
Abstract
The SIBLING (Small Integrin-Binding LIgand, N-linked Glycoprotein) family of secreted glycophosphoproteins includes bone sialoprotein (BSP), dentin matrix protein-1 (DMP1), dentin sialophosphoprotein (DSPP), osteopontin (OPN), and matrix extracellular phosphoglycoprotein (MEPE). For many years, they were thought in normal adults to essentially be limited to metabolically active mesenchymal cells that assembled the mineralized matrices of bones and teeth. Over the last decade they have also been upregulated in a variety of tumors. Three of these proteins (BSP, OPN, and DMP1) have been shown to interact with three matrix metalloproteinases (MMP-2, MMP-3, and MMP-9, respectively). Recently, all five SIBLINGs and their MMP partners when known were observed in specific elements of normal ductal epithelia in salivary gland and kidney. We have hypothesized that the SIBLINGs and their MMP partners may be expressed in ductal cells with high metabolic activity. In this paper, we show that all the SIBLINGs (except MEPE) and their MMP partners are expressed in the metabolically active epithelia of human eccrine sweat gland duct but not in the more passive ductal cells of the macaque (monkey) lacrimal gland. It is hypothesized that MEPE expression may be limited to cells involved in active phosphate transport. This manuscript contains online supplemental material at http://www.jhc.org. Please visit this article online to view these materials.
Keywords
B
Except for OPN, expression of the SIBLINGs in normal tissues was generally thought to be limited to bones and teeth (Robey 2002; Qin et al. 2004). However, some earlier reports indicated the expression of OPN in human breast milk (Senger et al. 1989), chronic inflammatory cells (Patarca et al. 1990), and kidney (Shiraga et al. 1992), as well as some other epithelia (Brown et al. 1992). Terasawa et al. (2004) have also reported expression of DMP1 in several mouse soft tissues including liver, muscle, brain, pancreas, and kidney, by immunohistochemistry (IHC) and RT-PCR. Recently it was shown that all five members of the SIBLING family of proteins are expressed in the duct cells of the salivary gland (Ogbureke and Fisher 2004) and kidney (Ogbureke and Fisher 2005). Equally significant, these reports showed that three of the SIBLINGs (BSP, OPN, and DMP1) consistently colocalized with their specific MMP binding partners (MMP-3, MMP-2, and MMP-9, respectively). Using highly purified proteins, it has been shown in vitro that specific SIBLING proteins bind and induce a conformational change in only its partner MMP, and that this conformational change causes the proteases' inhibitory proteins, TIMPs (tissue inhibitor of metalloproteinase), to be ineffective at inhibiting the enzyme at normal concentrations (Fedarko et al. 2004). Furthermore, for at least the BSP/MMP-2 and DMP1/MMP-9 partners, it has been shown that the MMPs are bridged to the cell surface of cancer cell lines and enhance the invasion potential of the cells by digesting local protein matrix barriers (Karadag et al. 2004,2005). The roles of these SIBLING/MMP complexes in normal tissues, however, remain unclear.
Eccrine sweat glands perform an important thermo-regulatory function of the body. They are distributed nearly widely across the entire human body surface with the exception of the margins of the lips, nail beds, nipples, inner preputial surface, labia minora, glans penis, and glans clitoridis (Groscurth 2002; Kreyden and Scheidegger 2004). Their greatest densities in humans are on the soles of the feet (620 ± 20/cm2), the forehead and the axilla (360 ± 60/cm2), and the palms and the cheek (300 ± 80/cm2). They are least numerous on the trunk (65 ± 20/cm2) and the extremities (120 ± 30/cm2) (Groscurth 2002). Although the total number of human sweat glands is somewhere between 2 and 4 million, only ∼5% of sweat glands are active at the same time, indicating the enormous potential for sweat production (Kreyden and Scheidegger 2004).
The eccrine sweat gland is an elongated tubular structure (Figure 1 schematic) with a highly coiled secretory portion (Figures 1C and 1D) and a straight ductular portion (Figure 1B) (Schaller and Plewig 2003). The duct fuses with the base of the epidermal papillae to open via a rounded aperture onto the skin surface, which is visible as a regular series of puncta along the centers of friction ridges (Groscurth 2002). The secretory coil, ∼0.5 mm in diameter, is situated deep in the dermis or hypodermis, and both clear cells and dark cells can be distinguished within its epithelium (Kreyden and Scheidegger 2004). Clear cells secrete the major components of sweat, water, and electrolytes, whereas the dark cells are known to secrete glycoproteins. Other functions of the clear cells, however, remain unknown (Kreyden and Scheidegger 2004).
The initial fluid produced by the secretory portion of the sweat gland is isotonic (comparable to the initial fluid of saliva from salivary gland acini, for example). This fluid is modified in the excretory portion of the duct so that the fluid delivered to the skin surface has low Na+ and Cl− concentrations (30–70 mEq/liter in each case) and an elevated (compared with serum) high K+ (up to 5 mEq/liter), lactate, urea, and ammonia concentrations (Kreyden and Scheidegger 2004). This energy-intensive recovery of salts and other low molecular mass molecules in the distal portion of the ductal system is performed by ductal cells rich in mitochondria.
The lacrimal gland is the major source of fluid-tears-to the surface of the eye. Morphology of the lacrimal gland mirrors that of the salivary gland in that there are distinct acini and ductal structures (Pflugfelder et al. 2000). Unlike the salivary gland, however, the isotonic fluid in tears is not made hypotonic as it passes down the ductal system (Wu et al. 2006). This lack of ion transport is reflected in the comparatively low abundance of mitochondria in the ductal epithelia of the lacrimal gland.
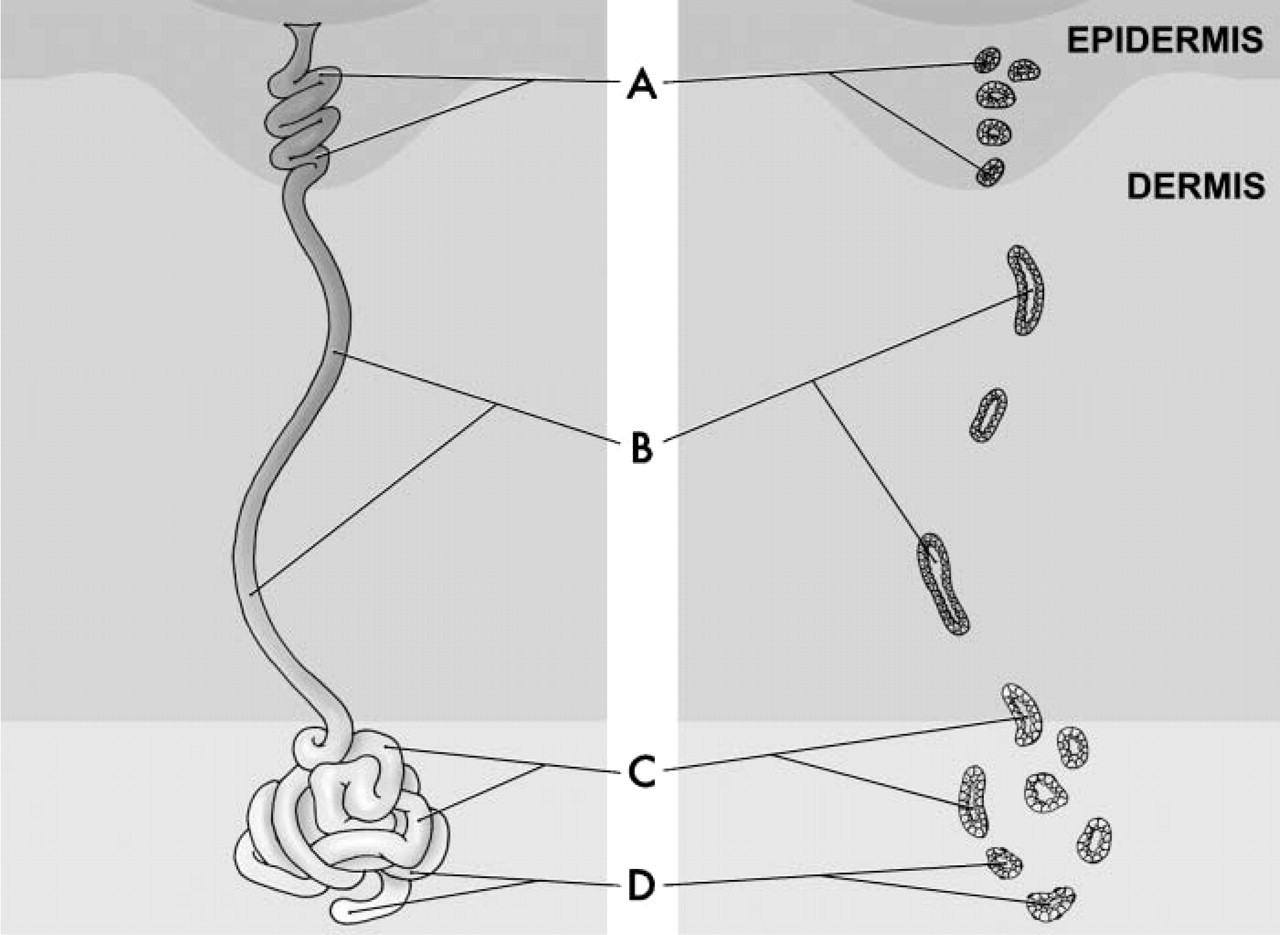
Schematic illustration of the eccrine unit of the human sweat gland. Eccrine units consist of three portions: the acrosyringium or intra-epidermal spiralled duct (
Based on the hypothesis that the SIBLINGs and their partner MMPs (when known) are highly expressing in ductal cells with high metabolic requirements, the present study compared the expression of the SIBLINGs and their cognate MMPs in the sweat gland (high metabolic epithelium) and the lacrimal gland (low metabolic epithelium) by IHC.
Materials and Methods
Tissue Collection
Surgical waste from normal human skin in paraffin blocks without patient identifiers was obtained from the Mid-Atlantic Division of the Human Tissue Network (Charlottesville, VA) under an NIH-approved (exempt) protocol. Fresh surgical waste from lacrimal glands of monkey (M. fascicularis) was obtained from the Tissue Distribution Program of the National Primate Research Center (University of Washington; Seattle, WA). Slices of fresh lacrimal gland tissues were fixed in 10% neutral-buffered formalin for 24 hr and routinely processed for paraffin embedding. Four-μm sections were serially cut from all blocks.
IHC
Mouse monoclonal antibodies (MAbs) and rabbit polyclonal antibodies to the SIBLINGs used in this study are summarized in Table 1. Their methods of production and their effectiveness on paraffin sections of human and monkey tissues have been previously described (Ogbureke and Fisher 2004,2005). The following SIBLING mouse MAbs were used: LFMb-25 for BSP, LFMb-14 for OPN, LFMb-21 for DSPP, LFMb-31 for DMP1, and LFMb-33 for MEPE. Their polyclonal counterparts LF-84, LF-123, LF-151, LF-148, and LF-155, respectively, were used to validate corresponding MAb results. Rabbit polyclonal antibodies to MMP-2 (LF-183), MMP-3 (LF-182), and MMP-9 (LF 184) were made from synthetic peptides in our laboratory (Ogbureke et al. in press). The MAb against mitochondrial protein was obtained commercially (mouse IgG1, cat. #MAb1273; Chemicon, Temecula, CA). Affinity-purified rabbit polyclonal antibody to human aquaporin 5 (AQP5) was a kind donation from Dr. Bruce Baum (NIDCR, NIH, Bethesda, MD) (Wellner et al. 2000,2005).
IHC methods were the same as those recently described for the detection of the SIBLING proteins in salivary and kidney sections (Ogbureke and Fisher 2004,2005). The Zymed ST5050 automated immunostaining system in conjunction with Zymed detection reagents were used with modifications as recently described (Zymed; South San Francisco, CA) (Ogbureke and Fisher 2005). In brief, sections were manually dewaxed in three 5-min xylene washes and rehydrated through graded ethanol (100%, 95%, and 75%) and water. Endogenous peroxidase activity was destroyed by treating the sections with 3% hydrogen peroxide (in methanol) for 30 min. Sections were then washed three times in PBS for at least 5 min each and covered with PBS + 0.05% Tween-20 (PBS-T) before loading the slides onto the preprogrammed ST5050 automated IHC machine.
Sections were incubated for 1 hr with appropriate antibody diluted in 10% normal goat serum in PBS. The sections then underwent a 4 × 1-min wash cycle with PBS-T before incubation with SuperPicTure Polymer HRP-conjugated broad-spectrum secondary antibody (cat. #87–8963; Zymed) for 10 min. After washing, aminoethylcarbazole (AEC Single Solution, cat. #00–1122; Zymed) was used as chromogenic substrate for 2 min before rinsing and manually counterstaining with Mayer's hematoxylin for 10–20 sec. All steps were performed at room temperature. An overlay of Clearmount glaze was applied and, after drying, the slides were coverslipped with Histomount (both from Zymed). Negative controls included the substitution of primary antibody with non-immune rabbit serum or mouse IgG control (cat. #08–6599; Zymed). Positive controls consisted of human salivary gland and monkey kidney sections stained with SIBLING, MMP, mitochondria, or aquaporin antisera. Representative photographic images were captured using the Axioplan2 Universal microscope equipped with an Axiovision digital camera and Axiovision program (Carl Zeiss GmbH; Jena, Germany).
Results
This study surveyed the expression of the SIBLINGs and their known MMP partners in the ductal epithelia of metabolically active eccrine sweat gland and compared this to their expression in the relatively inactive duct of the lacrimal gland. As shown in Figure 2, immunoreactivity was present for BSP, DMP1, DSPP, OPN, MMP-2, MMP-3, and MMP-9, mainly in the ductal cells. The SIBLINGs with known MMP partners, OPN (Figure 2A), BSP (Figure 2B), and DMP1 (Figure 2C) colocalized with their cognate MMPs (MMP-3) (Figure 2D), MMP-2 (Figure 2E), and MMP-9 (Figure 2F), respectively. DSPP expression was uniformly intense in cells of the secretory and excretory eccrine sweat gland (Figure 2G). For all four SIBLINGs, expression was diffusely cytoplasmic and perinuclear. Furthermore, the intensity of immunoreactivity for OPN was observed to be higher in the excretory portion (Figure 2A, asterisk) than in the secretory portion (Figure 2A, arrow) of the sweat gland. There was no immunoreactivity for the fifth member of the SIBLING family, MEPE, in the eccrine sweat glands (not shown). Immunoreactivity for all SIBLINGs and their cognate MMPs was absent in the connective tissue cells and stroma of the eccrine sweat gland. Intense immunoreactivity for human mitochondria (Figure 2H) was included to emphasize the high metabolic potential of the eccrine sweat duct. Similar intensely positive immunoreactivity for human mitochondria was observed in the salivary gland ducts (see supplementary Figure 2S-K) and kidney nephron (not shown) to serve as positive control. Negative control consisted of preimmune mouse IgG1 (Figure 2I) and preimmune rabbit IgG (see supplementary Figure 2S-J) on eccrine sweat glands.
Antibodies used in this study
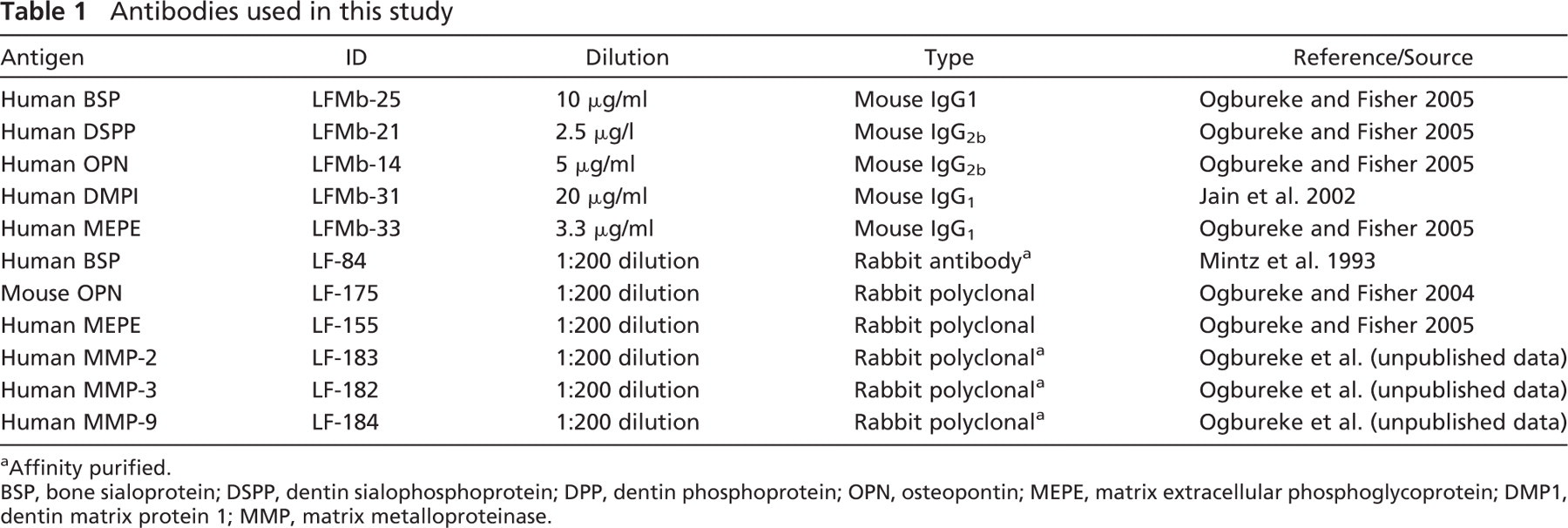
Affinity purified.
BSP, bone sialoprotein; DSPP, dentin sialophosphoprotein; DPP, dentin phosphoprotein; OPN, osteopontin; MEPE, matrix extracellular phosphoglycoprotein; DMP1, dentin matrix protein 1; MMP, matrix metalloproteinase.
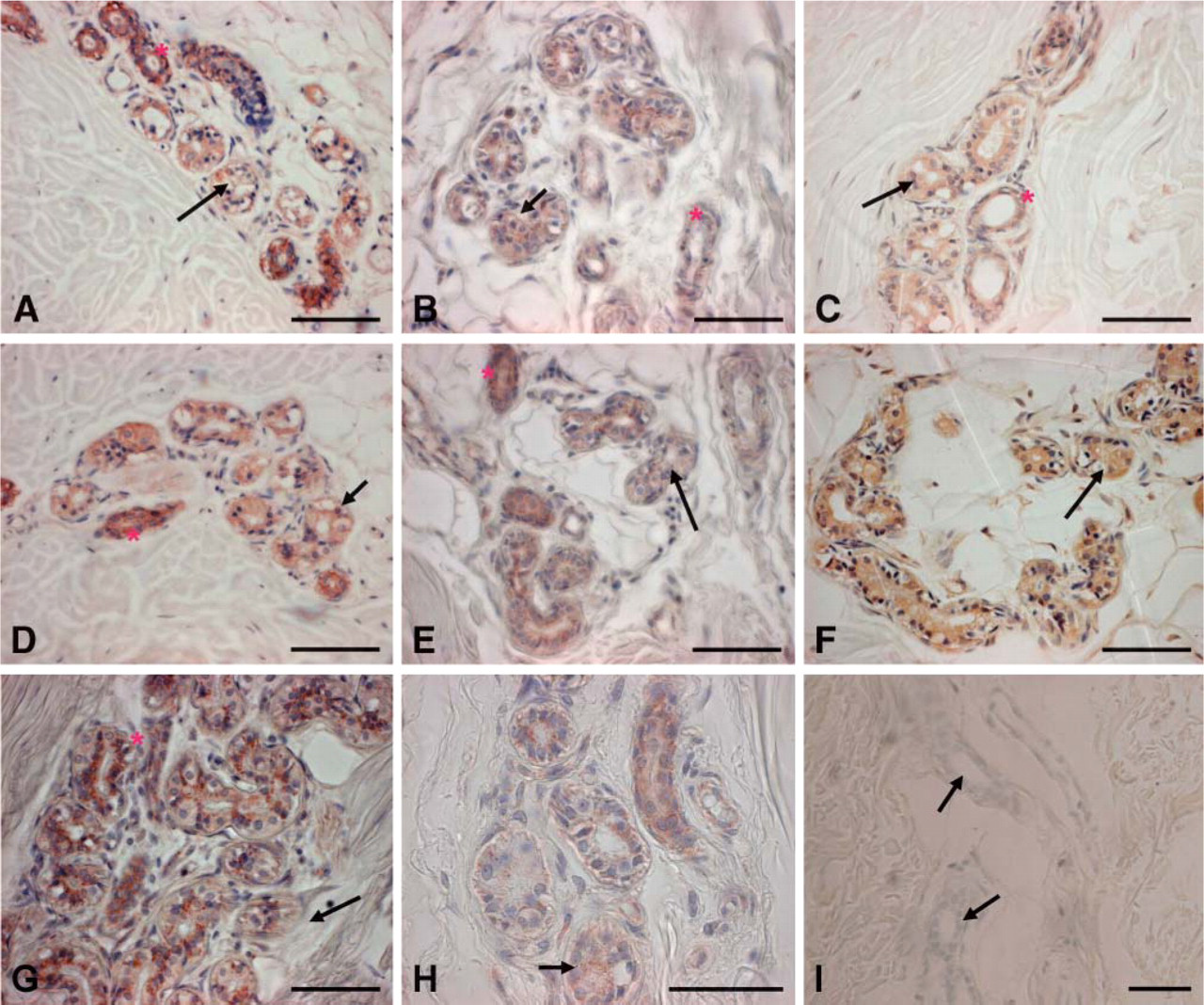
Immunohistochemical (IHC) staining of human eccrine sweat glands for the SIBLINGs and their known MMP partners. Positive reactions are denoted by brownish-red coloration of the gland tissue. Positive immunoreactivity to osteopontin [LFMb-14 monoclonal antibody (MAb); see Table 1] is present (A) in both the coiled (arrow) and straight ductular portion (asterisk) of the gland, although the intensity of staining is greater in the straight portions. Immunoreactivity for bone sialoprotein (B) LFMb-25 MAb (see Table 1), dentin matrix protein-1 (
In contrast, no immunoreactivity in the acini and duct systems of the lacrimal gland was observed for any of the five SIBLINGs or their known MMP partners (representative results, Figure 3A for OPN). Immunoreactivity for mitochondria antibody within the lacrimal gland tissue was low to negative (Figure 3B, also right half of Figure 3C) but positive within the adjoining muscular structure (Figure 3C, left half), verifying the relatively low metabolic potential of the lacrimal gland ductal epithelial cells. To validate the quality of lacrimal tissues for IHC, immunoreactivity for aquaporin 5 (AQP5), known to be highly expressed in the acini of lacrimal glands (Ohashi et al. 2003), was positive (Figure 3D) when carried out simultaneously with the survey for SIBLINGs and MMPs. Positive control with human kidney sections exhibited intense immunoreactivity for the SIBLINGs and MMPs in their respective nephron locations, as shown in the representative result for MEPE in the proximal tubules (Figure 3E) as previously reported (Ogbureke and Fisher 2005). The negative control consisted of treatment with preimmune rabbit IgG for lacrimal gland (Figure 3F; representative lacrimal gland stain with preimmune rabbit IgG).
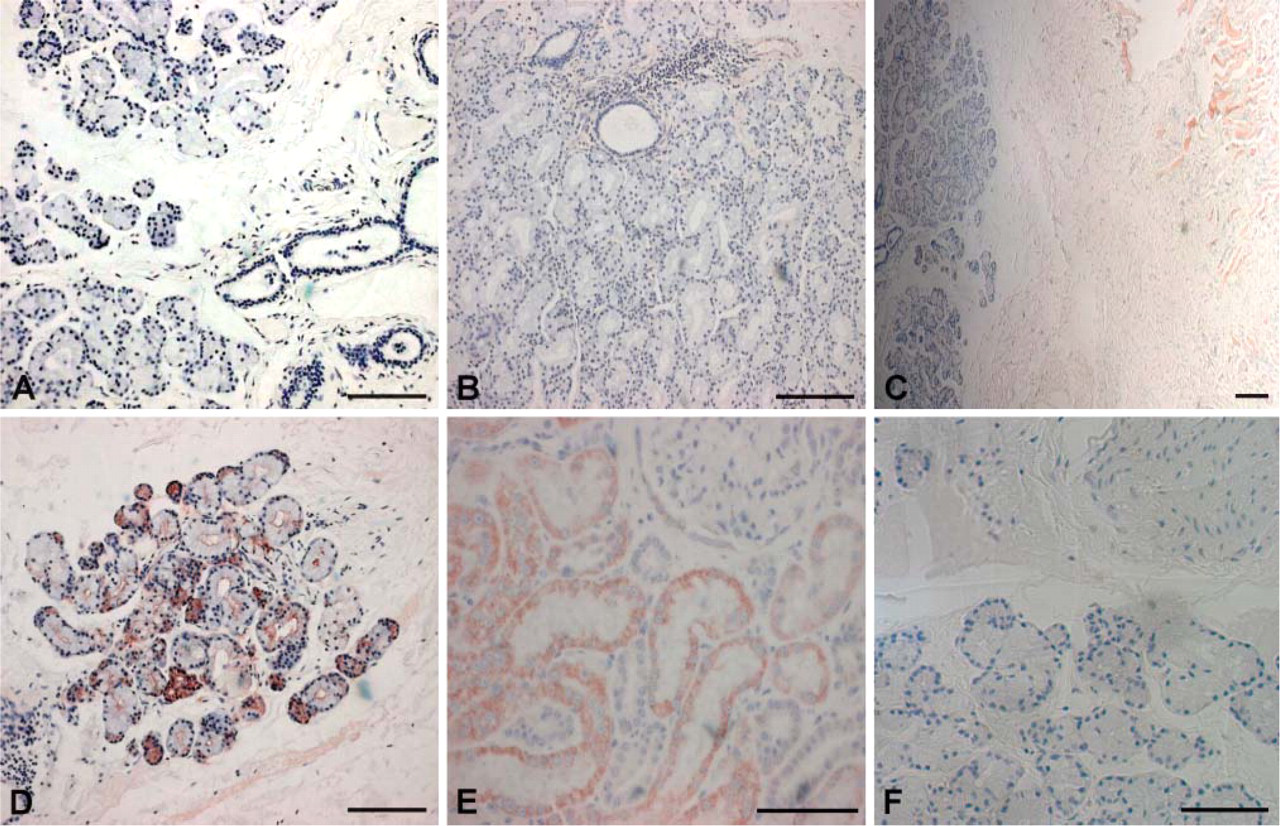
IHC staining of monkey lacrimal glands for the SIBLINGs and their known MMP partners. Positive reactions are represented by brownish-red coloration. Unlike the eccrine sweat glands, the lacrimal acini and ducts lacked immunoreactivity for all five SIBLINGs and their known MMP partners. (
Discussion
Individual members of the SIBLING family of proteins were first discovered in bones and teeth. These tissues are known to have very high metabolic rates due to the synthesis and biomineralization of large amounts of matrix. SIBLING expression outside the mineralized tissues was next described in a variety of epithelial tumors including those of breast, prostate, thyroid, and lung (Bellahcene et al. 1994; Bellahcene and Castronovo 1995; Bellahcene et al. 1996; Chambers et al. 1996; Waltregny et al. 1998), all cancers also known to have high rates of metabolism. A recent report also documented the temporal and transient expression of the OPN, BSP, and DMP1 in the cytoplasm of Sertoli cells of developing gonads of normal male mice. Although the authors proffered no distinct function for BSP and DMP1, they suggested that OPN may be involved in regulating migration, cell process formation, or adhesion during testis development (Wilson et al. 2005).
Expression of SIBLINGs and their partner MMPs in the salivary gland and kidney duct systems led to the hypothesis that these proteins may be expressed in other metabolically active ductal epithelia. To test this hypothesis, we chose to look for the SIBLINGs and their MMP partners (when known) in two other relatively simple epithelial ductal systems: one with a high metabolism potential, the other with a relatively low metabolism.
The sweat gland, with its high requirement for energy for the rapid recovery of salts from the initial isotonic fluid, was hypothesized to express some or all of the SIBLING/MMP complexes, whereas the lacrimal gland duct with its relatively low requirement for energy for essentially passively conducting the isotonic fluid to the surface should exhibit low or complete lack of SIBLING/MMP expression. The findings indicate that the high energy-requiring sweat gland ducts do express four acidic SIBLINGs, OPN, BSP, DSPP, and DMP1, along with their known MMP partners (MMP-2, MMP-3, and MMP-9), whereas the lacrimal gland expressed none. There was no expression of MEPE in either of the tissues. It is logical that the high metabolic activities of the ductal cells of the sweat and salivary glands and the tubular cells of the nephron result in damage to the cell surface and pericellular and/or local basement membrane proteins by oxidative byproducts. Because these epithelial cells possess significant longevity (and are not thought to be aided by any helper cells), it is reasonable to hypothesize that they must have their own mechanisms to digest and remove proteins damaged by free radicals and other oxidative byproducts.
The structurally similar but more passive and low-energy-requiring lacrimal gland ductal system lacked SIBLING and MMP expression but was positive for the control protein, AQP5 (Ohashi et al. 2003). The tonicity of initial tear fluid produced by the lacrimal gland acini is similar to that of initial saliva at the secretory end-piece, both being isotonic. However, the tonicity of the final tear fluid from the tear duct remains unmodified (Pflugfelder et al. 2000; Wu et al. 2006) in contrast to the final products of saliva or urine, which are hypotonic (Ferguson 1988). This casts the lacrimal duct as a passive conduit for essentially fully processed tears passing along the lumen for deposition on the surface of the eyes. This apparently passive process may account for the low-energy activity of the functional lacrimal gland ducts and suggests a relatively low rate of damage to its cell surface and pericellular and basement membrane proteins.
Lack of MEPE expression in both the sweat glands and the lacrimal glands provides additional evidence that MEPE function in duct epithelia may be different from that of the other SIBLING family members. MEPE was first found associated with small tumors that caused phosphate wasting (hypophosphatemic osteomalacia) and was hypothesized to be a phosphate-controlling hormone (Rowe et al. 2000). Later, FGF23 was shown to be the major phosphatonin in that disease (Strewler 2001; Silve and Beck 2002; Weber et al. 2003; Schiavi and Kumar 2004), although the role of MEPE and phosphate transport in that type of tumor has not been clarified. In the kidney, MEPE expression is limited to the proximal tubule elements, particularly the luminal surfaces (Ogbureke and Fisher 2005). This is the major site for active phosphate transport in the kidney (Berndt et al. 2005). The level of phosphate ions in unstimulated whole saliva is substantially higher than that found in the blood (Shannon et al. 1962; Macgregor and Edgar 1986; Ferguson 1988; Agha-Hosseini et al. 2006). This appears to be due to active ion transport (Delzer and Meyer 1984), and MEPE has previously been shown to be expressed also in the salivary gland (Ogbureke and Fisher 2004). Unlike the kidney and salivary gland where phosphate is actively transported, neither the sweat gland nor the lacrimal gland is thought to do so. This continues to suggest that MEPE expression is associated with cells involved in active phosphate transport against a concentration gradient (Rowe et al. 2000; Rowe 2004).
In summary, SIBLING/MMP profile of the lacrimal gland has provided a significant comparison to that of the sweat gland, as well as that of the salivary gland and kidney previously reported (Ogbureke and Fisher 2004,2005). Cells that express the acidic SIBLINGs and their partner MMPs appear to be metabolically active, and these data suggest the hypothesis that the SIBLING/MMP complexes may be involved in the turnover of extracellular proteins damaged by oxidation byproducts. On the other hand, MEPE expression appears to be associated more closely with active phosphate ion transport.
Footnotes
Acknowledgements
This research was supported in part by the Intramural Research Program of the National Institute of Dental and Craniofacial Research (NIDCR), National Institutes of Health, Bethesda, MD.
We thank Ms. Li Li (Craniofacial and Skeletal Disease Branch, NIDCR) for assistance with histological tissue preparation and immunohistochemistry.