
Abstract
We tested the hypothesis that mechanical loading of human bone increases expression of the transcription factor RUNX2 and bone matrix proteins osteopontin (OPN), bone sialoprotein (BSP), dentin matrix protein-1 (DMP1), and matrix extracellular phosphoglycoprotein (MEPE). We examined this in tissue sections of atrophic mandibular bone taken from edentulous patients who had undergone distraction osteogenesis. In undistracted bone, weak to moderate staining for OPN and BSP was found in osteoblasts and bone matrix of immature woven bone. RUNX2 was also detectable in osteoblasts and in cells of the periosteum. In woven bone, but not in lamellar bone, a small number of osteocytes stained for all proteins tested. After distraction, staining intensity had increased in the existing old bone and staining was seen in more bone cells than before distraction. We also found a high expression of DMP1 and MEPE in many osteocytes embedded in woven bone and in some osteocytes of lamellar bone not seen before distraction. New bone trabeculae were forming in the fibrous tissue of the distraction gap containing all stages of intramembranous bone formation. Moderate to strong staining was seen for all five proteins tested in osteocytes located in woven bone of these trabeculae and for RUNX2, OPN, and BSP in osteoblasts lining the trabecular surfaces. We conclude that loading of atrophic human jawbone by distraction activates matrix synthesis of bone cells in and around existing bone. Increased staining of DMP1 and MEPE in osteocytes after loading is in line with the concept that these proteins may be involved in signaling the effector cells to adapt the bone structure to its mechanical demands.
Keywords
D
Despite extensive experimental studies in animals and the clinical success of DO, detailed cellular and molecular information on human bone regeneration of distracted bone and the role of mechanical loading is lacking. Cellular and molecular aspects in different tissues and cell types during human bone regeneration can be examined in greater detail by histology rather than by non-invasive methods but require obtaining bone samples from regenerating areas. From an ethical point of view, sampling bone biopsies in orthopedic surgery is not possible without considerable discomfort or morbidity of the patients. However, in craniofacial surgery, DO of the mandible is usually followed by placement of dental implants for which holes need to be drilled in the bone. By using a hollow trephine burr, the investigator can take bone biopsies of the regenerated bone without additional discomfort to the patients. In this way we could carry out a prospective histomorphometric analysis of bone and blood vessel formation in the distraction gap from atrophic mandibular bones of edentulous elderly patients (Amir et al. 2006a, b).
Regeneration of bone shares many similarities with bone formation (Ferguson et al. 1999). The onset of bone formation is marked by the deposition of osteoid, an unmineralized extracellular matrix containing collagen type I that is synthesized and secreted by bone-forming cells, the osteoblasts. Following osteoid deposition, mineralization takes place, which turns the osteoid into bone that makes it function as a load-bearing tissue. During the transformation of osteoid into bone, some of the osteoblasts become incorporated in the bone and become osteocytes, by far the most abundant bone cell type. These cells form a network throughout the bone tissue through their cell processes that run in bone canaliculi. These cell processes are in physical contact with each other and with the cells lining the outer surface of bone. Osteocytes are believed to perceive the stress put upon the bone tissue leading to deformation (strain) and to transfer signals through their canalicular network to the effector cells at the bone surface, the bone-forming osteoblasts, and bone-degrading osteoclasts (Burger and Klein-Nulend 1999).
Several non-collagenous proteins (NCPs) found in bone are involved in formation, maintenance, and turnover of bone and, hence, in adaptation of bone structure to changing endocrine conditions or mechanical loading. One group of these NCPs is termed the small integrin-binding ligand, N-linked glycoprotein (SIBLING) family and includes osteopontin (OPN), bone sialoprotein (BSP), dentin matrix protein-1 (DMP1), and matrix extracellular phosphoglycoprotein (MEPE, also called osteoblast/osteocyte factor- 45 or OF45) (Fisher et al. 2001). The functions of these proteins have not yet been completely clarified, but they are believed to be multifunctional and involved in aspects as diverse as nucleating bone crystals (BSP, phosphorylated DMP1), acting as transcription factor (unphosphorylated DMP1), modulating the rate of crystal growth (OPN and MEPE fragments), anchoring the osteoblasts and osteoclasts to the matrix (OPN), or activating matrix metalloproteinases (MMPs) (for reviews, see Qin et al. 2004; Alford and Hankenson 2006). In particular, DMP1 recently received much attention because it was shown that it influences bone mineralization not only directly by providing extracellular phosphorylated matrix proteins, but also indirectly by regulating serum phosphate levels by controlling secretion of a soluble signal (FGF23) to renal cells (Feng et al. 2006; Lorenz-Depiereux et al. 2006). In addition, the DMP1 gene contains load-sensitive regulatory elements that puts its transcription under control of mechanical loading (Gluhak-Heinrich et al. 2003; Yang et al. 2005). Expression of these SIBLING proteins has been studied predominantly in developing and adult bone of rodents (McKee and Nanci 1995a, b; Petersen et al. 2000; Toyosawa et al. 2001). Studies recently also demonstrated SIBLING protein expression in non-skeletal tissues such as in kidney, salivary glands, and a variety of cancers (Fisher et al. 2004; Ogbureke and Fisher 2004, 2005; Toyosawa et al. 2004c). Apart from OPN- and BSP-localization studies in adult bone (Ingram et al. 1993; McKee and Nanci 1995a, b), little detailed information is available on the expression and role of SIBLING proteins in human bone.
We previously showed that human mandibular bone that undergoes mechanical loading by distraction formed substantially more osteoid than undistracted bone (Amir et al. 2006a). This suggested that, by distraction, cells along or near the bone surface become activated to deposit new layers of bone against existing bone. In the present study we tested the hypothesis that mechanical loading activates the expression of the SIBLING proteins. To this end we immunolocalized OPN, BSP, DMP1, and MEPE in biopsies from mandibles of elderly patients who had undergone vertical distraction osteogenesis prior to insertion of dental implants. We also collected bone biopsies from two patients before their mandibles were osteotomized in order to examine the effect of traction on the protein expression before and after DO. Apart from SIBLING proteins, we examined RUNX-2 localization as an early bone cell marker to detect cells with osteogenic potential present in the soft fibrous tissue area of the distraction gap. RUNX2 is a transcription factor essential for osteoblast differentiation and for maintaining their differentiated state because Runx2 is needed for gene expression of collagen type I and for most of the SIBLING proteins (Komori et al. 1997; Otto et al. 1997; Karsenty et al. 1999).
Material and Methods
Clinical Procedure
Patient selection and surgical procedures have been described in detail (Amir et al. 2006a). Briefly, 10 edentulous patients (between 52 and 74 years of age) with severely atrophic mandibles (∼8-mm residual height) underwent anterior alveolar bone augmentation by vertical distraction osteogenesis using extraosseous distractors (KLS Martin and Mondeal Medical Systems; Tuttlingen, Germany). Pre-distraction bone biopsies were taken from two patients to compare with postdistraction bone (see Amir et al. 2006a). After performing an osteotomy and placing the distractor, a latency period of 5–7 days was introduced to enable callus formation. Next, the distractor was activated, moving the bone segment approximately 10 mm upward (Figure 1A). The rate of distraction was either 0.5 or 1.0 mm/day (2 × 0.5 mm), and the bone segments were maintained in neutral position without activation for 10–22 weeks (neutrofixation period) to allow the formation of new bone in the distraction gap. The selection criteria of patients included good general physical and mental health, no drug and/or alcohol abuse, no smoking, and no systemic diseases that could interfere with normal wound healing.
At the time of removal of the distractor at the end of the neutrofixation period, a hollow trephine burr with 3.5-mm outer diameter (2-mm inner diameter; Straumann, Waldenburg, Switzerland) was used for implant preparation vertically through the bone segments and distracted area. The inner core of the trephine was collected as a biopsy sample, and dental implants were placed at the site prepared with the trephine burr. All procedures were approved by the Medical Ethical Committee of Northern Holland, and all patients gave informed consent.
Fixation and Embedding
Biopsies were fixed for 24 hr in 4% formaldehyde in 0.1 M phosphate buffer solution and decalcified with 5% EDTA and 1% formaldehyde, pH 7.3, at room temperature for 8 weeks. After rinsing with 0.1 M phosphate buffer, these biopsies were dehydrated through ethanol 90% and 100%, cleared twice in xylene, and finally embedded in molten paraffin at 56C. Five- to 6-μm-thick sections were cut with a microtome, placed on poly-L-lysine-coated glass slides, and dried overnight at 37C to be later used for immunohistochemistry.
Immunohistochemical Procedures
Tissue sections were deparaffinized in xylene and rehydrated through a descending series of aqueous ethanol solutions. Endogenous peroxidase was quenched by incubation in 3% H2O2 in PBS for 10 min, followed by washing three times in PBS, pH 7.4, with 0.05% Tween 20 (PBS-T). Sections were blocked with appropriate blocking sera supplied by the manufacturer of the kits (see below) and incubated for 2 hr with primary antibodies to OPN, BSP, and RUNX2 in PBS in a humidified chamber at room temperature. Antibodies were diluted in PBS with 10% normal goat serum and were specified as reported (Table 1) (Fisher et al. 1995; Ogbureke and Fisher 2004, 2005; Toyosawa et al. 2004c; Bronckers et al. 2005). To visualize DMP1 and MEPE, epitopes were first retrieved by enzyme digestion with trypsin for 10 min at 37C followed by a blocking step according to the manufacturer's protocol (Zymed; South San Francisco, CA). Sections were incubated with primary antibodies overnight at 4C in a moist chamber. After washing with PBS-T, SuperPicTure Polymer horseradish peroxidase-conjugated broad-spectrum secondary antibody (Zymed) was added, and the sections were incubated at room temperature for 10 min. The sections were washed again with PBS-T and developed with 3,3′ diaminobenzidine substrate solution for a maximum of 10 min. Sections were counter-stained with methyl green or Mayer's hematoxylin for 1 min, mounted, and covered with a glass coverslip. As negative controls, sections were incubated with normal IgG replacing the primary antibody.
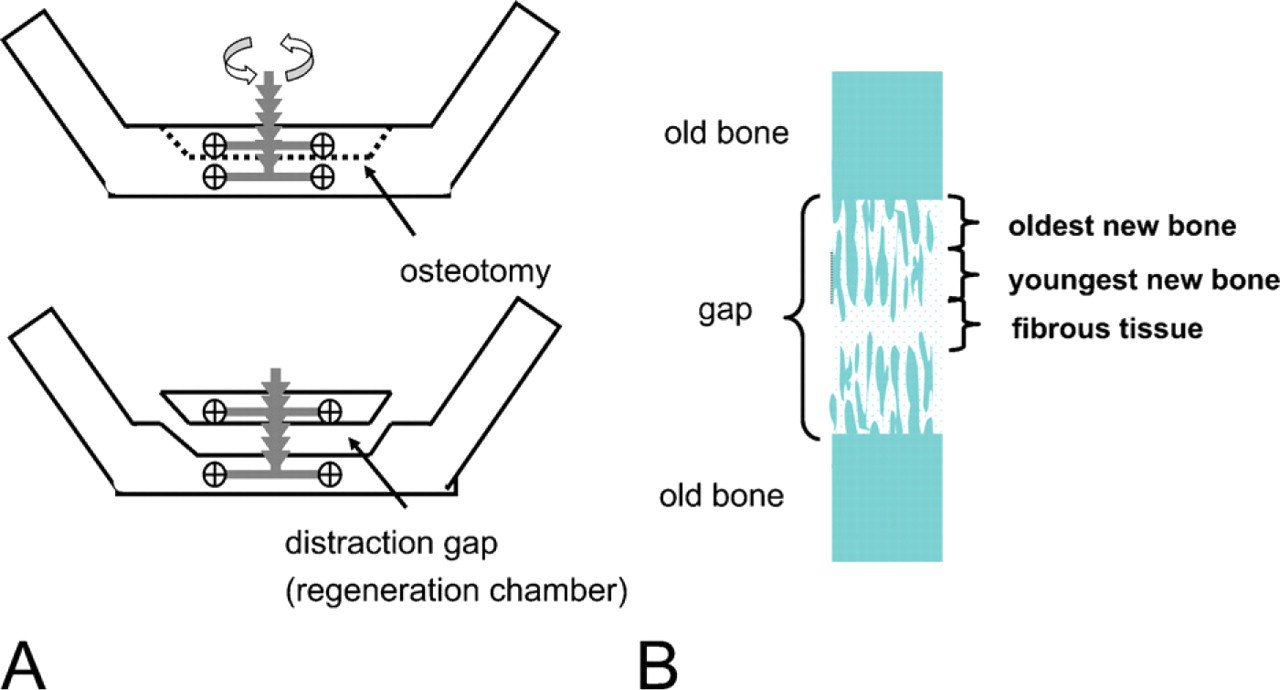
Vertical distraction osteogenesis in the human mandible (
Staining Analysis
Staining analysis was done in two main areas in each distracted biopsy (Figure 1B). The first main area was the old bone of the transport segment, already present before osteotomy and distraction were applied. The second main area was the distraction gap area in which new bone was formed as bone trabeculae (bone columns) parallel to the distraction vector. These trabeculae had started to grow from the osteotomy line (still visible) into the soft connective tissue of the gap and contained the ‘oldest’ new bone. Progressively younger (and thinner) trabecular bone was found more distant from the osteotomy line towards the center of the gap. The earliest stages of bone formation were found near the growing tips of the trabeculae. Photographs were taken with an AxioCam MR-MRGrab camera imaging system (Carl Zeiss Vision; Munich, Germany), which included an Axioplan2 microscope, an AxioCam MR camera, and Axio Vision 4.4 software. For both of the main areas, the following were examined: whether immunostaining was intracellular and/or extracellular, which cells stained (preosteoblasts, osteoblasts, osteocytes, osteoclasts), which type of matrix stained (osteoid, woven bone, lamellar bone), and in which part of the bone staining was noted (preexisting bone, new bone in distraction gap). Mature lamellar bone was discriminated from immature woven bone on the basis of polarized light microscopy. Approximately five tissue sections of each biopsy and each antibody were examined. Staining intensity was assessed visually and estimated from weak (+/−) to strong (+++).
Specifications of primary antibodies

e Stock antibodies were diluted with 10% normal goat serum in PBS.
OPN, osteopontin; BSP, bone sialoprotein; DMP1, dentin matrix protein-1; MEPE, matrix extracellular phosphoglycoprotein.
Results
Predistraction Bone
Biopsies taken before distraction served as controls to establish baseline-staining patterns in atrophic mandibular bone for each of the five proteins. OPN protein was found in cement lines of the osteons, in osteoid, newly formed woven bone around Haversian canals, and in osteoblasts in the periosteum (Figure 2A). Some osteocytes of woven bone near the bone surface, suggesting they were recently embedded, stained positive for OPN as well. BSP protein was localized in woven but not lamellar bone matrix, in some osteocytes of woven bone, and in (few) osteoblasts. MEPE and DMP1 proteins were detected in osteocytes of woven bone in areas near the periosteum and bone marrow (BM) spaces but were not, or very sparingly, seen in osteoblasts (Figures 2B–2D). Strongest expression of DMP1 and MEPE was identified in the lacunar wall of the osteocytes of woven bone. No MEPE- or DMP1-positive staining was found in (flattened) osteocytes in more mature lamellar bone located farther away from BM spaces or from the periosteum. RUNX2-positive staining was localized exclusively cytoplasmic and/or nuclear in some osteocytes of woven bone, osteoblasts, marrow stromal cells, and in the cells of the periosteum. A summary is presented in Table 2.
Postdistraction (Old) Bone
The presence of an osteotomy line in the bone enabled us to make a clear distinction between the area with predistraction (old) bone (mainly dense lamellar type) and the gap area with bone trabeculae (woven and lamellar bone) that had formed by distraction. The latter will be discussed under ‘distraction gap’ (see below).
After distraction, distribution of OPN in the existing bone was similar as before distraction, but now more cells were positive and staining was stronger than before distraction, mainly in the woven bone deposited against the (old) lamellar bone. This was principally seen in the zones near the osteotomy line and the mucosa but less in the more central zone in between (Figures 3A–3C).
Extracellular expression of BSP closely followed that of OPN except for cement lines where BSP was seen only at the interface between lamellar and woven bone but not in cement lines of the osteons (Figure 3D). At the periphery of the marrow spaces, new bone had been deposited against old bone and replaced areas of non-vital bone containing empty osteocyte lacunae (Figure 3B). The osteotomy line stained strongly for OPN and BSP protein and was thicker than regular cement lines (Figure 3B). New bone with positive staining for OPN and BSP was also observed within bone marrow spaces containing adipose tissue. Areas with high bone-forming activity contained many OPN- and BSP-positive osteoblasts, osteoid, and young osteocytes. Some OPN- and (few) BSP-positive osteoclasts (Figure 3C for OPN) were noted in areas where bone resorption was high, some of which presumably caused by phagocytosis (Bronckers et al. 2003). In young osteocytes, BSP expression was stronger than in osteoblasts (Figure 3D).
MEPE and DMP1 protein was found almost exclusively in osteocytes. MEPE staining was also seen in a few fully functional osteoblasts but much weaker than in osteocytes. Compared with their expression in pre-distraction bone, after distraction MEPE- and DMP1-stained cells were located more widely throughout the bone and seen in osteocytes of both woven and lamellar bone (Figure 3E). Lamellar bone osteocytes had a flattened shape parallel to the bone lamellae with scarce cytoplasm, whereas woven bone osteocytes had a round shape with abundant cytoplasm, with many dendrite-like cytoplasmic projections and distributed in high density around bone marrow spaces. Strongest staining for MEPE and DMP1 was found in the walls of osteocyte lacunae and in canaliculi of woven bone osteocytes.
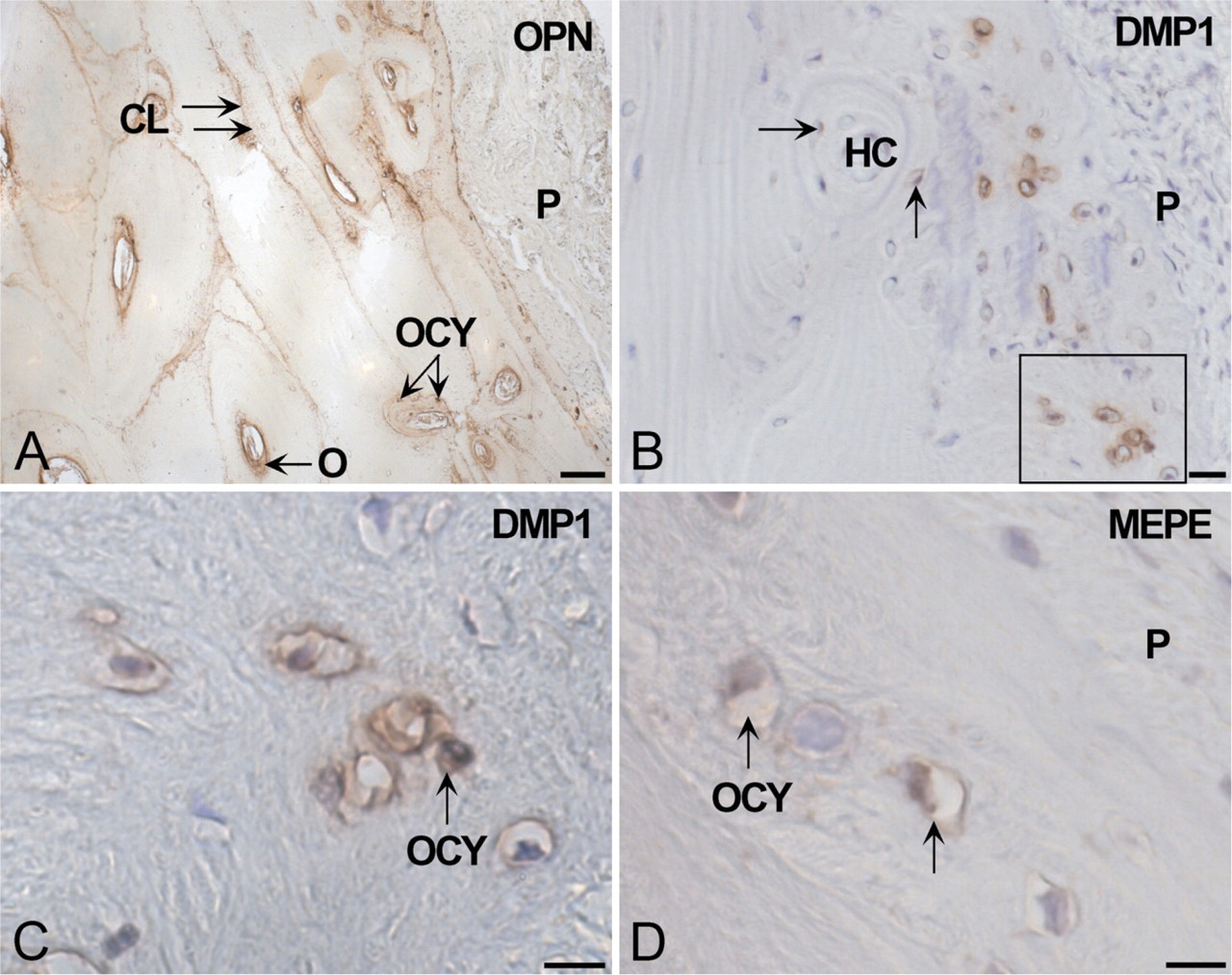
Immunolocalization of SIBLING proteins in undistracted bone (baseline levels). (
RUNX2 protein was found in a variety of cells in bone marrow spaces, most periosteal cells, osteoblasts, and osteocytes of woven bone (Figures 3F and 3G). Control sections in which primary antibodies had been replaced were negative (Figure 3H and Figure 4L).
Distraction Gap
New bone trabeculae had formed starting at the osteotomy line and extended into the distraction gap parallel to the traction vector. In these trabeculae the distribution pattern of OPN and BSP staining in bone matrix changed as the newly formed bone columns grew older, going from the fibrous tissue in the center of the gap (youngest part) to the osteotomy line (oldest part). In the oldest part of these trabeculae near the osteotomy line, extracellular staining of the bone matrix for OPN and BSP was weak and seen only in the core (Figure 4A). This core consisted of woven bone as assessed by polarized light microscopy (Figure 4B). The peripheral lamellar bone matrix deposited against this inner core of woven bone stained much weaker or not at all for OPN and BSP.
Distribution and intensity of staining of SIBLINGs and RUNX2 protein in the old bone and in the new bone formed in the gap after distraction
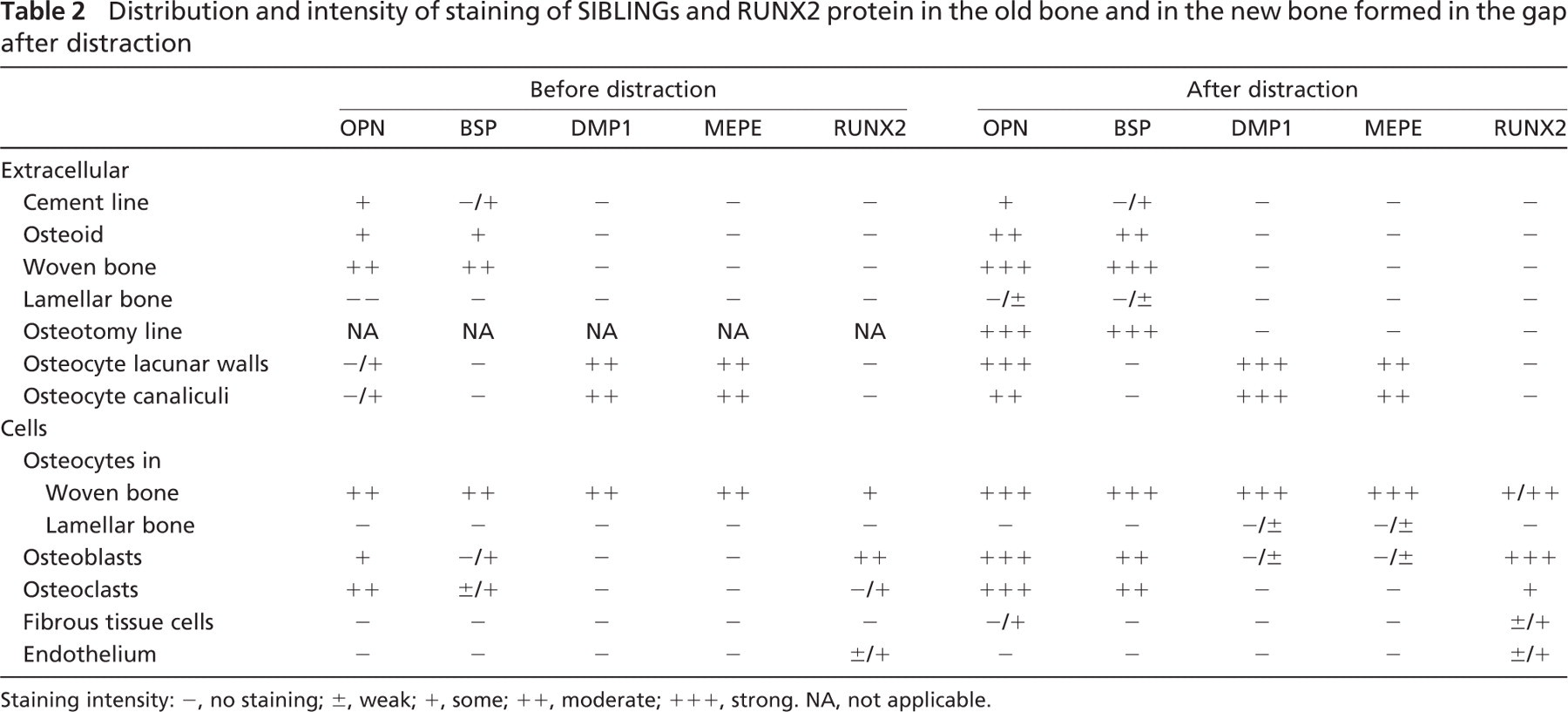
Staining intensity: −, no staining; ±, weak; +, some; ++, moderate; +++, strong. NA, not applicable.
Less lamellar bone had been deposited against the core of woven bone in younger parts of the trabeculae (halfway between the osteotomy line and growth area of trabeculae), whereas the youngest part consisted of woven bone only. These observations were seen in tissue sections with a neutrofixation period of ∼12 weeks. The younger the trabeculae, the stronger and more evenly distributed the staining for OPN and BSP in the woven bone in the core (Figures 4C and 4D). In biopsies taken at later time points (neutrofixation up to 22 weeks), OPN-positive cement lines were observed in the oldest part of the newly formed trabecula, whereas in the younger parts OPN-positive woven bone was still present.
OPN and BSP staining was also noted intracellularly in osteoblasts lining the trabeculae surface and in a few osteoclasts (Figure 4E). Elongated connective tissue cells (presumably preosteoblasts and other cells of the periosteum) surrounding the trabeculae stained for OPN (Figure 4E) with less intensity than osteoblasts but did not stain for BSP. In woven bone, the density of osteocytes was rather high, their shape irregular, and staining for BSP and OPN strong. In lamellar bone, however, osteocyte density was lower, and staining for these two proteins was weaker. Osteoclasts (large multinuclear cells near a resorption pit) localized mostly in the oldest part of the trabeculae near the osteotomy line and were strongly stained for OPN.
In the trabeculae, high expression of MEPE and DMP1 protein (as for OPN and BSP) was also found in osteocytes of woven bone but less or no staining in (flattened) osteocytes of lamellar bone; cytoplasm, canaliculi (or osteocytic processes), and osteocyte lacunar walls stained positive for MEPE and DMP1 (Figures 4F–4I). Occasionally, weak staining for MEPE and DMP1 was also noted in a few osteoblasts, but most of these cells were negative (Figures 4F–4I). No differences in distribution were noted between the monoclonal and polyclonal anti-DMP1 antibodies (Figures 4F and 4G).
Virtually all fibrous tissue cells that surrounded the trabeculae stained for RUNX2, a marker for cells with osteogenic potential (Figures 4J and 4K). Strongest expression was found in osteoblasts. Many young osteocytes embedded in osteoid or still located close to the bone surface were also positive. RUNX2-positive cells were scattered throughout the fibrous tissue area in the center of the gap. Staining intensity decreased in the area farthest away from the youngest new-bone zone. No histological evidence was found for the presence of chondrocytes or cartilage in the gap zone, and no positive staining for cartilage was seen after staining with safranin-O or toluidine blue (not shown).
A summary of the expression patterns in the various cells and tissue layers is presented in Table 2.
Discussion
We report here that mechanical loading of bone, as seen in the upper bone segment after distraction, enhances the number of bone cells immunoreactive for SIBLING proteins. After distraction, more (pre)osteoblasts lining Haversian canals stained for RUNX2, OPN, and BSP, and more osteoid and bone stained for OPN and BSP than before distraction. This suggested that mechanical loading increases the number and/or activity of bone cells that synthesize and deposit bone extracellular matrix proteins. This was in line with our morphometric data showing that four times as much osteoid is present in old, atrophic human bone after distraction than before distraction (Amir et al. 2006a). After loading, some of the mature osteocytes embedded in the old lamellar bone were also positive for DMP1 and MEPE, an observation not seen in the samples taken before distraction. Thus, all data are consistent with the concept that mechanical loading of bone stimulates bone formation by (re)activating bone cells.
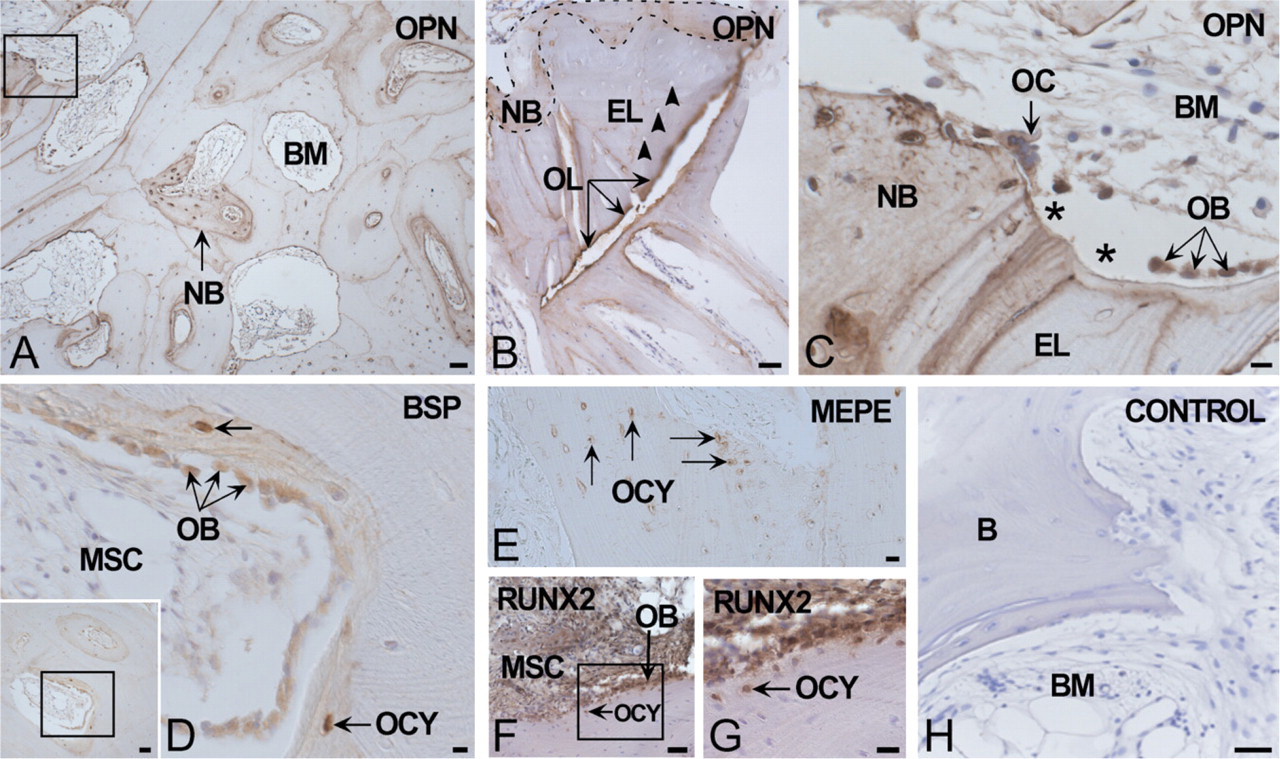
Immunolocalization of SIBLING proteins in the postdistracted old bone. (
Our studies also show that tension on the soft connective tissue callus between two separate bone segments recruits cells in the callus tissue to become bone cells, induces expression of ECM proteins, and promotes de novo formation of bone trabeculae to fill the gap. We found no histological evidence for any cartilage formed as an intermediate tissue prior to deposition of bone, suggesting that in the gap the trabeculae had formed directly by intramembranous ossification. This finding differs with reports on distraction osteogenesis in long bones that show cartilage formation preceding bone formation at an early stage of healing (Yasui et al. 1997; Sato et al. 1998; Li et al. 2000). One could argue that cartilage was present only initially but disappeared by remodeling (Sato et al. 1998). Because we could not find any cartilage remnants in the gap area after staining with safranin-O and toluidine blue either in the present or previous study (Amir et al. 2006a) and because bone remodeling in human is relatively slow, it is likely that the trabeculae were formed intramembranously, as happens during embryonic development of jawbone.
Formation of bone in the gap area during distraction osteogenesis in the human jaw closely resembles embryonic ossification and normal fracture healing (Sato et al. 1998; Ferguson et al. 1999; Li et al. 2000; Toyosawa et al. 2004a). mRNA expression of BMPs in the gap area in rat long bones is substantially higher in healing during distraction than during static fracture healing or embryonic bone formation (Sato et al. 1998). Although in our study we could not compare healing in static fractures with distracted ones, we were able to compare distracted with undistracted bone and found more intense and more extended protein staining in the distracted situation. Thus, our data are consistent with the concept that mechanical loading of the soft tissue in the gap is likely to increase matrix synthetic activity of bone-forming cells more than unloaded static fracture healing.
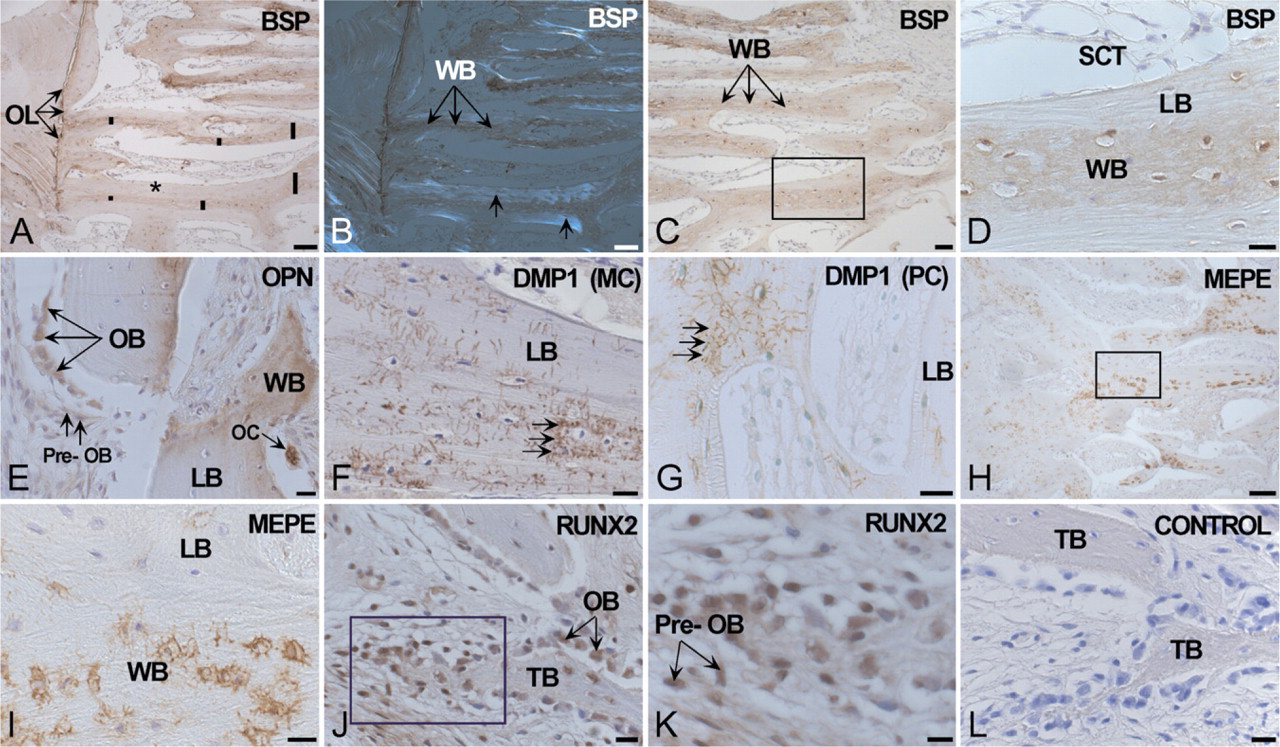
Immunolocalization of SIBLING proteins in bone trabeculae formed in the distraction gap. (
The function of these SIBLING proteins in bone formation and resorption is not yet clear. BSP and OPN are relatively restricted to osteoblasts and osteoclasts participating in active remodeling (for review, see Alford and Hankenson 2006). MEPE and DMP1, however, are restricted to osteocytes (Toyosawa et al. 2001; Feng et al. 2003; Ye et al. 2005). None of the four bone matrix protein species examined here seems essential for bone formation as suggested by single knockout studies for SIBLING members in mice (Aubin et al. 1996; Rittling et al. 1998; Boskey et al. 2002; Feng et al. 2003; Gowen et al. 2003; Ye et al. 2005; Ling et al. 2005). Some functions of SIBLING members only became apparent after challenging single null mutant mice, such as by mechanical disuse of bone. For instance, non-loading of long bones in tail-suspended wild-type mice reduces bone formation and enhances bone resorption. However, in OPN-deficient, tail-suspended mice, bone formation in non-loaded long bones is not reduced and bone resorption is not increased (Ishijima et al. 2001). These data suggest that OPN is a molecule required for the bone loss induced by mechanical disuse that regulates the functions of osteoblasts and osteoclasts. Loss-of-function studies do suggest that SIBLING proteins have overlapping functions in bone formation and bone resorption and may compensate for each other.
The increased exclusive expression of MEPE and DMP1 in osteocytes after bone distraction and the mechanosensitivity of osteocytes (Klein Nulend et al. 1995), along with the report that the DMP1 gene contains mechanosensitive regulatory elements (Gluhak-Heinrich et al. 2003; Yang et al. 2005), suggest that these two SIBLING proteins likely play a role in adaptation of bone structure to loading. MEPE and DMP1 are multifunctional proteins (Narayanan et al. 2003, 2006; Rowe et al. 2004; Tartaix et al. 2004). DMP1 has originally been associated directly with biomineralization (George et al. 1994; Toyosawa et al. 2004a, b) both by stimulating and inhibiting mineral deposition, depending on protein size and degree of phosphorylation (Tartaix et al. 2004; Toyosawa et al. 2004a; Yang et al. 2005). DMP1 also indirectly acts on biomineralization by regulating serum phosphate levels via its control of FGF23 secretion, a signal from osteocytes to the kidney, to influence the reabsorption of phosphate in kidney (Feng et al. 2006; Lorenz-Depiereux et al. 2006). Also, MEPE has been proposed to act as a signaling molecule that inhibits the activity and/or recruitment of new osteoblasts (Gowen et al. 2003). The presence of mechanosensitive regulatory elements in the DMP1 gene (Yang et al. 2005) brings the expression of this gene under control of mechanical loading. Hence, it is conceivable that mechanical loading enhancesDMP1 and MEPE expression in osteocytes, which then act as signal molecules to effector cells to adapt the bone structure.
In short, bone deformation during distraction osteogenesis in atrophic mandibular human bone (re)activates bone cells in existing bone and induces protein expression of SIBLING proteins. Also, in human bone, MEPE and DMP1 are found exclusively in osteocytes. The exclusive location of DMP1 and MEPE in and around osteocytes suggests that these proteins act either to locally influence mineralization or that they are soluble signals of osteocytes to influence activity of the effector cells to adapt bone structure to mechanical demands.
Footnotes
Acknowledgements
The authors are grateful to Dr. Larry Fisher, National Institutes of Health (NIH), Bethesda, MD for generously providing antibodies to OPN, BSP, DMP1, and MEPE and for helpful discussions and support to the first author (LA) during her visit to NIH, to Dr. Kenichi Sasaguri (Kanagawa Dental College, Yokosuka City, Kanagawa, Japan) for providing anti-RUNX2 antibodies, to Dr. Michiko Yuki (Department of Oral Pathology, Osaka University, Osaka, Japan), and to Mr. Dirk-Jan Bervoets (Oral Cell Biology ACTA, Amsterdam) for excellent technical assistance.