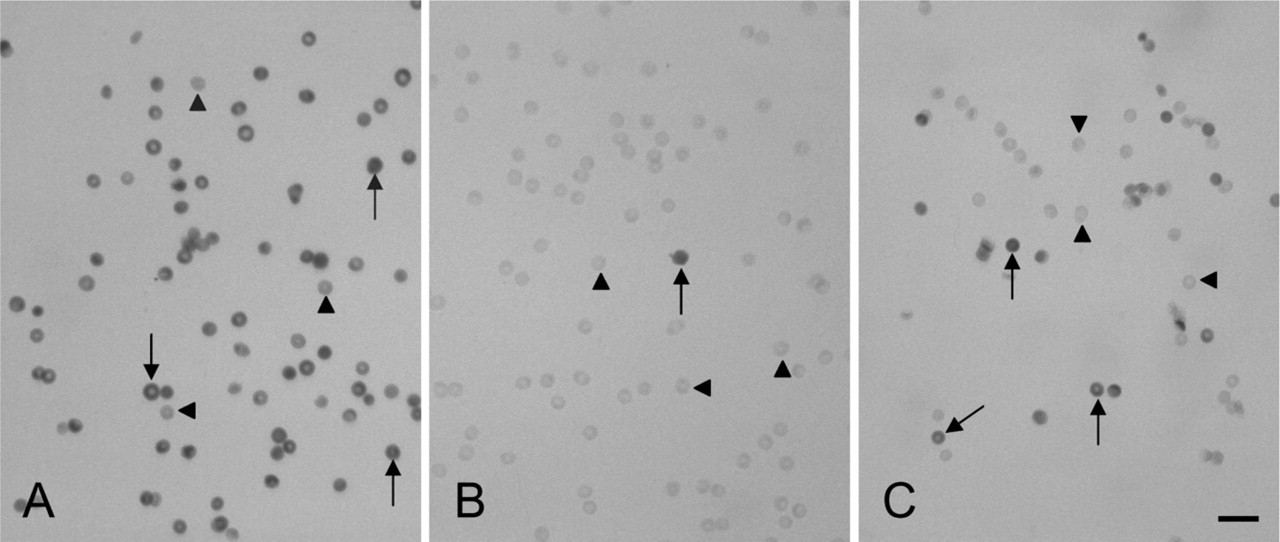
Erythrocytes after cytochemical staining of glucose-6-phosphate dehydrogenase (G6PD) activity showing stained G6PD-containing erythrocytes (arrows) and unstained G6PD-deficient erythrocytes (arrowheads). (
G6PD in the Erythrocyte
During aging of the erythrocyte, the quantity of active G6PD decreases, and older erythrocytes become vulnerable to oxidative stress. Mature erythrocytes do not have a nucleus, mitochondria, or ribosomes, and thus are not able to synthesize new proteins (Marks et al. 1958). When the oxidative pentose phosphate pathway functions poorly due to G6PD deficiency, transcription of the G6PD gene is increased in cells with a nucleus (all types of human cells except erythrocytes and platelets) as a compensation for the deficiency. Erythrocytes are not able to synthesize new proteins and compensate for poor G6PD function. As the deficient erythrocyte ages, the quantity of active G6PD diminishes rapidly, increasing the vulnerability of not only old but also younger erythrocytes to oxidative stress.
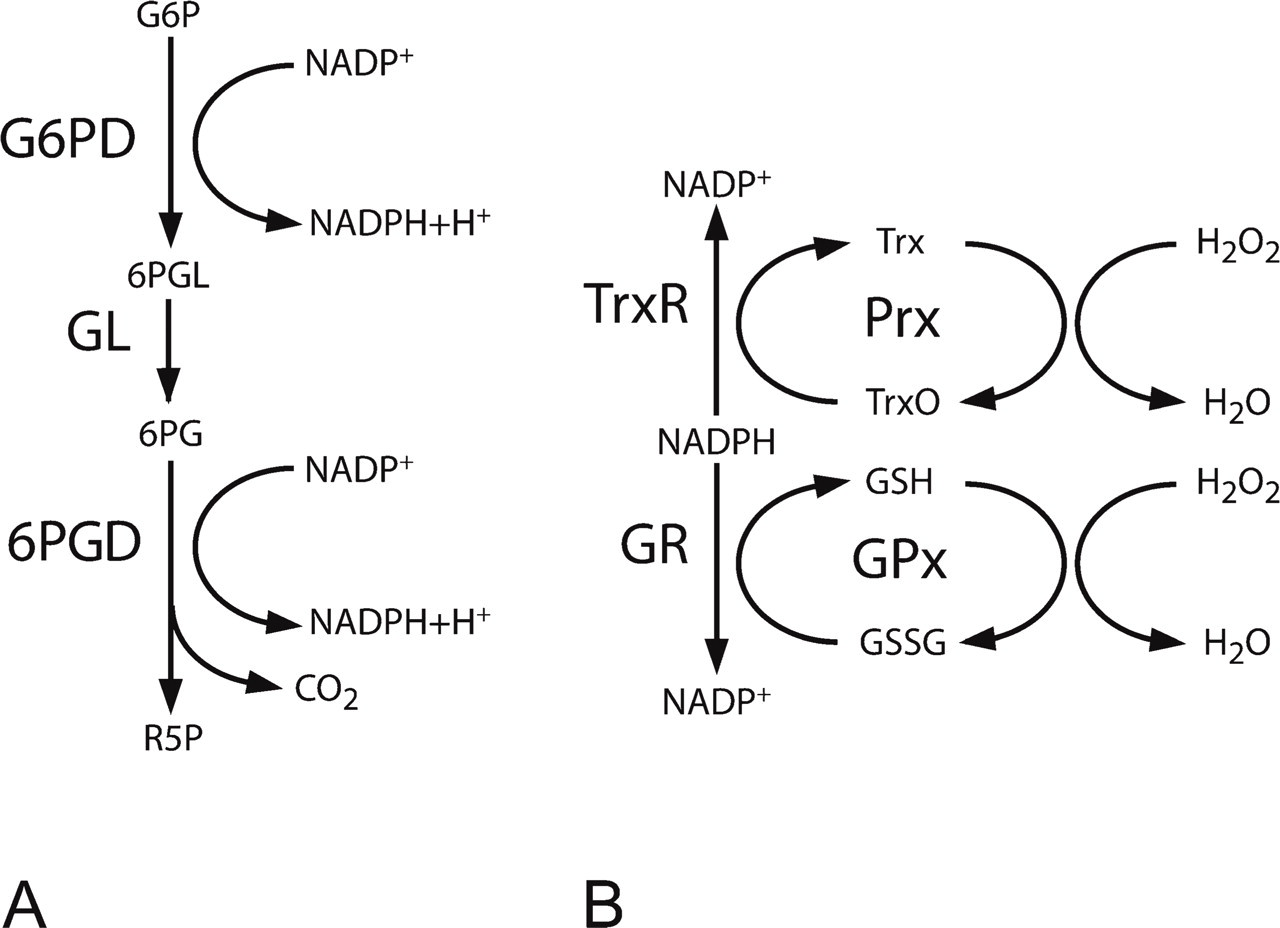
The NADPH-producing steps of the oxidative pentose phosphate pathway (G6PD and 6PGD; Wamelink et al. 2008). (
The severity of G6PD deficiency is usually measured in four classes (World Health Organization 1989; Beutler 1994). These are described as: class I (severe mutations with chronic non-spherocytic hemolytic anemia); class II (intermediate, <10% of normal G6PD function); class III (mild, 10–60% of normal G6PD function); and class IV (asymptomatic, 60–100% of normal G6PD function). Sometimes a fifth class of mutations causing increased G6PD activity is included (Beutler 1994; Frank 2005).
Class I defects are probably caused by mutations in the region of the enzyme where NADP+ or G6P binds. The genetic background of the other classes is unknown. Class I deficiencies are rare and can be severe enough to cause transfusion dependency. Patients in this class usually suffer from chronic non-spherocytic hemolytic anemia. Erythrocytes of these patients have a considerably shorter lifespan then healthy erythrocytes, even when they are not exposed to oxidative stress. G6PD deficiency renders the erythrocytes susceptible to damage from stress encountered in the circulation (Luzzatto 1975; Beutler 1996).
In healthy erythrocytes, only 1–2% of the total enzyme capacity is used, even in episodes of great hemolytic stress (World Health Organization 1989). G6PD-deficient erythrocytes do not have the same spare capacity that can be used in case of hemolytic stress. Nonetheless, patients with class II, III, or IV mutations are usually asymptomatic. Class III deficiencies are the most prevalent defects. Hemolysis due to stress is self-limiting in this class. Only the older erythrocytes lack sufficient active G6PD to counteract oxidative stress episodes (Beutler and Duparc 2007). Class II deficiencies are usually asymptomatic as well, but both young and old erythrocytes are susceptible to oxidative stress. Hemolysis in this class is more severe and not self-limiting. Hemolysis stops after removal of the stress-causing agent (Beutler et al. 1954; Dern et al. 1954; Beutler 1994).
Peroxide Metabolism in the Erythrocyte
It is the lack of NADPH that causes hemolysis of erythrocytes, because H2O2 and other reactive oxygen species cannot be reduced, leading to oxidation of hemoglobin to methemoglobin and membrane damage (Ruwende and Hill 1998).
Glutathione peroxidase is a major reductor of H2O2 in cells (Low et al. 2008). This enzyme uses reduced glutathione as substrate, but glutathione is usually present in an oxidized state. NADPH is required for the reduction of oxidized glutathione by glutathione reductase. End products of this reaction are reduced glutathione and NADP+ (Figure 2). Not only glutathione peroxidase reduces H2O2 in the erythrocyte. Recently, peroxiredoxins have been found to play a major role in the reduction of H2O2 in erythrocytes. Peroxiredoxins reduce H2O2 via the thioredoxin/thioredoxin reductase system (Figure 2; Low et al. 2008). Furthermore, catalase converts H2O2 into O2 and H2O. The reduction of H2O2 by catalase is NADPH independent, but NADPH reduces the susceptibility of catalase to inactivation by its own substrate H2O2 (Kirkman and Gaetani 1984). The exact role of these three systems in peroxide metabolism in erythrocytes has not yet been established.
In the case of G6PD deficiency, erythrocytes cannot produce enough NADPH, which affects glutathione peroxidase, thioredoxin reductase, and catalase activity. When erythrocytes are exposed to oxidative stress, cells cannot eliminate this stress, causing hemolysis.
Malaria and G6PD Deficiency
Acute and chronic malaria can cause anemia. When severe G6PD deficiency complicates malaria infection, treatment with primaquine or dapsone can lead to life-threatening acute intravascular hemolysis followed by anemia and acute renal failure (Wickramasinghe and Abdalla 2000). Primaquine and dapsone are not very frequently used in malaria therapy, but recently, White (2008) proposed a role for primaquine in a combination therapy with artemisinin derivatives for the treatment of
Prevalence of G6PD Deficiency
Worldwide, 300–400 million people carry at least one deficient G6PD gene (World Health Organization 1989; Nkhoma et al. 2009). The frequency of mutations varies greatly among different populations (Nkhoma et al. 2009). G6PD A- is the most prevalent mutation in Africans and Afro-Americans. This class III mutation has a gene frequency of 11%. G6PD B-(Mediterranea) is a more severe class II deficiency. This mutation is commonly found in populations living in or originating from around the Mediterranean Sea. The prevalence of G6PD Mediterranea varies greatly; frequencies of 2–20% are found in Greece, Turkey, and Italy. The highest prevalence of G6PD deficiency (70%) is found in Kurdish Jews (Figure 3; Beutler 1996; Nkhoma et al. 2009).
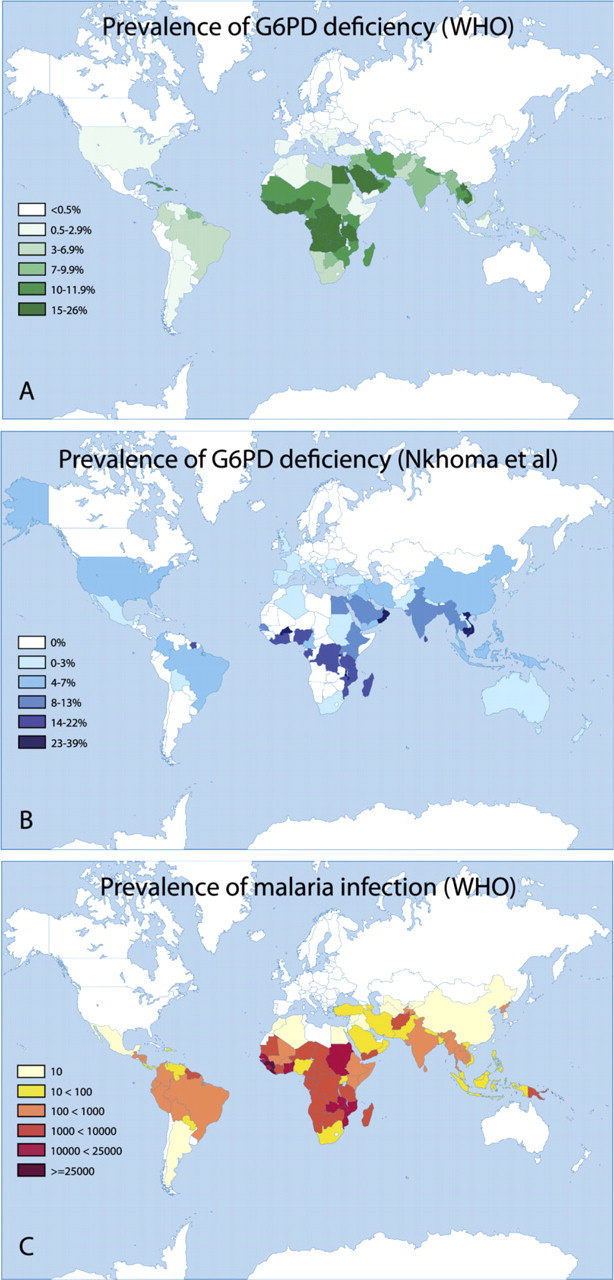
Worldwide prevalence of G6PD deficiency according to the World Health Organization (1989). With permission, World Health Organization. Bull World Health Organ 67: 601–611, 1989. (
{kind=link}
G6PD deficiency is mainly found in Africa, Asia, and Mediterranean Europe, areas where malaria is endemic, or has been endemic (Figure 3; Tishkoff et al. 2001). It has been suggested that the deficiency offers protection against malaria infection (Luzzatto and Bienzle 1979). Hemizygously-deficient males and homozygously-deficient females that have been infected with Plasmodium falciparum are less ill than non-G6PD-deficient individuals. The infection is usually not lethal in G6PD-deficient individuals (Tripathy and Reddy 2007), possibly because Plasmodium can proliferate less efficiently in their erythrocytes (Luzzatto et al. 1969; Roth et al. 1983). It is still uncertain whether this advantage applies only to hemizygous males and homozygous females. According to Guindo et al. (2007), heterozygous females suffer from the same morbidity and mortality of
Detection of G6PD Deficiency
There are several tests that can be used for the detection of G6PD deficiency, but only a few tests diagnose G6PD deficiency in heterozygous women reliably. DNA tests can be used for the diagnosis of G6PD deficiency. In these extremely reliable tests, primers are used to check whether the G6PD gene contains a mutation (Lin et al. 2005; Ko et al. 2006). They can be used for the diagnosis of homozygously, hemizygously, and heterozygously-deficient patients but have the great disadvantage that only one mutation can be analyzed with one primer. At present, 140 mutations are known, and the DNA sequences of these mutations have to be known before primers can be designed. It is unlikely that rare or new mutations will be found with DNA tests. At the moment, DNA tests can only be used for screening of prevalent mutations, for example G6PD A- (Bang-Ce et al. 2004). Moreover, DNA tests are expensive and require sophisticated equipment, which makes them unsuitable for diagnosis on a large scale and impossible to apply in third-world countries.
Tests based on the measurement of the NADPH production capacity of G6PD can be used for the diagnosis of all mutations. The most frequently used tests that measure NADPH production are the fluorescent spot test, the spectrophotometric assay, and the cytochemical assay. The tests are based on the formation of fluorescence (the fluorescent spot test and the spectrophotometric assay) or color (cytochemical assay). NADPH fluoresces when it is excited with light of a wavelength of 340 nm, whereas NADP+ does not, and NADPH converts colorless tetrazolium salt into colored formazan, whereas NADP+ does not. The Brilliant Cresyl Blue (BCB) dye test, based on the reduction of BCB to a colorless state by NADPH, was previously a commonly used test but has largely been replaced by the fluorescent spot test (World Health Organization 1967; Beutler 1994). In some tests, the BCB is replaced by dichlorophenol indophenol (Bernstein 1962). In Asia, the metosulfate-3-(4,5-dimethylthiazal-2-yl)-2,5-diphenyltetrazolim bromide test (PMS/MTT test) is commonly used (Nkhoma et al. 2009), but this test also does not detect heterozygous women reliably (Tantular et al. 1999).
Erythrocytes of a heterozygously G6PD-deficient patient after cytochemical staining. The erythrocytes marked with arrows contain G6PD activity; the erythrocytes marked with arrowheads are G6PD deficient. (
Fluorescent Spot Test
When erythrocytes contain enough functional G6PD, NADP+ is converted into NADPH (Figure 2). After addition of G6P and NADP+, the spot of blood fluoresces when excited at 340 nm (Tan and Whitehead 1969; Beutler 1971). The fluorescent spot test is reliable for the detection of hemizygous males and homozygous females, but is unreliable for the detection of heterozygous females. A sensitivity of 32% and specificity of 99% has been found for the detection of heterozygously-deficient women (Table 2). The population of healthy erythrocytes in heterozygous females (on average 50% of the erythrocytes) converts enough NADP+ into NADPH to produce fluorescent spots, and then the heterozygous deficiency is missed. The fluorescent spot test requires a black light (a lamp that emits light of 340 nm only).
Spectrophotometric Assay
This test is also reliable for the detection of hemizygous males and homozygous females. The detection of heterozygous females raises problems, because the population of healthy erythrocytes in these women produces NADPH in a way similar to that seen in the fluorescent spot test (Figure 2; Vogels et al. 1986). The sensitivity of this test for the detection of heterozygous women is 11%, whereas the specificity is 99% (Table 2). The spectrophotometric assay requires a spectrophotometer.
Cytochemical Assay
The first cytochemical assay was developed by Fairbanks and Lampe (1968). In this test, G6PD activity causes staining of the individual erythrocyte by converting exogenous G6P and NADP+ (Figure 2). Unstained erythrocytes have little or no G6PD activity. The percentage of stained and unstained cells can be determined by light microscopy. Leakage of colored end products from healthy erythrocytes into the medium and unstained cells makes the test unreliable, but optimization of the staining procedure by Van Noorden et al. (1982) improves the reliability of the test. Leakage of the colored end product is largely prevented.
The cytochemical assay is based on the reduction of water-soluble colorless tetranitro blue tetrazolium via the electron carrier 1-methoxyphenazine methosulfate, in its water-insoluble dark-colored formazan by NADPH (Van Noorden et al. 1982; Van Noorden and Vogels 1985). Dark-purple granules are present in erythrocytes that contain G6PD activity, whereas G6PD-deficient erythrocytes remain unstained (Figure 1).
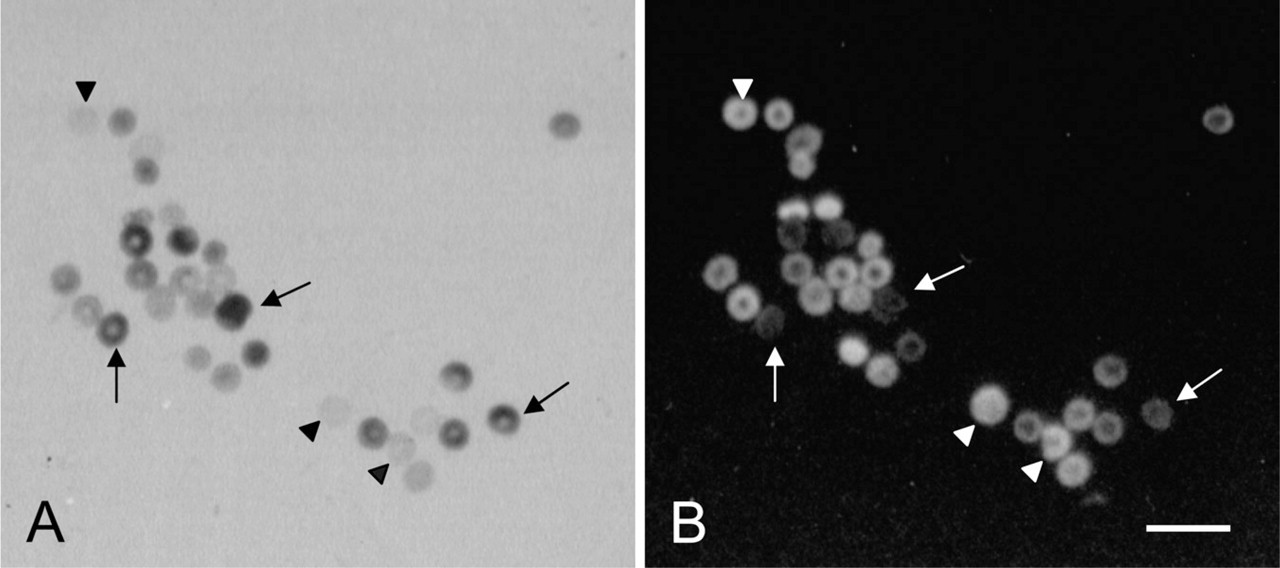
The cytochemical assay is reliable for detection of hemizygously, homozygously, and heterozygously-deficient patients because it shows G6PD activity in individual erythrocytes (Figure 1; Gurbuz et al. 2005). When the percentage of stained and unstained cells has to be determined manually, at least 1000 erythrocytes have to be included in each count to estimate G6PD deficiency. This procedure is time-consuming and prone to error. Van Noorden et al. (1989) developed a new procedure for the evaluation of positive and negative cells using flow cytofluorometric analysis. Formazan crystals in erythrocytes quench the autofluorescence of erythrocytes. Therefore, cells that lack G6PD activity, and thus do not contain formazan crystals, show strong auto-fluorescence. The more formazan crystals are present in cells, the less fluorescence can be observed (Figures 4 and 5). On this basis, percentages of stained and unstained cells can be determined objectively.
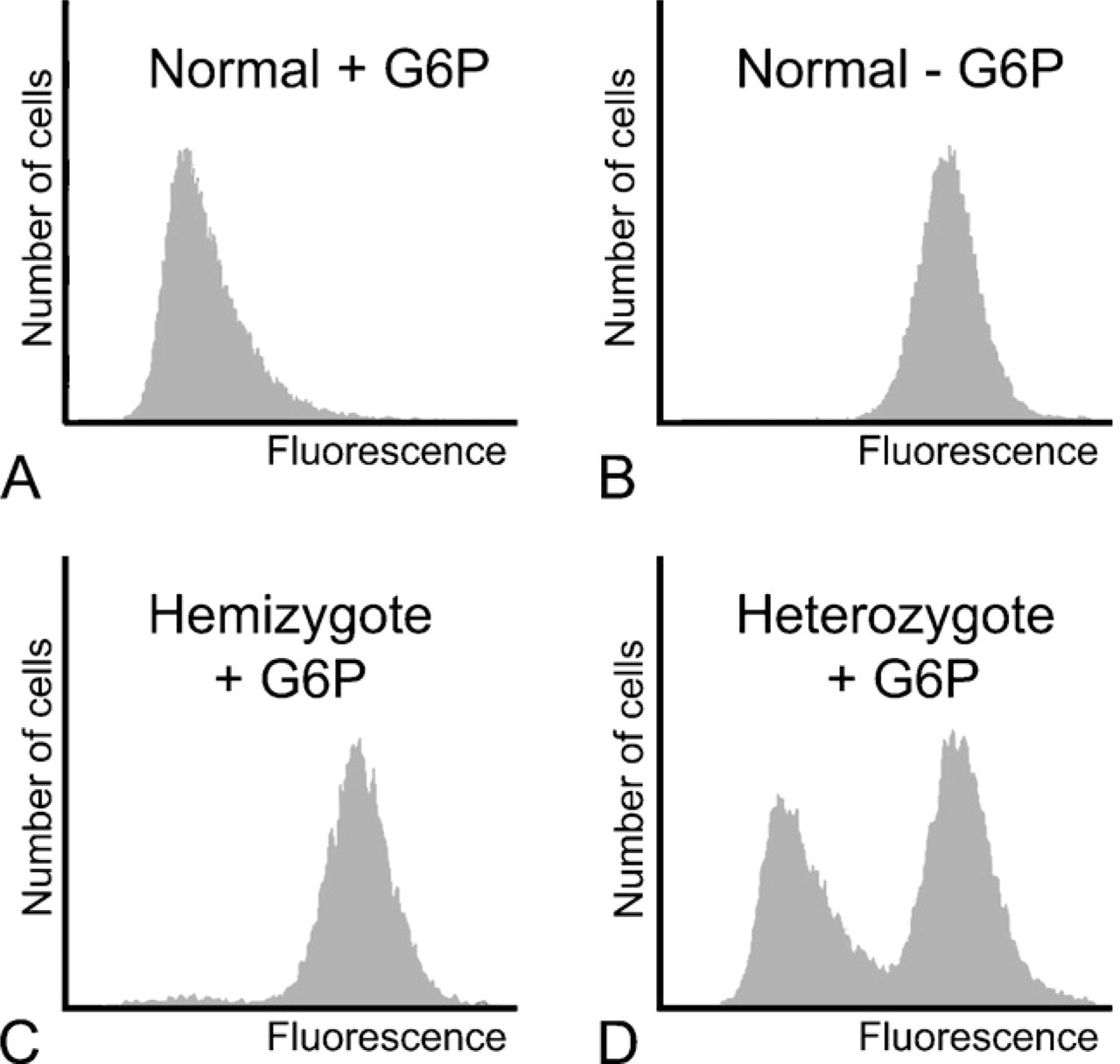
Flow cytometric analysis of erythrocytes that have been stained by the cytochemical assay for the detection of G6PD activity. The final colored reaction product of G6PD activity, formazan, quenches fluorescence. The highly fluorescent cells are G6PD negative. (
The cytochemical assay can detect all forms of G6PD deficiency in erythrocytes of patients. However, the assay has the disadvantage that in comparison with the fluorescent spot test and the spectrophotometric assay, it is a time-consuming and technically difficult test. The assay takes almost 3 hr to perform, and many different steps have to be carried out. The cytochemical assay should be simplified to make it applicable for routine use.
Conclusion
G6PD deficiency causes problems primarily when the deficiency is complicated by the treatment of malaria. Treatment can cause (severe) hemolysis in G6PD-deficient patients. Therefore, patients should be screened for G6PD deficiency before treatment with these potential hemolytic agents. The test used for this purpose should be inexpensive and easy to perform, but also should diagnose heterozygous women reliably. The fluorescent spot test, the spectrophotometric assay, and the cytochemical assay are all relatively inexpensive. They cost one to five dollars; the fluorescent spot test is the least expensive, and the cytochemical assay is the most expensive. The fluorescent spot test is easiest to perform but is very unreliable for the detection of heterozygous women. The cytochemical assay detects heterozygous women reliably, but is cumbersome.
For the best of both worlds, the authors recommend the use of two different tests for the detection of G6PD deficiency in men and women: the inexpensive fluorescent spot test is appropriate for the detection of G6PD deficiency in men, whereas a simplified cytochemical assay should be used for the detection of G6PD deficiency in women to discriminate between healthy, heterozygously-deficient, and homozygously-deficient women.