
Abstract
The sarcoglycan complex (SGC) is a multimember transmembrane complex interacting with other members of the dystrophin–glycoprotein complex (DGC) to provide a mechanosignaling connection from the cytoskeleton to the extracellular matrix. The SGC consists of four proteins (α, β, γ, and δ). A fifth sarcoglycan subunit, ∊-sarcoglycan, shows a wider tissue distribution. Recently, a novel sarcoglycan, the ζ-sarcoglycan, has been identified. All reports about the structure of SGC showed a common assumption of a tetrameric arrangement of sarcoglycans. Addressing this issue, our immunofluorescence and molecular results showed, for the first time, that all sarcoglycans are always detectable in all observed samples. Therefore, one intriguing possibility is the existence of a pentameric or hexameric complex considering ζ-sarcoglycan of SGC, which could present a higher or lower expression of a single sarcoglycan in conformity with muscle type—skeletal, cardiac, or smooth—or also in conformity with the origin of smooth muscle.
T
The SGC consists of four transmembrane proteins: α-sarcoglycan, a type-I transmembrane protein with N terminus on the intracellular face of muscle cells specifically expressed in skeletal and cardiac muscle, even though low amounts are also detected in bladder, lung, and small intestine (Roberds et al. 1993, 1994); β-sarcoglycan, a type-II transmembrane protein most abundant in cardiac and skeletal muscle but also expressed in placenta, kidney, liver, and lung (Lim et al. 1995; Bönnemann et al. 1996); and γ- and ζ-sarcoglycans, also type-II glycosylated transmembrane proteins that are highly similar among themselves and similar to β-sarcoglycan (Yoshida et al. 1994; Jung et al. 1996).
Whereas δ-sarcoglycan is detected in all types of muscle, γ-sarcoglycan is expressed exclusively in striated muscle (Noguchi et al. 1995). Attempts to immunolocalize γ-sarcoglycan in smooth muscle have failed, and the question of whether γ-sarcoglycan or a smooth muscle isoform existed was unanswered, although previous reports anticipated the presence of γ-sarcoglycan in smooth muscle (Durbeej and Campbell 1999; Durbeej et al. 2000).
The synthesis of all four sarcoglycans is required to ensure proper localization of the complex to the cell surface membrane (Holt and Campbell 1998); thus, mutations in the genes encoding individual sarcoglycans often lead to the concomitant loss or reduction of all sarcoglycans from the sarcolemma. This suggests that complex formation and localization of SGC require all four subunits (Straub and Campbell 1997) because direct interaction among the sarcoglycans has been demonstrated biochemically by coimmunoprecipitation (Yoshida et al. 1994; Chan et al. 1998).
A fifth sarcoglycan subunit, ∊-sarcoglycan, is more broadly expressed, showing a wider tissue distribution (Ettinger et al. 1997; McNally et al. 1998). The ∊-sarcoglycan, homologous to α-sarcoglycan, is a single-pass type-I transmembrane protein with a short cytoplasmic tail containing three putative protein kinase phosphorylation sites, whereas the extracellular domain contains one N-linked glycosylation site and five cysteine residues (Betto et al. 1999).
In previous reports it has been demonstrated that ∊-sarcoglycan associates with β- and δ-sarcoglycans in smooth muscle (Durbeej and Campbell 1999). Despite its homology to α-sarcoglycan and its presence in skeletal muscle, endogenous ∊-sarcoglycan is unable to rescue phenotypes associated with α-sarcoglycan loss (Duclos et al. 1998). ∊-Sarcoglycan immunoreactivity, like β- and δ-sarcoglycans, was also widely distributed in non-muscle tissues (Ettinger et al. 1997). Recent observations demonstrated that, in lung, this glycoprotein was associated with both alveoli and bronchioles and that the sections of urogenital and digestive tracts were ∊-sarcoglycan positive (Ettinger et al. 1997).
Recently, a novel mammalian sarcoglycan, the ζ-sarcoglycan, has been identified by an antibody specific to ζ-sarcoglycan (Wheeler et al. 2002). This protein, encoded by a gene on human chromosome 8p22, is a protein highly related to γ- and δ-sarcoglycans and is reduced at membrane in muscular dystrophy, consistent with a role in mediating membrane stability. ζ-Sarcoglycan was also found as a component of the vascular smooth muscle sarcoglycan complex and presented a costameric pattern (Wheeler et al. 2002).
The key role of all sarcoglycans was demonstrated with a model for the assembly, processing, and membrane stability of SGC (Hack et al. 2000). In particular, δ-sarcoglycan is critical for the initial stability of the sarcoglycan complex in the endoplasmic reticulum, with the β-sarcoglycan. After formation of the β/δ sarcoglycan core, the presence of γ-sarcoglycan is required later for efficient complex assembly and maturation. Finally, an interaction with dystrophin late in the secretory pathway is required for the sarcoglycan subcomplex to insert in the cell surface (Hack et al. 2000).
Previous investigations have demonstrated that in skeletal and cardiac muscle, SGC is a heterotetrameric unit constituted by the α-, β-, γ-, and δ-sarcoglycans (Holt and Campbell 1998). Other authors confirmed that expression of α-sarcoglycan is restricted to striated muscle cells (Roberds et al. 1993), whereas ∊-sarcoglycan is also expressed in several other tissues (Ettinger et al. 1997; McNally et al. 1998). On this basis, it was hypothesized that, in skeletal muscle, ∊-sarcoglycan interacts with β-, γ-, and δ-sarcoglycans, constituting a second SGC that coexists with the conventional subcomplex (Liu and Engvall 1999), whereas in smooth muscle—vascular and visceral—it associates with β- and δ-sarcoglycans and sarcospan (Durbeej and Campbell 1999; Straub et al. 1999).
Moreover, further analysis showed the presence of another sarcoglycan subcomplex in vascular and visceral smooth muscle consisting of ∊-, β-, γ-, and δ-sarcoglycans and is associated with sarcospan (Barresi et al. 2000). Therefore, it is hypothesized that ∊-sarcoglycan might replace the α-sarcoglycan in smooth muscle to form a unique SGC (Straub et al. 1999).
Successive reports suggest that two sarcoglycan subcomplexes exist. One containing α-, β-, γ-, and δ-sarcoglycans, characteristic of skeletal and cardiac muscle, and the other consisting of β-, δ-, ζ-, and ∊-sarcoglycans, characteristic of vascular smooth muscle (Straub et al. 1999; Wheeler et al. 2002).
The starting point of our study was the recent evidence that these disagreeing hypotheses about the composition of SGC present only a common assumption of a tetrameric arrangement of sarcoglycans (Liu and Engvall 1999).
On this basis, in our recent immunohistochemical and molecular investigations carried out on surgical biopsies of human adult visceral smooth muscle, we showed that these sarcoglycans all coexist in the same fiber. Based on these findings, we hypothesized the presence of a pentameric structure of SGC and not a conventional heterotetrameric unit (Anastasi et al. 2005).
Addressing this issue, in this work we extend our previous studies, performing immunofluorescence and molecular investigations to better verify whether this tetrameric structure also exists in all other regions that contain smooth muscle fibers. In particular, we performed an immunofluorescence and molecular analysis using samples of normal human smooth muscle obtained from the gastrointestinal, urogenital, vascular, and respiratory tracts.
Materials and Methods
Samples of normal human smooth muscle were obtained from 10 male patients who underwent surgery but who were not affected by any neuromuscular pathology. Patients were between 30 and 60 years of age. We obtained biopsies from all regions of the gastrointestinal tract (stomach, duodenum, jejunum, ileum and cecum), urogenital tract (bladder, ureter, and uterus), bronchioles, and saphena. All patients gave informed consent and all procedures followed were in accordance with the Helsinki Declaration of 1975.
Collected biopsies were treated for analysis by immunofluorescence and RT-PCR techniques, respectively.
Immunohistochemistry
Biopsies were fixed in 3% paraformaldehyde in a 0.2 M phosphate buffer, pH 7.4, for 2 hr. After numerous rinses in 0.2 M phosphate buffer and PBS, the biopsies were infiltrated with saccharose at 12% and 18% to obtain a gradual substitution of saline solution with glucosate solution and then to avoid disruption of cellular membranes during successive phases. Finally, sections were frozen in liquid nitrogen.
Twenty-μm-thick sections were cut on a cryostat and collected on glass slides coated with 0.5% gelatin and 0.005% chromium potassium sulfate. To block nonspecific sites and to render the membranes permeable, sections were preincubated with 1% BSA and 0.3% Triton X-100 in PBS at room temperature for 15 min. Finally, sections were incubated with primary antibodies for 2 hr.
The following primary antibodies obtained from Novocastra Laboratories (Newcastle upon Tyne, UK) were used: mouse monoclonal anti-α-sarcoglycan (diluted 1:100), mouse monoclonal anti-β-sarcoglycan (diluted 1:200), mouse monoclonal anti-γ-sarcoglycan (diluted 1:100), mouse monoclonal anti-δ-sarcoglycan (diluted 1:50), and mouse monoclonal anti-∊-sarcoglycan (diluted 1:100).
In all reactions, TRITC-conjugated IgG anti-mouse in goat was used as the first fluorochrome (1:100 dilution; Jackson ImmunoResearch Laboratories, West Grove, Pa) and applied for 1 hr after incubation with the primary antibody.
For double-localization reactions, after many rinses with PBS and incubation with a biotinylated IgG in goat to obtain saturation of residual free binding sites, sections were incubated with a second antibody conjugated with FITC-conjugated secondary IgG as second fluorochrome (1:100 dilution; Jackson ImmunoResearch Laboratories).
Slides were finally washed in PBS and sealed with mounting medium. Sections were then observed and photographed using a Zeiss LSM 510 confocal microscope (Carl Zeiss; Jena, Germany), equipped with an argon laser (458, 488 λ) and two HeNe lasers (543 and 633 λ). All images were digitalized at a resolution of 8 bits into an array of 2048 × 2048 pixels. Optical sections of fluorescence specimens were obtained using HeNe laser (543 nm) and argon laser (458 nm) at a 1-min, 2-sec scanning speed with up to eight averages; 1.50-μm-thick sections were obtained using a pinhole of 250. For each reaction, at least 100 fibers were observed to obtain a statistical analysis.
Contrast and brightness were established by examining the most brightly labeled pixels and choosing settings that allowed clear visualization of structural details while keeping the highest pixel intensities close to 200. The same settings were used for all images obtained from the other samples that had been processed in parallel.
The function called “display profile” allowed us to show the intensity profile across the image along a freely select-able line. Intensity curves are shown in a graph below the scanned image.
Digital images were cropped and figure montages prepared using Adobe Photoshop 5.0 (Adobe Systems; Palo Alto, CA).
RT-PCR
In the present study we collected tissue samples of human smooth muscle of saphena, bladder, ureter, uterus, bronchioles, and gastrointestinal tract. We evaluated the expression of α-, β-, γ-, δ-, ∊-, and ζ-sarcoglycans in each of them by RT-PCR.
Total RNA Isolation
Fifty to 100 mg of each tissue sample was homogenized using a power homogenizer (Ultra Turrax; IKA–Werke GmbH, Staufen, Germany). Total RNA was isolated by procedures based on monophasic solutions of phenol and guanidine isothiocyanate upon single-step RNA isolation (TRIzol Reagents; Invitrogen, Carlsbad, CA) (Chomczynski and Sacchi 1987).
RT-PCR Analysis
RT-PCR procedure was carried out using the two-step protocol (GeneAmp Gold RNA PCR Reagent Core Kit; Applied Biosystems, Foster City, CA) in a thermal cycler (GeneAmp PCR System 9600 PE; Applied Biosystems).
In the first step, an initial reverse transcription reaction (RT) was carried out in a 20-μl volume containing 3 μg of total RNA, 10 U RNase inhibitor, DTT 10 mM, 15 U Multiscribe Reverse Transcriptase and oligod (T)16 1.25 μM under the following thermal cycler conditions: hold 10 min at 25C and 12 min at 42C.
In the second step, a further independent PCR was carried out in a 50-μl volume containing 5 μl of cDNA of the first step (RT) as template, 2.5 U AmplTaq Gold DNA Polymerase, and 0.2 μM of each primer designed by us on mRNA sequences (Table 1).
DNA amplification was performed conventionally; each sample together with an internal control was subjected to 30 cycles of amplification (exponential phase of amplification) consisting of 30 sec of denaturation, 30 sec of annealing, and 40 sec of extension. The final extension step at 72C was extended to 7 min. The annealing temperature was optimized for each primer set.
For each component of the SGC, human GAPDH cDNA as internal control was used. The sequence of sarcoglycans was later confirmed by nucleotide sequencing analysis.
Oligonucleotide primer sequences used for genotyping

Nucleotide Sequencing Analysis
Amplified DNA was purified using a commercially available kit (GFX PCR DNA and Gel Band Purification Kit; Amersham Biosciences, Piscataway, NJ). The fragments extracted were directly sequenced with the primers used for the RT-PCR assay and labeled with the ABI PRISM Big Dye Terminator Cycle Sequencing Method, according to manufacturer's instructions, on 377 ABI PRISM Sequencer Analyzer (Applied Biosystems). ABI Sequencing Analysis 3.4.1 was used to process the raw sequence data and ABI Sequencing Navigator to align sequence data.
Results
A common feature of all sarcoglycans, including ∊-sarcoglycan, is their sarcolemmal expression. Their differential distribution in muscle and non-muscle cells is well known.
To design a targeting model to better define the real structure of the SGC, we analyzed the immunofluorescence and expression of all sarcoglycans in normal adult smooth muscle obtained from all regions of the human body. In particular, we studied the gastrointestinal, urogenital, vascular, and respiratory tracts using the semiquantitative analysis by confocal laser scanning microscopy and the molecular analysis by RT-PCR.
Immunohistochemistry
Through confocal laser scanning microscopic observations, we studied the immunostaining patterns of α-, β-, γ-, δ-, and ∊-sarcoglycans using specific antibodies.
To give a membrane control, we performed a single reaction using dystrophin antibody on a stomach sample (Figure 1A). Dystrophin staining showed a normal sarcolemmal distribution. We also performed a negative control on a stomach sample, using the secondary antibody only (Figure 1B). In Figure 1C, we showed the corresponding transmitted light of Figure 1B.
Indirect immunofluorescence applied in smooth muscle fibers of the gastrointestinal tract (Figure 2 and Figure 3) revealed a relatively normal pattern of all five tested sarcoglycans.
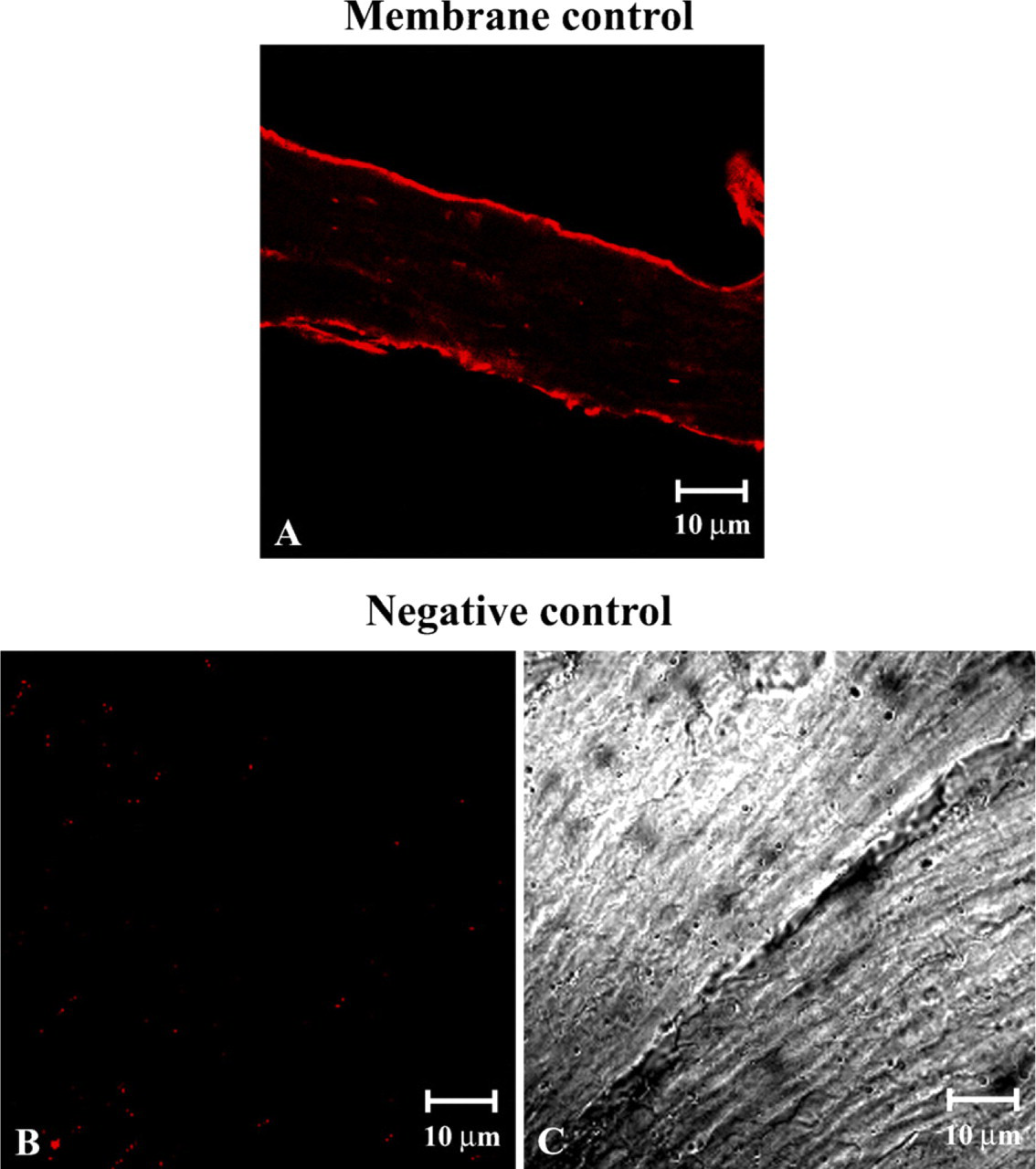
Longitudinal section of human smooth muscle immunolabeled with dystrophin antibody (
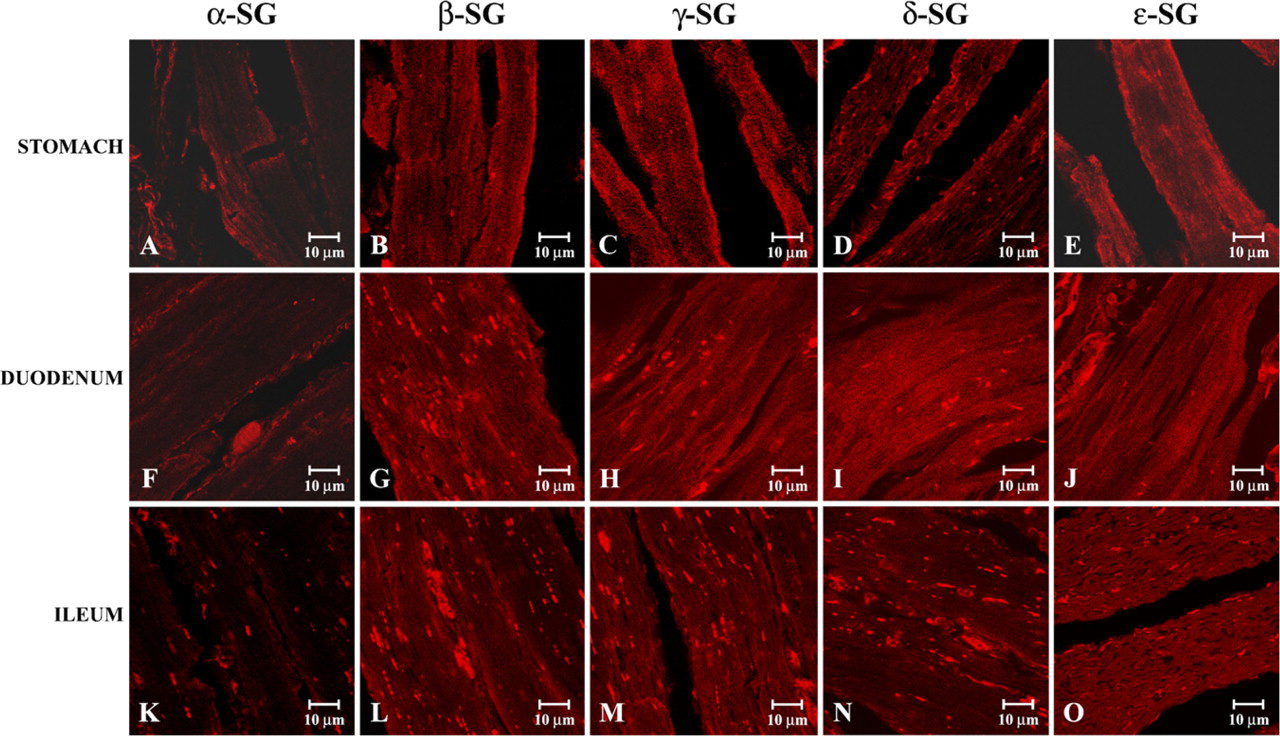
Compound panel of immunohistochemical findings in human smooth muscle of some gastrointestinal tracts. Tested proteins are indicated at the top, and the gastrointestinal tracts are indicated along the left of the figure. In the stomach (
In particular, three-dimensional reconstructions using a stack of 16 sections of 0.8 μm from the scan step showed that there was a reduced but clearly detectable staining for α-sarcoglycan in smooth muscle of stomach (Figure 2A), duodenum (Figure 2F), and ileum (Figure 2K). In the observations of other sarcoglycan stainings we showed that β-, γ-, δ-, and ∊-sarcoglycanshad a normal pattern whether in stomach (Figures 2B–2E), duodenum (Figures 2G–2J), or ileum (Figures 2L–2O).
In immunofluorescence analysis of other regions of the gastrointestinal tract (jejunum and cecum) we observed a contrary behavior of sarcoglycans. In detail, our data revealed a normal staining pattern of α-, β-, γ-, and δ-sarcoglycans, clearly visible in jejunum (Figures 3A–3D) and in cecum (Figures 3F–3I), whereas observations of ∊-sarcoglycan showed a minimally reduced staining pattern of this glycoprotein, whether in jejunum (Figure 3E) or in cecum (Figure 3J).
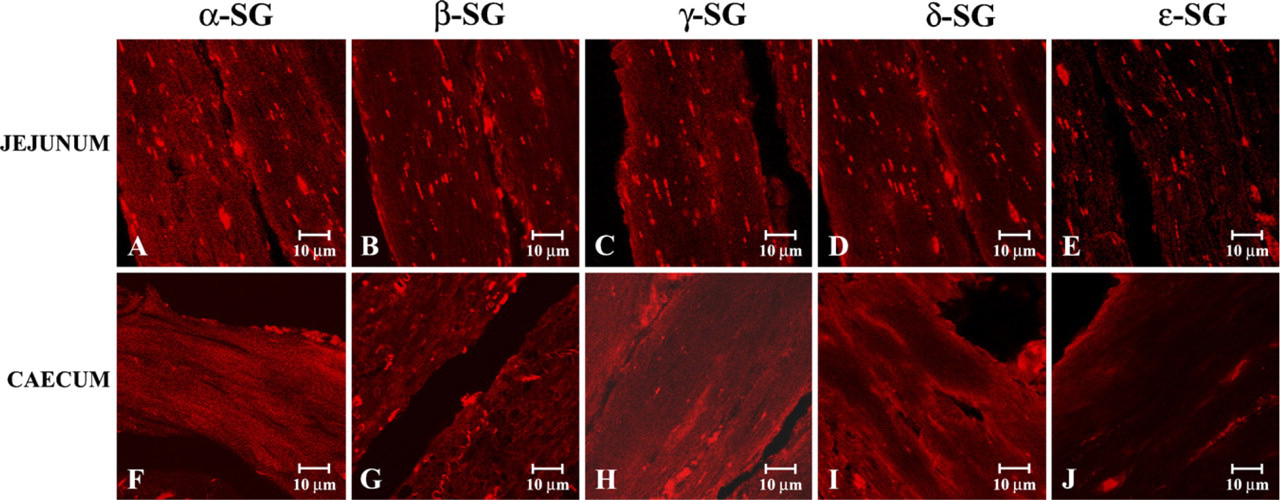
Compound panel of immunohistochemical findings in human smooth muscle of jejunum and cecum. In these images we illustrated that α-, β-, γ-, and δ-sarcoglycan staining showed a normal pattern either in jejunum (
To investigate the immunofluorescence of sarcoglycans in smooth muscle fibers of the urogenital tract, we applied the single localization reactions in smooth muscle fibers obtained from bladder, ureter, and uterus. In all observations we always detected the immunofluorescence for all sarcoglycans. In detail, in bladder we showed a normal staining pattern of α- (Figure 4A), β-(Figure 4B), γ- (Figure 4C), and δ-sarcoglycans (Figure 4D), whereas the ∊-sarcoglycan staining was normal even if minimally reduced (Figure 4E). On the contrary, in the ureter and uterus, immunofluorescence analysis showed that α-sarcoglycan staining was reduced, even if minimally (Figures 4F and 4K). The β-, γ-, δ-, and ∊-sarcoglycans staining appeared normal whether in ureter (Figures 4G–4J) or in uterus (Figures 4L–4O).
Applying the single localization reactions at smooth muscle fibers of bronchioles, we observed that all sarcoglycans showed a normal immunofluorescence staining (Figures 5A–5E). With regard to smooth muscle fibers of saphena, our data showed a reduced, but always detectable, staining pattern of ∊-sarcoglycan (Figure 5J), whereas α- (Figure 5F), β- (Figure 5G), γ- (Figure 5H), and δ-sarcoglycans (Figure 5I) showed a clearly visible and normal immunofluorescence pattern.
In addition, to better investigate the real values of all tested protein stainings, we applied the software function of “display profile” to some reactions. With this further analysis it is possible to show the intensity profile across the image along a freely selectable line converting, in this way, the immunofluorescence in a graphic. Thus, applying the display profile software to reactions in Figures 2A and 2E, we confirmed that the α-sarcoglycan fluorescence intensity in smooth muscle of stomach was detectable, even if reduced, as demonstrated by graphics that showed an intensity not overstepping limits of 50 values (Figure 6A); the display profile of ∊-sarcoglycan in smooth muscle of stomach revealed a normal immunofluorescence of these proteins showing intensity values included between 80 and 150 (Figure 6B).
The display profile applied to reactions in Figures 4A and 4E confirmed that, in the bladder, the α-sarcoglycan staining pattern was normal by intensity values that reached 200 (Figure 6C). With regard to ∊-sarcoglycan immunofluorescence, the display profile showed a clear staining pattern, even if the intensity values rarely reached 100 (Figure 6D).
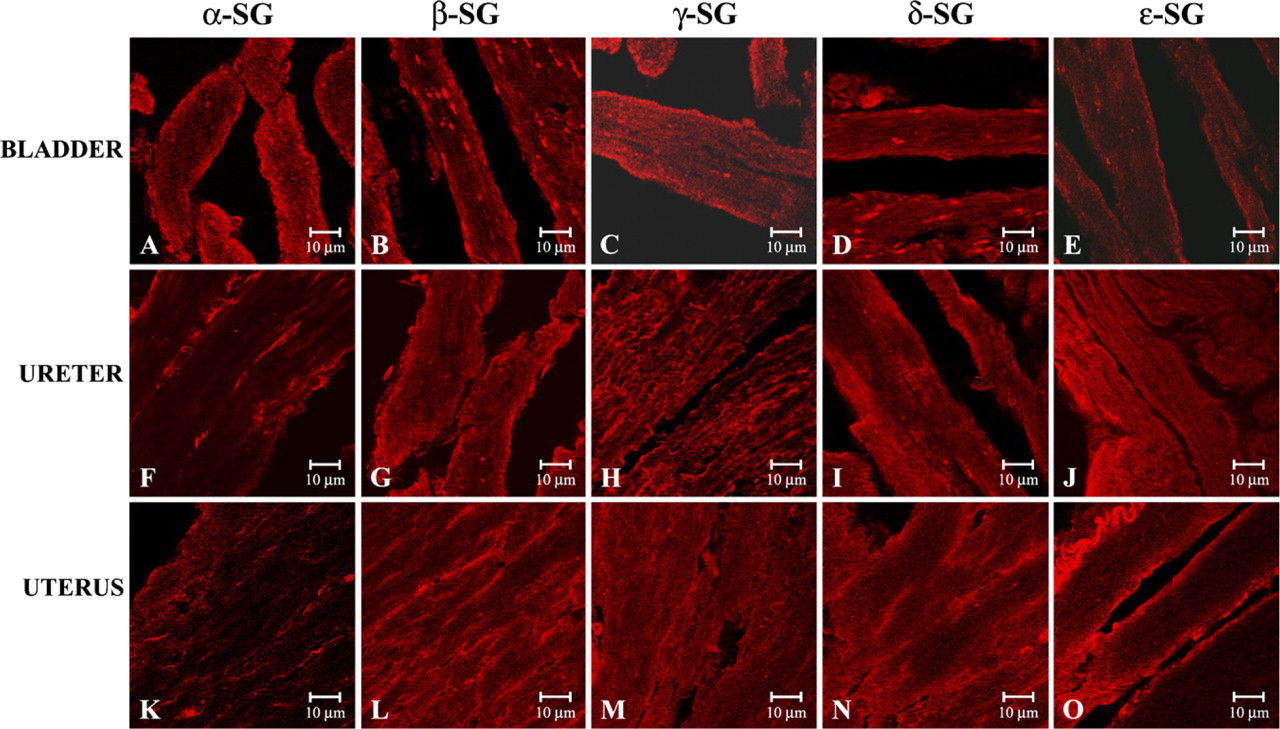
Compound panel of immunohistochemical findings in human smooth muscle of the urogenital tract. In the bladder, ∊-sarcoglycan staining (
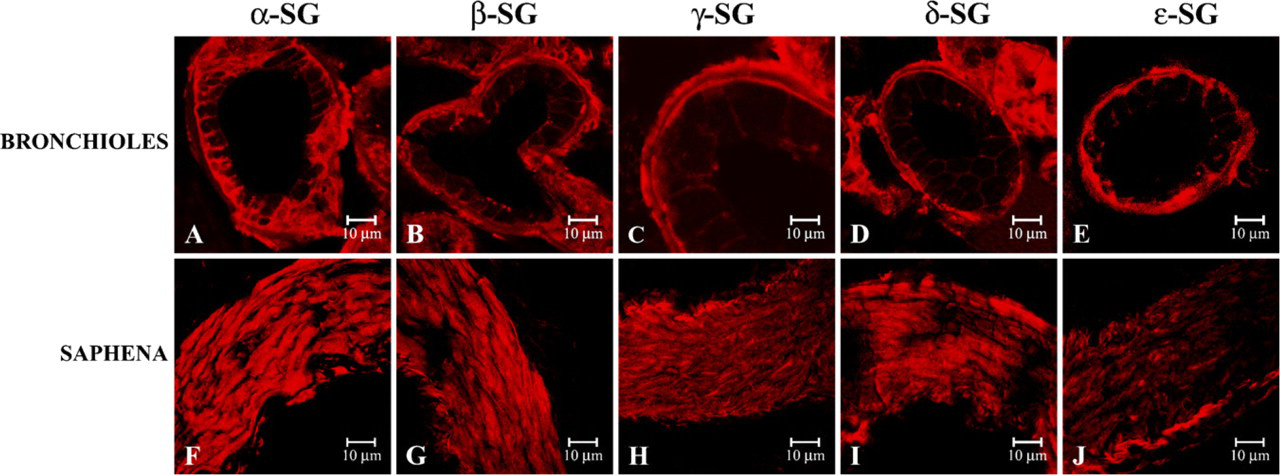
Compound panel of immunohistochemical findings in human smooth muscle of bronchioles (
Finally, the same condition was visible applying the display profile software to reactions in Figures 5F and 5J. In particular, the immunostaining patterns of α- and e∊sarcoglycans in smooth muscle of saphena was confirmed by the values included between 80 and 220 intensity around α-sarcoglycan (Figure 6E) and by values that did not overstep limits of 140 intensity (Figure 6F).
To better test the contemporary presence of all sarcoglycans and to evidence the same localization of all sarcoglycans, we performed a stock of double-localization reactions, matching antibodies to all sarcoglycans with themselves. In our observations on smooth muscle of saphena, each sarcoglycan constantly colocalizes with others. In detail, the results constantly showed a yellow fluorescence due to an overlapping of the red fluorescence of the primary channel with the green fluorescence of the secondary channel (Figure 7), indicating that sarcoglycans colocalize with each other. The same results were obtained in all other tested regions (data not shown).
RT-PCR
Using RT-PCR and nucleotide sequencing analysis with specific primers, we confirmed the presence of α-, β-, γ-, δ-, ∊-, and ζ-sarcoglycans in each sample of human smooth muscle: saphena, bladder, ureter, uterus, bronchioles, and gastrointestinal tract.
Results obtained in this work confirmed the presence of six sarcoglycans (Figure 8 and Figure 9) in all smooth muscle samples.
We have already demonstrated the presence of α-sarcoglycan in samples of human smooth muscle of several gastrointestinal tracts (Anastasi et al. 2005), and in this study we also confirmed the presence of α-sarcoglycan in human saphena, bladder, ureter, uterus, bronchioles, and gastrointestinal tract.
Discussion
We carried out a semiquantitative and molecular study on SGC using normal human samples of smooth muscle fibers. In this regard, we found the simultaneous expression of six sarcoglycans (α, β, γ, δ, ∊, and ζ), hypothesizing an exameric arrangement of SGC.
Initially, the sarcoglycans were considered as a complex of four transmembrane proteins (α, β, γ, and δ) primarily expressed in skeletal muscle (Roberds et al. 1993; Yoshida et al. 1994; Bönnemann et al. 1995) and closely associated with dystrophin and the dystroglycans in the muscle membrane (Suzuki et al. 1992; Cox et al. 1994; Greenberg et al. 1994).
Notoriously, the mutations in any sarcoglycan cause limb-girdle muscular dystrophy (Campbell 1995; Bönnemann et al. 1996). Interestingly, dysphagia, vomiting, chronic constipation, and acute digestive dilatations, all due to malfunctions of digestive smooth muscle, have been reported in patients with progressive muscular dystrophy (Barohn et al. 1988; Jaffe et al. 1990). Thus, clinical observations raise the possibility that the sarcoglycans play a key role in smooth muscle.
After these investigations, the SGC was mainly studied because the integrity of this subcomplex seems to be essential for the viability of muscle cells (Roberds et al. 1994; Tinsley et al. 1994; Campbell 1995; Lim et al. 1995; Noguchi et al. 1995). In fact, it was shown that anti-α-sarcoglycan coprecipitated integrin α5β1 and other focal adhesion-associated proteins. On this ground, a bidirectional signaling was demonstrated between sarcoglycans and integrins (Yoshida et al. 1998; Anastasi et al. 2003a, b, 2004a), also utilizing human skeletal muscle affected by sarcoglycanopathy (Anastasi et al. 2004b). Sarcoglycans thus seem to be functionally and pathologically as important as dystrophin (Yoshida et al. 1998).
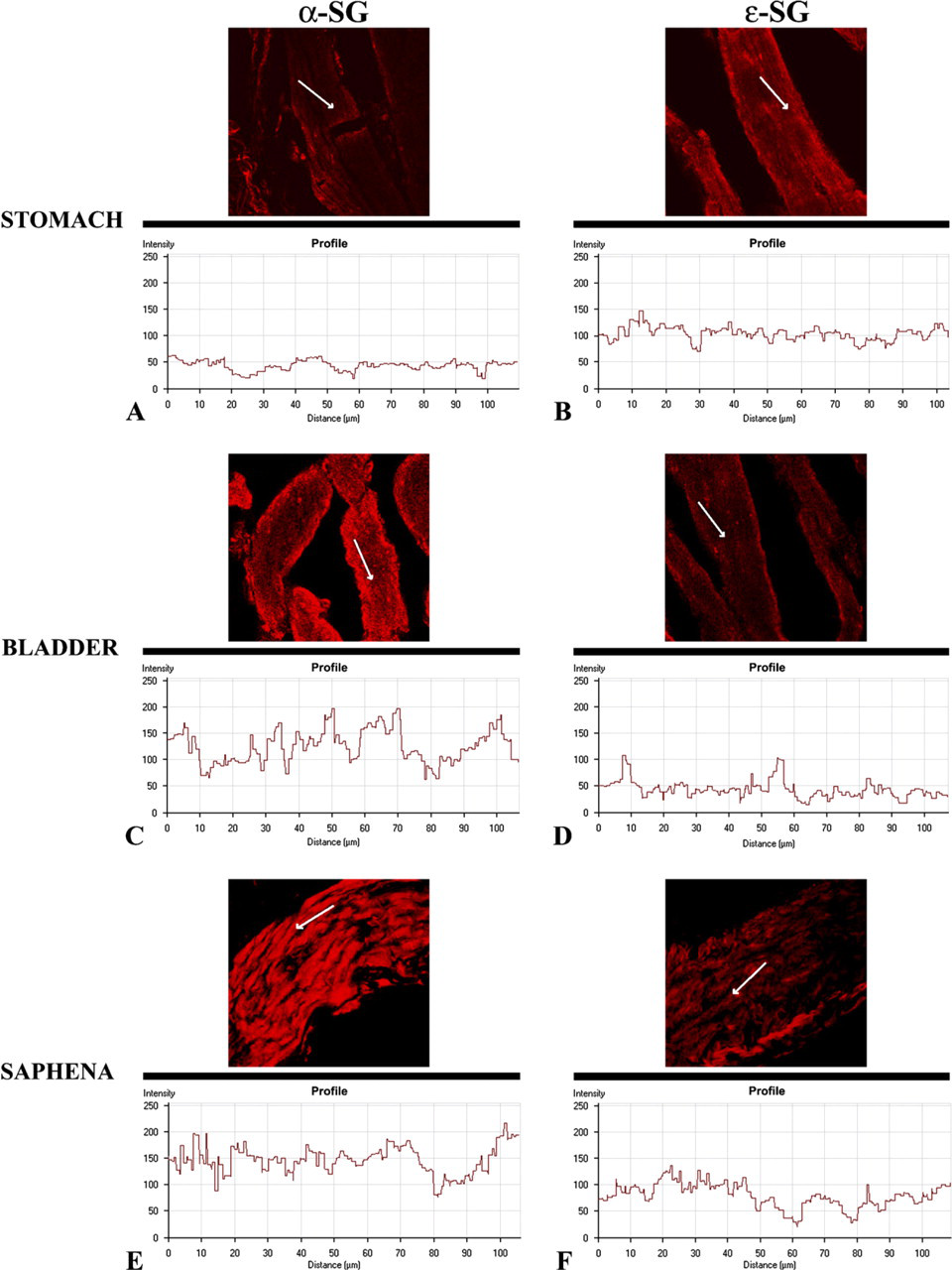
Display profiles of α- and ∊-sarcoglycans of stomach, bladder, and saphena. This further analysis shows the intensity profile across the image along a freely selectable line (arrow) of longitudinal sections of stomach, bladder, and saphena smooth muscle. In the stomach smooth muscle, α-sarcoglycan fluorescence intensity (
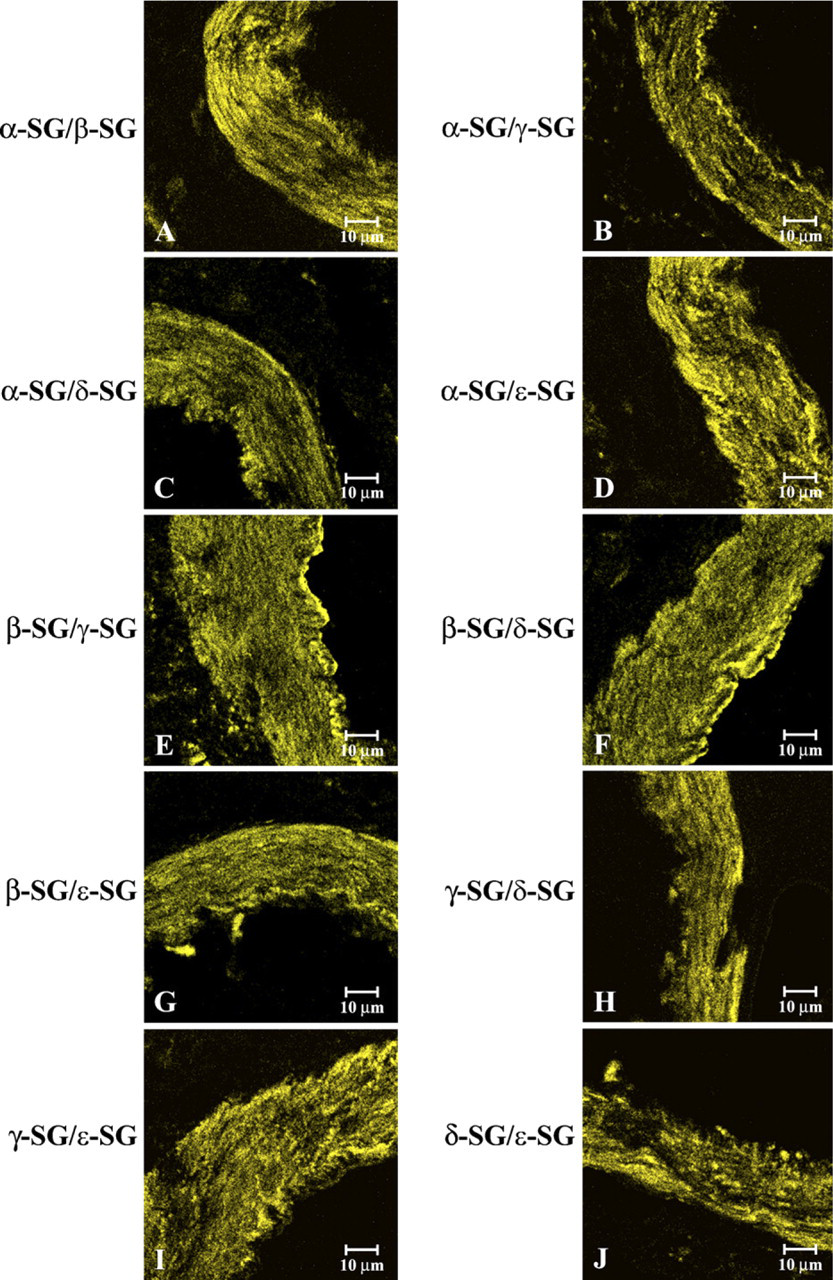
Compound panel of double-immunohistochemical reactions in human smooth muscle samples of saphena immunolabeled with α- and β-sarcoglycans (
Two additional sarcoglycan molecules were more recently described: ∊-sarcoglycan, a transmembrane glycoprotein showing 43% amino acid identity with α-sarcoglycan (Ettinger et al. 1997; McNally et al. 1998), and ζ-sarcoglycan, a protein highly related to δ-sarcoglycan and γ-sarcoglycan (Wheeler et al. 2002).
It is presently well known that the sarcoglycans include six transmembrane glycoproteins associated in different heterotetrameric units in skeletal, cardiac, and smooth muscle. In fact, it was hypothesized that ∊-sarcoglycan replaces α-sarcoglycan in smooth muscle to form a unique SGC formed by ∊-, β-, γ-, and δ-subunits (Straub et al. 1999). Further reports showed that the SGC of smooth muscle is characterized by β-, δ-, ∊-, and ζ-sarcoglycans (Wheeler and McNally 2003).
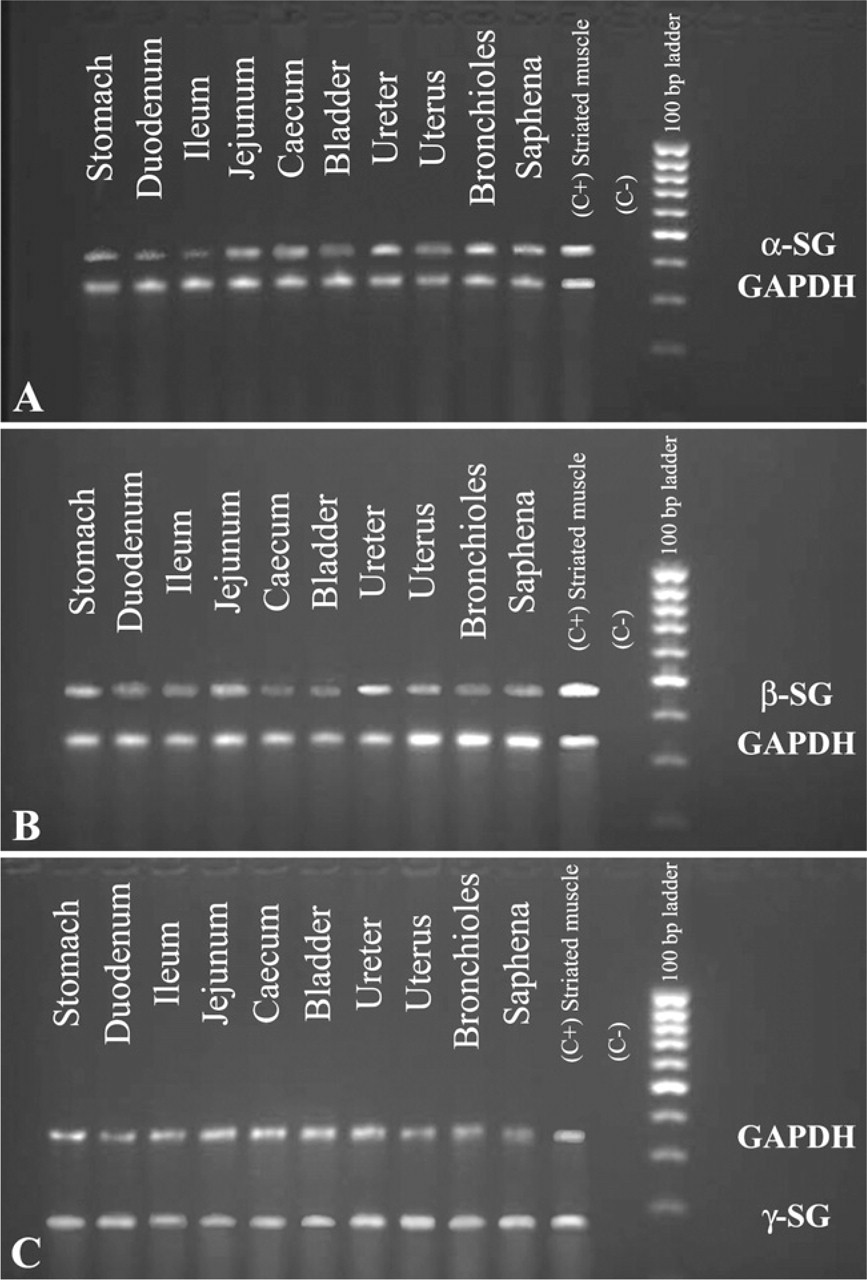
2% Agarose gel electropherogram of RT-PCR products amplified using human RNA as template, primer pairs of α-, β-, and γ-sarcoglycans, and primer pairs of GAPDH (internal control). (
A critical question is whether the SGC exists as a tetrameric or higher order structure in smooth muscle. This point was partially clarified in our previous immunohistochemical and molecular investigation, carried out on only human adult smooth muscle of gastrointestinal tract, in which we demonstrated the presence of a pentameric arrangement around the SGC (Anastasi et al. 2005).
To further investigate this question, we carried out an immunofluorescence study on α-, β-, γ- and δ- and ∊-subunits of sarcoglycans, paralleled by a molecular analysis of all six sarcoglycans including ζ-sarcoglycan. The analysis was extended to the gastrointestinal tract as well as to smooth muscle fibers of the urogenital, vascular, and respiratory tracts.
Our results showed for the first time that in all observed samples of human smooth muscle in a single localization, α-sarcoglycan is also always detectable, although its staining pattern is slightly lower than ∊-sarcoglycan, both with immunofluorescence observations and with molecular techniques. α-Sarcoglycan fluorescence was sometimes normally detected, whereas ∊-sarcoglycan staining pattern was reduced but clearly expressed. However, α- and ∊-sarcoglycans always coexist in all observed fibers. Finally, γ-sarcoglycan staining pattern, as well as β- and δ-sarcoglycans, is always normally detectable in all analyzed samples.
Previous reports anticipated the presence of γ-sarcoglycanin smooth muscle (Durbeej and Campbell 1999; Barresi et al. 2000; Durbeej et al. 2000), in contrast with data of other reports showing that this protein was not a member of the smooth muscle complex because γ-sarcoglycan was found to be expressed in striated muscle (Yamamoto et al. 1994; Barresi et al. 2000; Wheeler and McNally 2003). In particular, with the discovery of ζ-sarcoglycan, it is hypothesized that ζ-sarcoglycan, and not γ-sarcoglycan, is the fourth member of the SGC in smooth muscle, including the coronary arterial bed (Wheeler et al. 2002; Wheeler and McNally 2003). In our opinion, these hypotheses may have validity because these results were obtained using smooth muscle of mice with sarcoglycan deficiency (Wheeler and McNally 2003). Nevertheless, our results are not comparable with those of Wheeler et al. (2002) and Wheeler and McNally (2003) because we obtained our information using normal human smooth muscle. In addition, it is possible that the SGC of mice has some differences in comparison with SGC of human smooth muscle because, in the extracellular domain of sarcoglycans, substantial differences in amino acid sequences exist among different species (Betto et al. 1999).
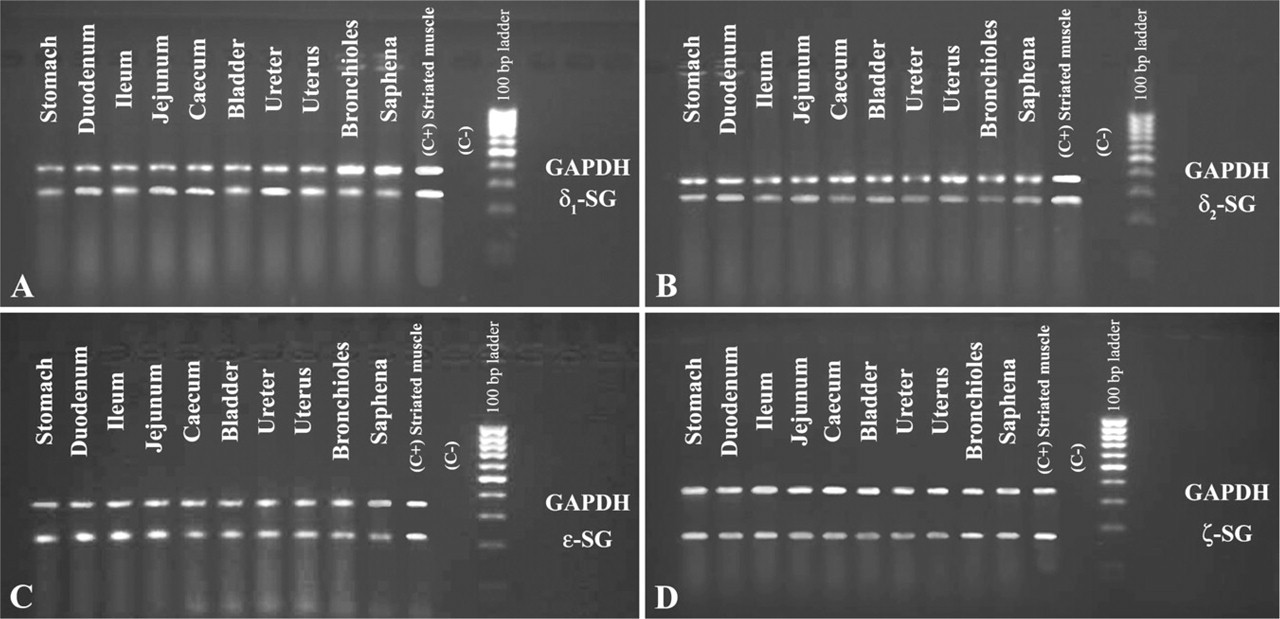
2% Agarose gel electropherogram of RT-PCR products amplified using human RNA as template, primer pairs of δ1-, δ2, ∊-, and ζ-sarcoglycans, and primer pairs of GAPDH (internal control). (
Moreover, Wheeler and McNally (2003) showed, in mice lacking γ-sarcoglycan, a focal degenerative cardiomyopathy but not a disruption of the smooth muscle SGC. Interestingly, in our opinion, these results could confirm our hypothesis of exameric structure because the function of γ-sarcoglycan in the context of SGC could be reinforced by ζ-sarcoglycan, which works together with all sarcoglycans.
Thus, our intention was to verify the interaction of γ-sarcoglycan with the other SGC components. To this end, we performed a stock of double reactions, with each sarcoglycan against all others. These data showed that all sarcoglycans colocalized with each other, confirming our pentameric or exameric hypothesis.
Our results, showing coexistence of α- and ∊-sarcoglycans in a single SGC, are in disagreement with previous studies indicating that the expressions of α- and ∊-sarcoglycans were mutually exclusive and hypothesizing that ∊-sarcoglycan serves a function similar to that of α-sarcoglycan (Liu and Engvall 1999). In particular, it is hypothesized that α- and ∊-sarcoglycans form separate complexes with β-, γ-, and δ-sarcoglycans, on wild-type and α-sarcoglycan-deficient mice and mouse C2C12 myocytes (Liu and Engvall 1999). This arrangement of two subcomplexes is valid only considering the embryological condition and origin of tested muscles.
Because myoblasts are undifferentiated cells, it is possible that these two subcomplexes change their arrangement in adult muscle condition and that all five or six sarcoglycans work together to better develop their signaling functions. Interestingly, the formation and expression of the SGC on the cell membrane begins after myotube formation (Ozawa et al. 2005). In addition, mouse α- and ∊-sarcoglycans have some homologies with human α- and ∊-sarcoglycans, but they have differences in ?10% of amino acid sequences (Ettinger et al. 1997). These differences could determine the different behavior in two types of muscles.
Finally, mouse α- and ∊-sarcoglycans are 44% identical at the amino acid level. The intracellular domain of ∊-sarcoglycan (98 aa) is thus larger than that of α-sarcoglycan (76 aa). Homology extends over the whole length of the molecules, with conserved regions both in the extracellular and transmembrane domains. The potential site for N-glycosylation in ∊-sarcoglycan corresponds to one of two such sites in α-sarcoglycan (Ettinger et al. 1997). The two sarcoglycans are similar but not identical and can thus play distinct roles in the context of the plasma membrane. In this regard, it is demonstrated that there is a pseudo-∊-sarcoglycan gene located at 2q21. In regard to α- and ∊-sarcoglycans, genes appear to have originated from a common ancestor by gene duplication (Ozawa et al. 2005), but the proteins have different roles.
In our opinion, this hypothesis could be confirmed by our results carried out onhuman adult muscle, showing a contemporary presence of all sarcoglycans both with single and with double localizations and with RT-PCR method, in comparison with other results obtained on muscle of mdx mice (Straub et al. 1999) or on mouse C2C12 cells (Liu and Engvall 1999).
Thus, our data clearly show that α- and ∊-sarcoglycans are coexpressed in different smooth muscle tissues and suggest that the two sarcoglycans could occupy two different sites in the same SGC.
Because most studies indicate the presence of four sarcoglycans as a requirement for the functionality and viability of the complex, Barresi et al. (2000) hypothesized the expectance of a yet unidentified sarcoglycan, a homolog of γ-sarcoglycan, in smooth muscle SGC. In our opinion, this unidentified sarcoglycan can be ∊-or ζ-sarcoglycan, singularly or contemporaneously. In fact, ∊-sarcoglycan is widely expressed in parts of most tissues and physically associated with other DGC components (Ettinger et al. 1997), and ζ-sarcoglycan has a high degree of similarity and homology with γ-sarcoglycan (Wheeler et al. 2002).
On this basis, our results provide the first suggestion that sarcoglycan subcomplex in smooth muscle and, potentially, also in skeletal and cardiac muscle, can consist of five or six distinct sarcoglycan subunits. Therefore, one intriguing possibility is the existence of a pentameric or, also considering ζ-sarcoglycan detected by RT-PCR, hexameric model of SGC. This hypothetical new complex formed by all sarcoglycans could present a higher or lower expression of a single sarcoglycan in conformity with muscle type—skeletal, cardiac, or smooth—or also in conformity with the origin of smooth muscle, gastrointestinal, urogenital, or respiratory tract.
The involvement of the hexameric structure of the sarcoglycan subcomplex is an intriguing possibility that may offer new approaches for the exact structural and signaling role of sarcoglycans and, consequently, for treatment of the sarcoglycanopathies.