
Abstract
A recent report provides new evidence for the presence of glucokinase (GK) in the anterior pituitary. In the present study, immunohistochemistry was used to identify the cells containing GK in the pituitary of rats and monkeys. In rats, GK was detected as a generalized cytoplasmic staining in a discrete population of cells in the anterior pituitary. In colocalization experiments, the majority of cells expressing follicle-stimulating hormone (FSH) or luteinizing hormone (LH) also contained GK. In addition to the gonadotropes, GK was observed in a subpopulation of corticotropes and thyrotropes. GK was not detected in cells expressing growth hormone or prolactin. In monkeys, GK was also observed in a discrete population of cells. Intracellular distribution differed from the rat in that GK in most cells was concentrated in a perinuclear location that appeared to be associated with the Golgi apparatus. However, similar to rats, colocalization experiments showed that the majority of cells expressing FSH or LH also contained GK. In addition to the gonadotropes, GK was observed in a subpopulation of corticotropes and thyrotropes. In the monkey, only a few cells had generalized cytoplasmic staining for GK. These experiments provide further evidence for the presence of GK in the anterior pituitary. Although some corticotropes and thyrotropes contained GK, the predominant cell type expressing GK was gonadotropes. In view of the generally accepted role of GK as a glucose sensor in a variety of cells including the insulin-producing pancreatic β-cells as the prototypical example, it is hypothesized that hormone synthesis and/or release in pituitary cells containing GK may be directly influenced by blood glucose.
Calorigenic nutrient deprivation has a dramatic impact on the endocrine system affecting the function of the majority of the endocrine glands. This is also true for cells of the pituitary as strikingly exemplified by reduction or cessation of the pulsatile luteinizing hormone (LH) and follicle-stimulating hormone (FSH) secretion from the gonadotropes, which results in infertility affecting primarily the female organism (Wade et al. 1996; Larsen et al. 2003; Wade and Jones 2004). It seems that metabolic cues, perhaps most importantly a lowering of blood glucose, trigger this “nutritional infertility.” Peripheral and central glucose sensors seem to be involved: (1) hepatic glucose sensor cells recognize the change of portal blood glucose levels and transmit this information via the vagus nerve to the nucleus tractus solitaris in the hindbrain, which then communicates with hypothalamic neurons that secrete gonadotropin-releasing hormones (GnRH); (2) the second signal arises from glucose sensors in the area postrema and also impacts on GnRH neurons. Lowering of glucose or inhibition of glucose metabolism at these sensor sites stops the GnRH oscillations and, consequently, the gonadotropin pulsations, leading in the end to cessation of estrogen and progestin production and engendering infertility. An “adipostatic hypothesis” proposes an alternative to the metabolic explanation for nutritional infertility (Ahima et al. 1996; Chan et al. 2003, 2006). It is based on the observation that starvation causes a drastic reduction of serum leptin levels and attributes the cessation of GnRH and LH oscillations to the lack of direct or indirect leptin activation of GnRH containing hypothalamic neurons. The possibility that leptin effects on gonadotropes might be glucose dependent was evaluated in studies of the estrus cycle in Syrian hamsters (Schneider and Zhou 1999; Schneider et al. 2002). It was demonstrated that intracerebroventricular leptin treatment prevented the fasting-induced anestrus in these animals, but that this beneficial effect of leptin was blocked by IP injection of 2-desoxy-
The molecular basis of cellular nutrient sensing governing regulation of fuel metabolism remains an intensively investigated and discussed topic (Prentki and Matschinsky 1987; Matschinsky 1995; Newgard and Matschinsky 2001; Matschinsky et al. 2006). Current understanding of the biochemical nature of the cellular glucose sensor is perhaps most advanced and is also highly pertinent to the present study. It is widely accepted that the glucose phosphorylating enzyme glucokinase (GK) serves a glucose sensor role in man and many laboratory animals, as most strikingly illustrated by the biochemical genetics of “glucokinase disease” (Matschinsky et al. 2006) and by the profound impact of newly discovered GK activator drugs (GKAs) on glucose homeostasis and diabetes mellitus (Grimsby et al. 2003). It is now very clear that GK functions as the glucose sensor or glucoreceptor for the insulin-secreting pancreatic β-cell, the archetypical fuel sensor of the organism, but there is also considerable evidence that GK is involved in hepatic, entero-endocrine, and hypo-thalamic glucose sensing (Matschinsky et al. 2006).
We have recently reported that the pituitary gland contains significant levels of the glucose sensor enzyme GK, as demonstrated by a new, highly specific, and sensitive spectrometric assay (Zelent et al. 2006). Even though the identity of the GK-containing cells was elusive at the time, we did speculate on the basis of this finding and in view of the generally accepted role of GK in fuel sensing that a modification of the current paradigm of anterior pituitary cell operation might be indicated, and proposed that glucose and perhaps other fuels might directly influence the production and release of hypophyseal hormones.
We report here on the identity of the GK-containing pituitary cells. We have found in both rat and monkey pituitary that GK is primarily located in the gonadotropes and also in a subpopulation of thyrotropes. On the basis of a compelling body of knowledge about the role of GK as glucose sensor in a network of cells regulating glucose homeostasis (Matschinsky et al. 2006), we interpret our findings to date as evidence that FSH/LH and perhaps thyroid-stimulating hormone (TSH) synthesis and/or release and thus reproductive function and energy metabolism might be directly controlled by blood glucose at the level of the pituitary.
Materials and Methods
Antibodies
Rabbit antibodies to the pituitary hormones were obtained from the National Hormone and Pituitary Program (Dr. A.F. Parlow; Harbor-UCLA Medical Center, Torrance, CA) of the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK; Bethesda, MD). Dilutions used for the antisera were 1:20,000 for prolactin (PRL), 1:10,000 for growth hormone (GH), 1:4000 for adrenocorticotropic hormone (ACTH), and 1:1500 for FSH, LH, and TSH. Sheep anti-GK was provided by Dr. Mark Magnuson (Vanderbilt University; Nashville, TN) and used at a final dilution of 1:800 (Jetton and Magnuson 1992). Goat anti-GK was obtained from Santa Cruz Biotechnology (Santa Cruz, CA) and used at a final concentration of 20 μg/ml IgG. Both GK antibodies detected a prominent band of 50 kDa in extracts from pancreatic islet tissue. Secondary antibodies were FITC donkey anti-rabbit (1:200), Cy3 donkey anti-sheep (1:600), and CY3 donkey anti-goat (1:600) IgGs obtained from Jackson ImmunoResearch Laboratories (West Grove, PA). The diluent was 1% BSA, 1% normal donkey serum, 0.3% Triton X-100, and 0.02% sodium azide in PBS (pH 7.3).
Tissue Processing
A total of eight Sprague Dawley rats, weighing ~125 g, were used for this study (four males and four females). Ovarian cycle status of the females was not determined. Three male and three female adult rat pituitaries were perfused transcardially with Tyrode's solution (room temperature, pH 7.2) followed by 100 ml ice-cold 4% paraformaldehyde in 0.05 M phosphate buffer. Two of these (one male and one female) were dehydrated and embedded in paraffin. Four of the pituitaries (two male and two female) were frozen and sectioned on a cryostat. Two male rat pituitaries were immersion fixed with 4% paraformaldehyde, dehydrated, and embedded in paraffin. An adult male and female monkey (Cynomologus macaque) were perfusion fixed with 4% paraformaldehyde, and their pituitaries dehydrated and embedded in paraffin. Paraffin-embedded tissue was cut into 5-μm sections, deparaffinized, and rehydrated in PBS. Perfusion fixation was done under anesthesia and tissue obtained for immersion fixation was removed within 10 min from animals terminated by exsanguination under anesthesia. All procedures were approved by the Institutional Animal Care and Use Committee at the University of Minnesota.
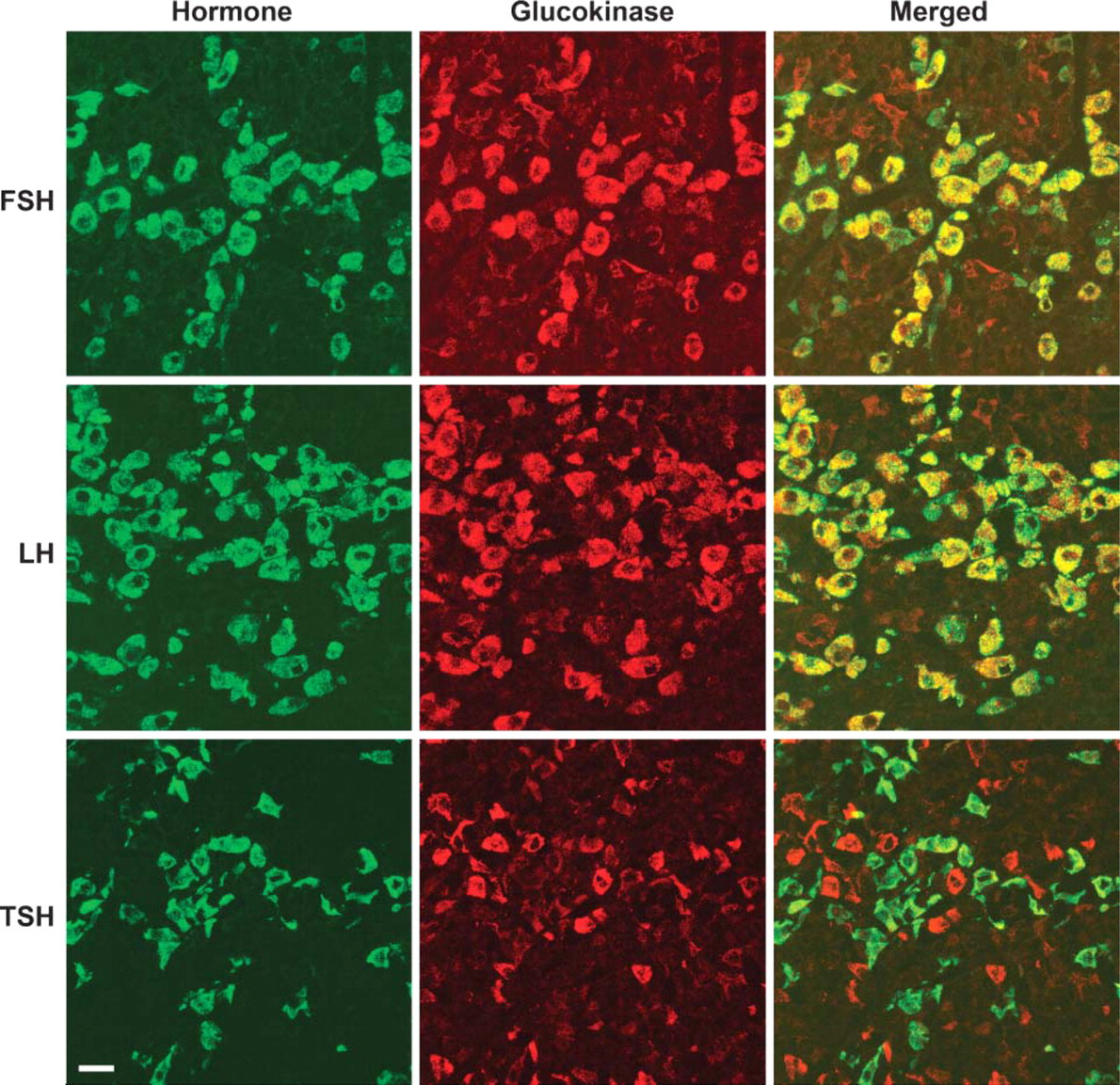
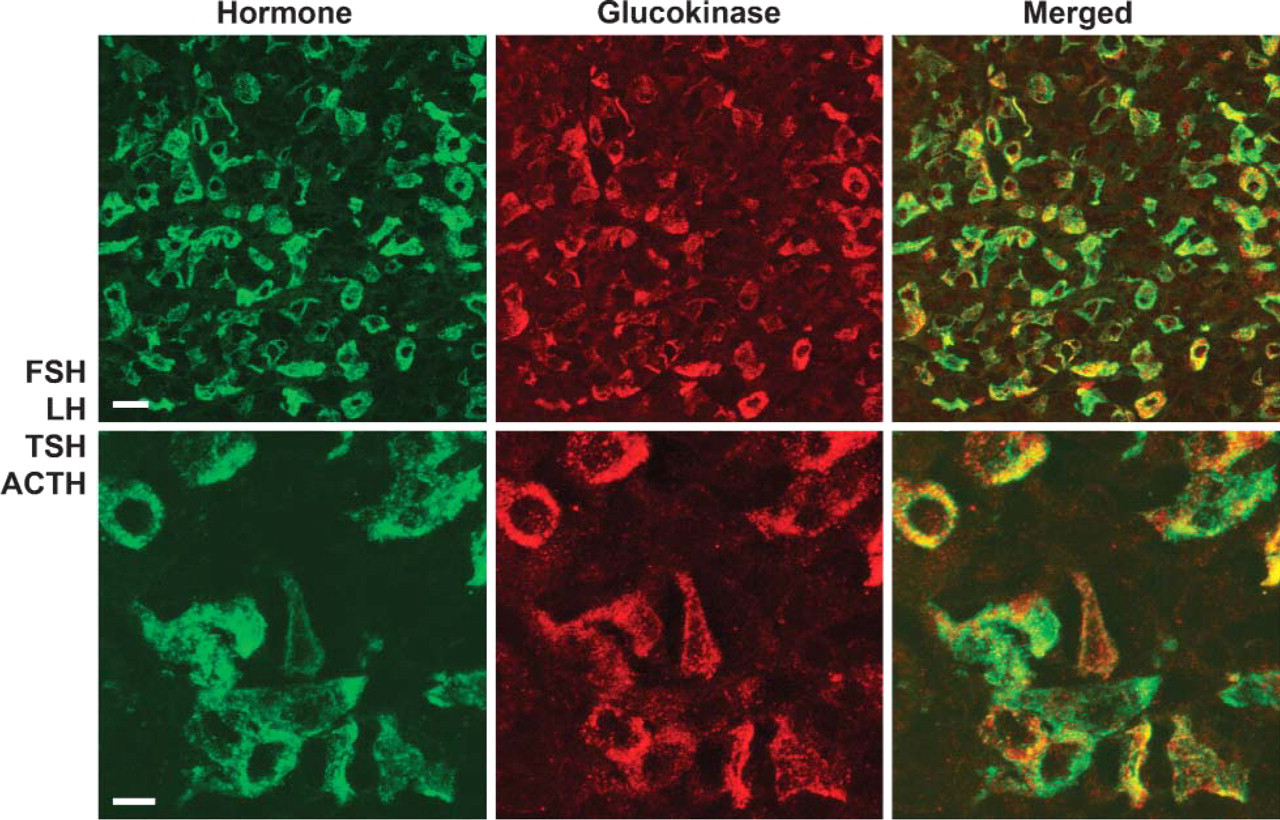
Rat pituitary double labeled with antibodies to glucokinase (GK) (red) and either follicle-stimulating hormone (FSH), luteinizing hormone (LH), or thyroid-stimulating hormone (TSH) (green). The large cells with GK immunoreactivity were gonadotropes containing FSH (top) and/or LH (middle). Most of the cells with less intense GK staining were thyrotropes containing TSH (bottom). Bar = 25 μm.
Staining Procedure
Tissue sections were incubated in the presence of the primary antibodies overnight in a humidified chamber maintained at 4C, then washed five times (10 min each) with PBS at room temperature, and incubated with the secondary antibodies at 37C in a humidified chamber for 90 min. Following five 10-min washes in PBS, the sections were mounted (85% glycerol, 3% propyl gallate in 100 mM Tris, pH 9.0) and coverslipped. For the absorption control experiments, a 1:800 dilution of the sheep anti-GK antibody was preincubated overnight with 2.5 nmol/ml GK, 10 nmol/ml glutathionyl-S-transferase (GST), or 10 nmol/ml GST-GK. A 20-μg/ml dilution of the goat anti-GK antibody was also preincubated overnight with 100 μg/ml of the immunizing peptide.
Imaging
Immunostained specimens were examined with a Bio-Rad Lasersharp 1024 Confocal Imaging System (Bio-Rad Laboratories; Hercules, CA) mounted on an Olympus microscope (AX70; Olympus, Tokyo, Japan) equipped for epifluorescence. FITC and Cy3 fluorophores were imaged using 488-nm excitation and a green bandpass emission filter (i.e., 505–540 nm) and 568-nm excitation and a red bandpass emission filter (i.e., 664–696 nm), respectively. Typically, five optical sections were acquired at 1.0-μm intervals along the z-axis through the sections using a ×60 lens (numerical aperture 1.42). Projections were then calculated from these images.
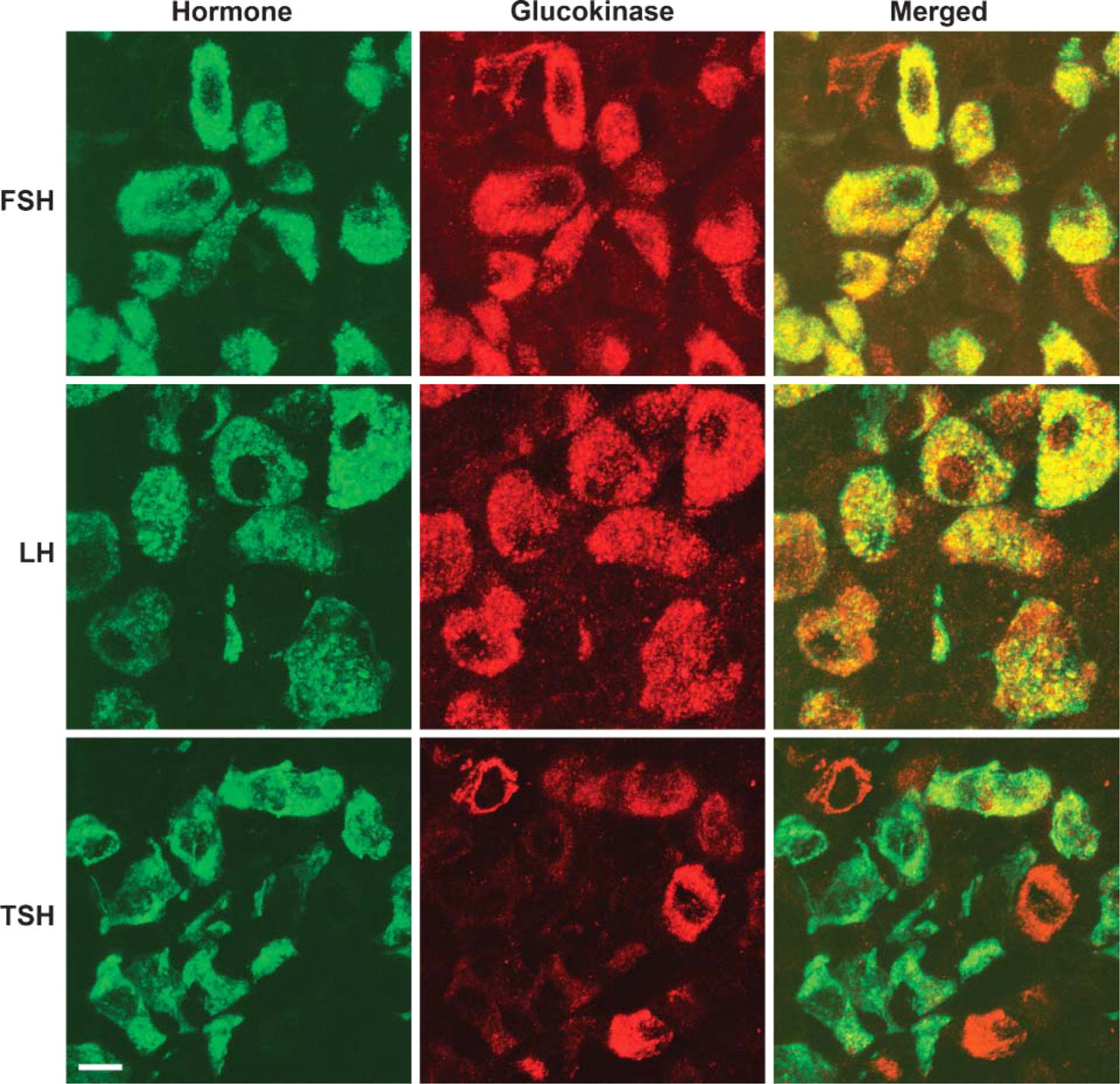
Higher magnification images from a rat pituitary double labeled with antibodies to GK (red) and either FSH, LH, or TSH (green). Staining intensity of GK in the large cells was strongly correlated with that for FSH (top). This was not true for LH where many large cells intensely stained for GK were lightly stained for LH (see cells in the lower left corner, middle). Most thyrotropes have less intense GK staining, although occasional cells have more intense staining (see cells in the upper right corner, bottom). Bar = 10 μm.
Results
In the six rat pituitaries fixed by perfusion and examined with the sheep anti-GK antibody, GK was seen as bright immunofluorescent staining of the cytoplasm. There appeared to be two populations of cells scattered throughout the pars distalis of the anterior pituitary. The most prominent of these were large round to oval cells. The second population of cells had a poorly defined angular, irregular shape. Colocalization studies were then done to determine which pituitary cells demonstrated GK immunoreactivity. FSH immunoreactivity was observed in the large cells and colocalized with GK immunoreactivity. In these cells, staining intensity for GK was strongly correlated with that for FSH. Few FSH cells were unstained for GK (Figure 1 and Figure 2). However, many GK-positive cells did not stain for FSH. LH immunoreactivity was also observed in the large cells and colocalized with GK immunoreactivity. However, there appeared to be a greater range of staining intensity for GK in the LH-positive cells than in the FSH-positive cells (Figure 1 and Figure 2). When a combination of FSH and LH antibodies were used, most of the large cells with GK immunoreactivity could be accounted for by cells expressing FSH or LH. The remaining cells with GK were mostly a second population of angular or irregularly shaped cells. Most of these cells were shown by colocalization studies to contain TSH immunoreactivity, but a small minority was ACTH positive (Figure 1-Figure 4). TSH was observed in irregularly shaped cells and some round or oval cells. GK immunoreactivity was highly variable in these cells including some with no staining for GK. ACTH stained all of the cells in the intermediate lobe and scattered irregularly shaped cells in the pars distalis. Except for an occasional cell, the cells of the pars intermedia were unstained for GK. However, some of the ACTH cells in the pars distalis did indicate the presence of GK. Similar to TSH, ACTH cells in the anterior pituitary were highly variable in staining for GK, but a small proportion of the hormone-positive cells were stained for GK. When a combination of the antibodies to FSH, LH, TSH, and ACTH was used, all cells with GK immunoreactivity could be accounted for by immunoreactivity of the hormones (Figure 5). GK immunoreactivity was not detected in cells containing either GH or PRL.
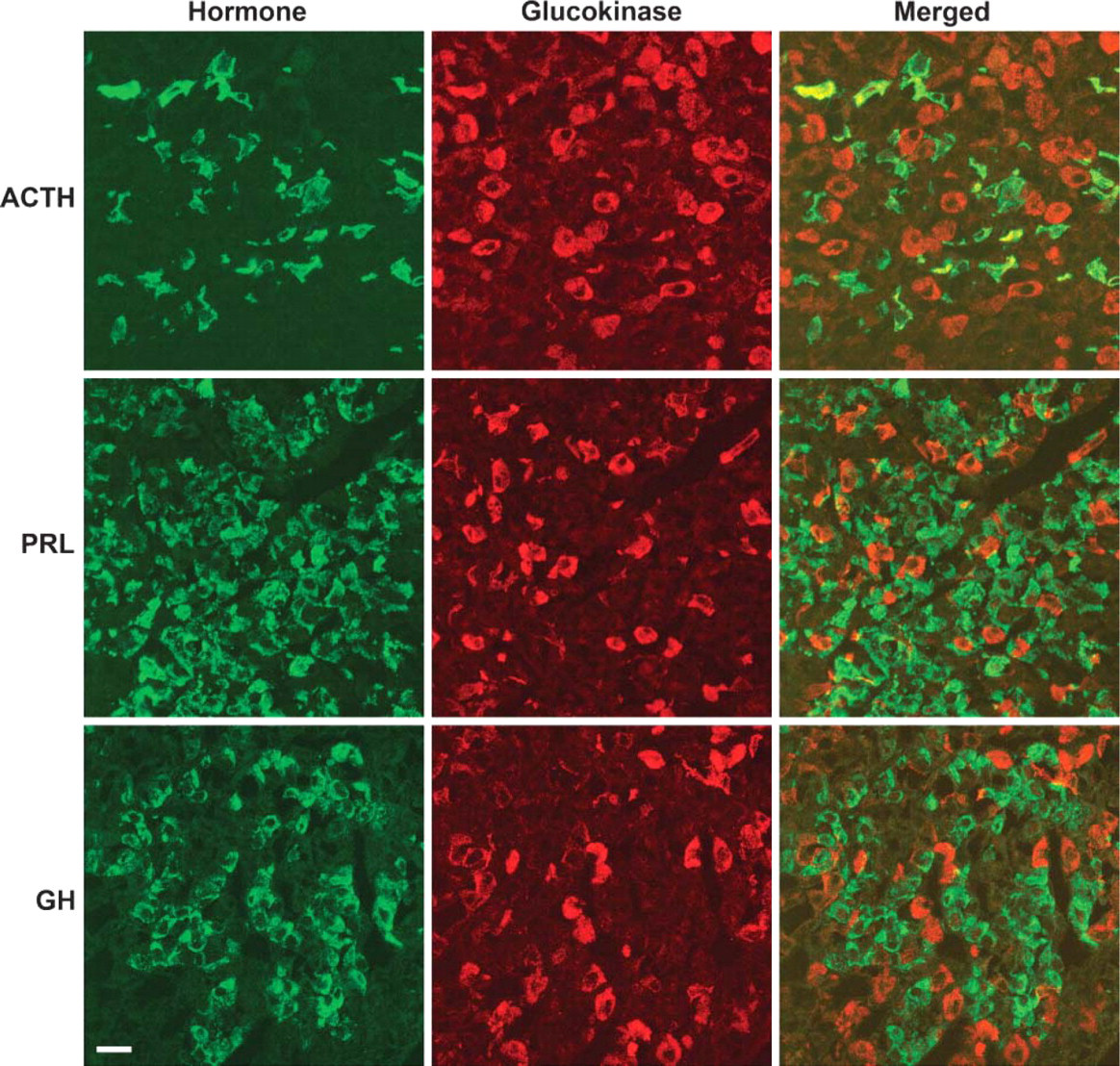
Rat pituitary double labeled with antibodies to GK (red) and either adrenocorticotropic hormone (ACTH), prolactin (PRL), or growth hormone (GH) (green). The irregularly shaped cells with less intense staining for GK were shown to be corticotropes containing ACTH (top). GK immunoreactivity was not detected in cells containing PRL (middle) or GH (bottom). Bar = 25 μm.
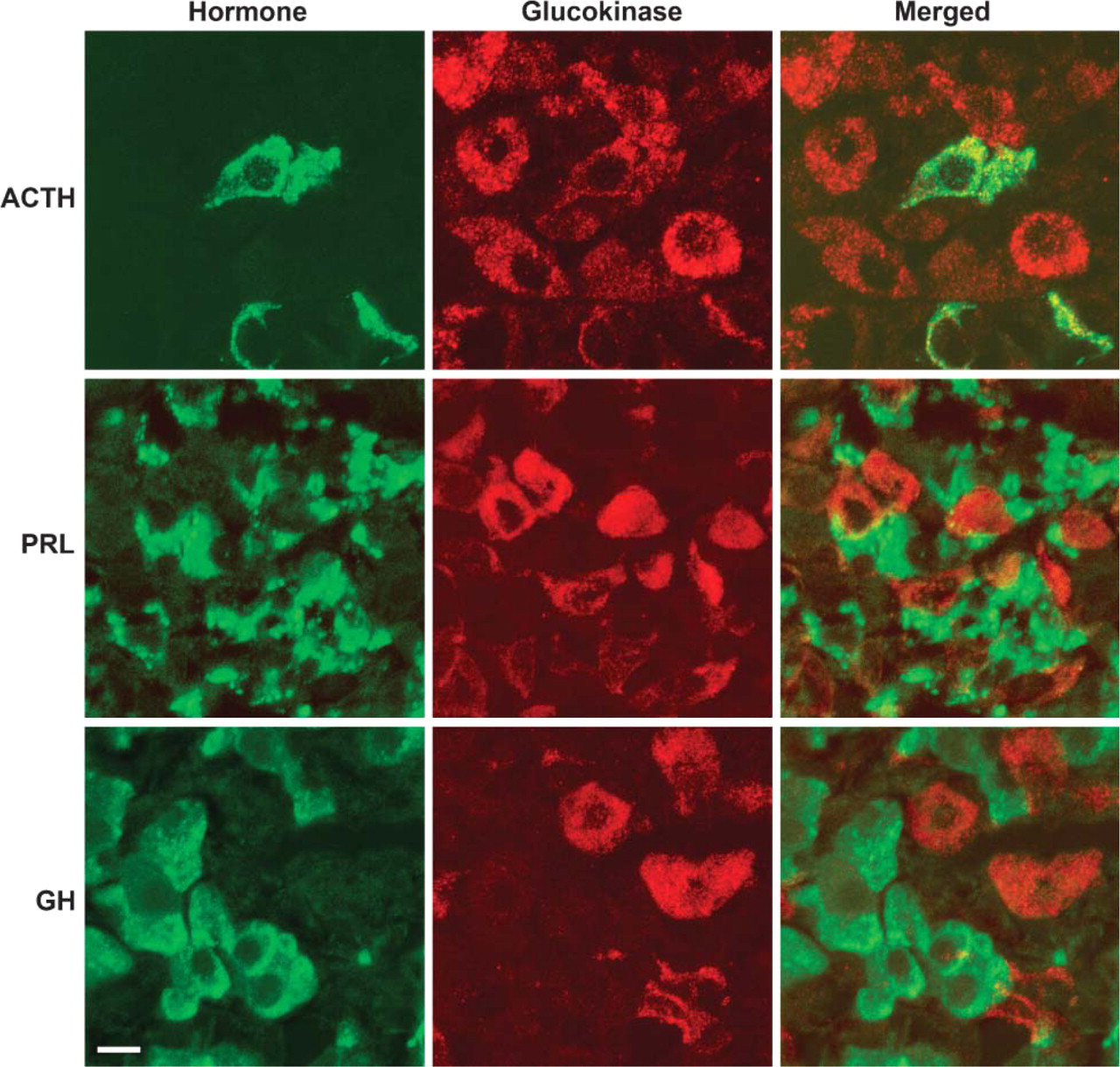
Higher magnification images from a rat pituitary double labeled with antibodies to GK (red) and either ACTH, PRL, or GH (green). The irregularly shaped cells with less intense staining for GK were corticotropes containing ACTH (top). Although intense PRL staining was often found around the large cells with GK immunoreactivity (middle), these fragments are actually brightly stained regions of the cytoplasm of adjacent lactotropes. A similar relationship was not observed with the somatrotropes containing GH (bottom). Bar = 10 μm.
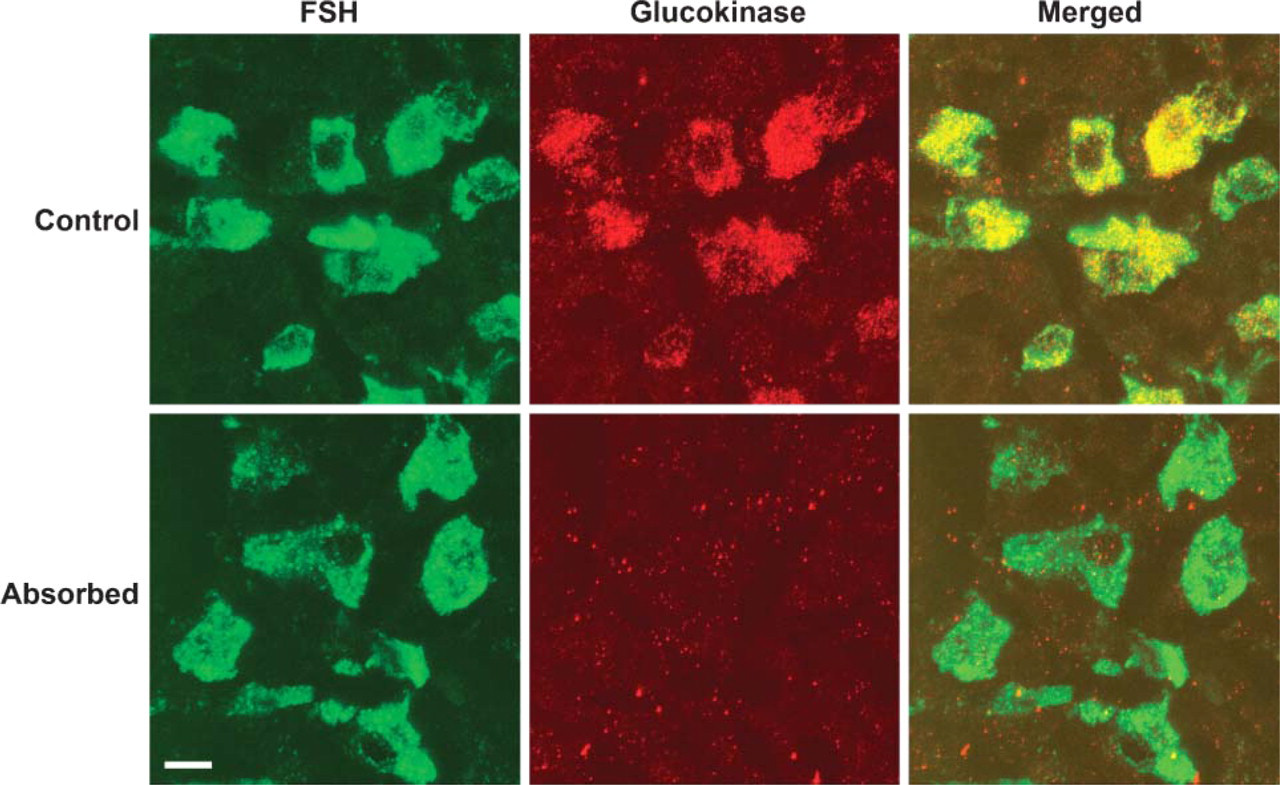
Absorption controls were done by preabsorbing the sheep anti-GK antibody with 2.5 nmol/ml GK, 10 nmol/ml glutathionyl-S-transferase (GST), or 10 nmol/ml GST-GK. Preabsorbing with GK or GST-GK reduced fluorescence intensity of GK by ~50%. Preabsorbing with GST alone had no effect on anti-GK fluorescence intensity. To determine whether there may be a nonspecific effect of the absorbing peptides, they were also preincubated with TSH antiserum. There was no effect of the absorbing peptides on the intensity of TSH staining.
Rat pituitary sections were also examined with a goat anti-GK antiserum. The staining pattern was similar to that previously observed with the sheep anti-GK antiserum. GK immunoreactivity was detected throughout the cytoplasm of a discrete population of cells in the anterior pituitary. Colocalization studies confirmed that GK was present in FSH- and LH-containing cells and not in cells with PRL or GH. Staining was completely blocked by preabsorbing the GK antibody with 200 μg/ml of the immunizing peptide (Figure 6).
Two monkey pituitaries were examined with both the sheep and goat anti-GK antisera. Unlike the rat pituitaries, GK immunoreactivity with the sheep anti-GK antiserum was confined to a small discrete perinuclear region. Only a few cells had the generalized staining of the cytoplasm for GK. The cells containing the perinuclear GK immunoreactivity also reacted to antibodies against FSH, LH, and TSH (Figure 7 and Figure 8). Similar to the rat, some ACTH-containing cells indicated the presence of GK. Perinuclear GK was not detected in cells immunoreactive for PRL or GH. When a combination of antisera against FSH, LH, TSH, and ACTH was used, all cells with perinuclear GK immunoreactivity were stained for these hormones. When a combination of PRL and GH was used, none of the cells immunoreactive for the hormones had detectable levels of immunoreactive perinuclear GK. The goat anti-GK antiserum did not detect the GK located in the perinuclear region of the monkey pituitary; however, it did detect cells with generalized cytoplasmic staining. The few cells with generalized cytoplasmic staining of GK were found to be a small subpopulation of the TSH- and PRL-containing cells.
Rat pituitary double labeled with antibodies to GK (red) and a combination of FSH/LH/TSH/ACTH. All cells containing GK could be accounted for by cells containing immunoreactivity for these hormones. Bar = 10 μm.
Rat pituitary double labeled with the goat anti-GK serum (red) and FSH (green). This antibody stained large cells containing FSH (top). Staining was completely blocked by preabsorbing the GK antibody with 200 μg/ml of the immunizing peptide (bottom). Bar = 10 μm.
Discussion
These immunohistochemistry experiments were undertaken to examine the cellular distribution of GK in the pituitary. Two GK antibodies were used in this study. The sheep anti-GK antibody was made against a fusion protein of GST-GK. This antibody was used without further purification. The goat anti-GK antibody was made against a 2000-Da peptide fragment from GK. In pituitaries from the perfusion-fixed rats, both antibodies detected GK immunoreactivity in a discrete population of cells in the anterior pituitary where it appeared as a generalized cytoplasmic immunofluorescence. When colocalization experiments were done, GK immunoreactivity corresponded with the cells containing FSH, LH, and TSH and a small portion of ACTH-containing cells. Only a small portion of the FSH- and LH-positive cells did not appear to contain GK. When a cocktail of FSH, LH, TSH, and ACTH antibodies was used, all the GK-positive cells could be accounted for by the FSH, LH, TSH, and ACTH cells. When GH or PRL antibodies were used, none of these cells demonstrated GK immunoreactivity. That two different antibodies to GK gave similar results supports the interpretation that the immunofluoresence seen with antibodies represents the presence of GK. Also, preabsorbing the antibodies with the immunizing peptides, significantly reducing or eliminating detection of GK, supports the fact that the tissue antigens detected were from GK.
Two of the rat pituitaries were fixed by immersion rather than perfusion. Immersion fixation resulted in a delay of 10 or more minutes after death of the animal. These pituitaries were examined with the sheep anti-GK antibody (data not shown). In these pituitaries, GK immunoreactivity was observed in the FSH, LH, and TSH cells. However, the staining pattern differed, in that prominent nuclear as well as cytoplasmic staining of GK was observed. Although it is not possible to rule out that the nuclear location of GK was a consequence of an altered physiological state, postmortem changes merit consideration. This suggests that rapid fixation, preferably by perfusion, is important in examining the cellular location and distribution of GK in the pituitary.
Two monkey pituitary glands were available for this study. Sheep anti-GK immunoreactivity was observed in a discrete population of cells in the anterior pituitary. When colocalization studies were done, cells positive for GK reacted with antibodies to FSH, LH, TSH, and ACTH. Perinuclear GK was not detected in cells containing GH or PRL. Thus, the cellular distribution corresponded with that observed in rat pituitaries. The distribution of GK within the cells differed, however, from that seen in rats. In the monkey, GK was confined to a small area close to the nucleus, oftentimes appearing in a circular profile or as filamentous strands. Its appearance and subcellular location suggests that it is associated with the Golgi complex. Supporting this interpretation is the observation that GK immunoreactivity was seen in the same region as β-COP, a marker for the Golgi apparatus (data not shown). A generalized cytoplasmic distribution of GK, comparable to that seen in rat pituitary cells, was only rarely observed. These cells represented a small portion of the TSH- and PRL-containing cells. The goat anti-GK antibody did not detect GK in the monkey pituitary that was restricted to the perinuclear location, but did detect the cells with generalized cytoplasmic immunoreactivity for GK. The reason for this is not apparent but suggests that the GK epitope that reacts with the goat antibody is sequestered when GK is restricted to the small perinuclear area. A Golgi form of GK has previously been reported in the liver. That study also showed that the hepatic Golgi GK did not cross-react with an antiserum specific for cytoplasmic GK (Berthillier et al. 1976).
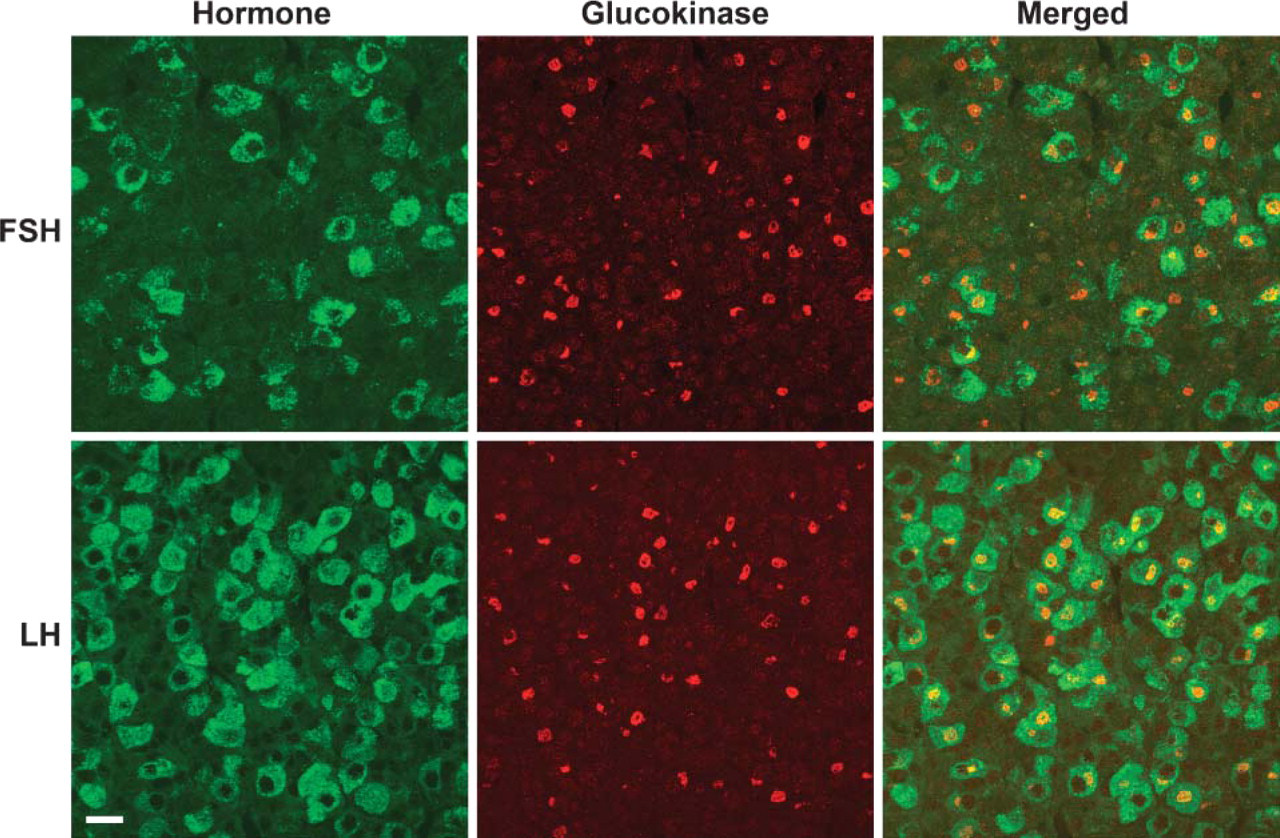
Monkey pituitary double labeled with antibodies to GK (red) and either FSH or LH (green). The large cells with GK immunoreactivity restricted to a small, discrete region were gonadotropes containing FSH (top) and/or LH (middle). Because the sections were only 7.0-μm thick, the apparent absence of GK immunoreactivity in some gonadotropes may be an artifact of the plane of section. Bar = 25 μm.
The presence of GK in gonadotropes and thyrotropes expands the network of GK-positive cells (Matschinsky et al. 2006). This body-wide system includes the hepatic parenchymal cells, which contain ~99% of the body's complement of the enzyme, the cells of the pancreatic islets of Langerhans and, most importantly, the insulin-producing β-cells but perhaps also the α- and δ-cells, entero-endocrine K- and L-cells, and cells in hypothalamic and other brain nuclei. The presence of the enzyme in cells of such widely divergent physiology suggests different roles. We tentatively identify three distinct functions of the enzyme: (1) the bulk of the enzyme present in the liver is part of a high-capacity system involved in glucose clearing from the blood and storage in the form of glycogen (Agius et al. 2004; Matschinsky et al. 2006); (2) GK in the cells of the pancreatic islets, the brain and, perhaps, the entero-endocrine K- and L-cells serves as a glucose sensor for minute to minute control of hormone release maintaining glucose homeostasis (Gribble et al. 2003; Levin et al. 2004; Matschinsky et al. 2006); and (3) GK of the gonadotropes and the thyrotropes may serve as a sensor monitoring the general nutritional status of the body and may thus manifest its role only when glucose falls to critically low levels and is maintained low for extended periods. This may occur as a result of forced or voluntary starvation, famine in the wake of natural disasters, or because of tumor cachexia, to list several obvious conditions causing hypogonadism, nutritional infertility, and altered thyroid function (Bates 1985; Bronson and Marsteller 1985; Schreihofer et al. 1993; Wade et al. 1996; Wade and Jones 2004). As alluded to in the Introduction, lowering of blood glucose and the associated decrease of glucose metabolism of peripheral hepatic and central hind brain glucose sensor cells are probably essential biochemical events that initiate the elaborate signaling sequence culminating in the reduction or cessation of pulsatile LH/FSH release from gonadotropes (Wade et al. 1996; Wade and Jones 2004). Our discovery of physiologically relevant amounts of GK in the gonadotropes suggests an additional level of control at the pituitary. The regulatory control strength of pituitary GK relative to that of other steps in the signaling pathway remains to be established.
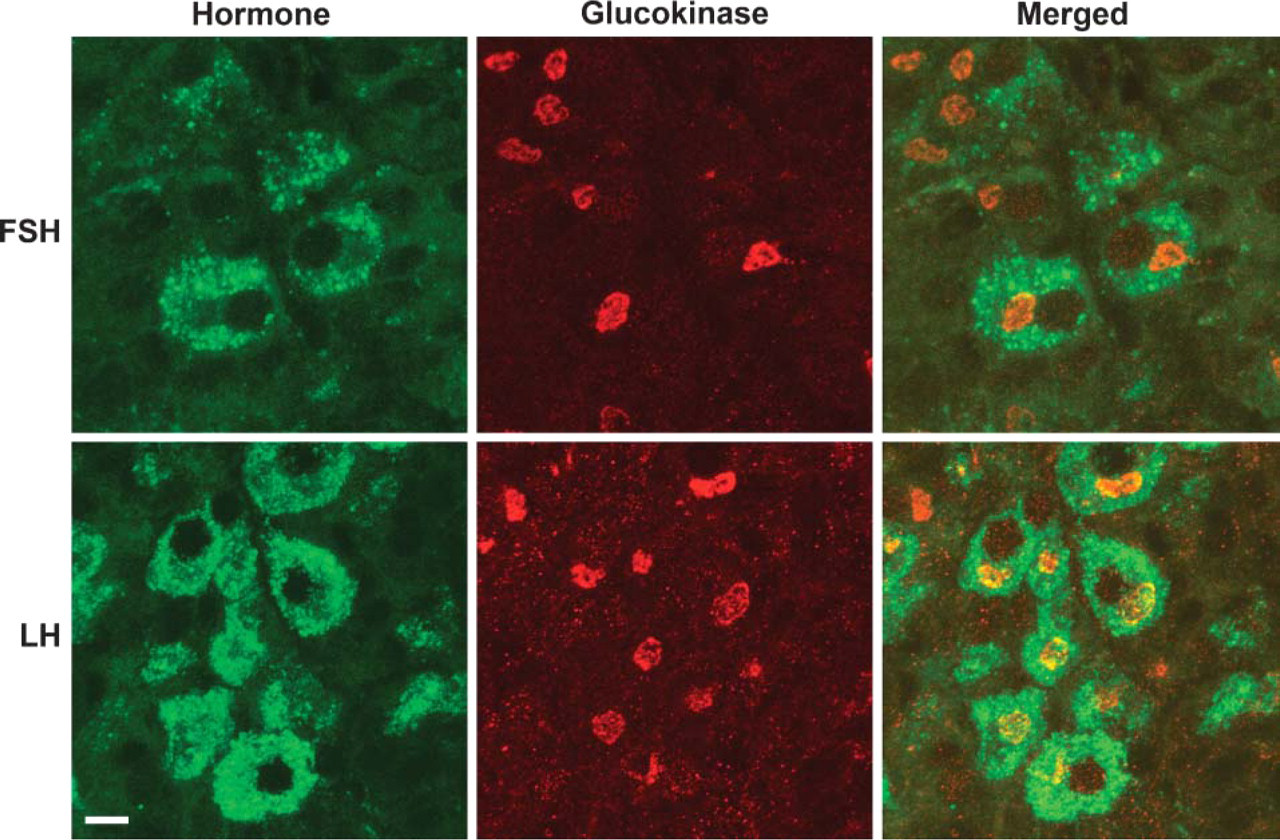
Higher magnification images from a monkey pituitary double labeled with antibodies to GK (red) and either FSH or LH (green). GK immunoreactivity was present as circular profiles and filamentous strands restricted to a perinuclear region that was lower in staining intensity for these hormones. Bar = 10 μm.
The molecular mechanism by which glucose alters the function of gonadotropes and thyrotropes remains to be elucidated. Glucose may exert effects on pituitary cells that are comparable to its actions on pancreatic β-cells, at least to some extent (Matschinsky et al. 2006). Similar to acetylcholine stimulation of insulin release from β-cells (Doliba et al. 2004, 2006) GnRH stimulaton of LH/FSH release from gonadotropes involves G-protein-coupled receptors that induce phospholipase C activity and elevate intracellular levels of inositol triphosphate, diacylglycerol, cyclic AMP, Ca2+, and associated activation of diverse protein kinase-mediated signaling pathways (Norman and Litwack 1997; Larsen et al. 2003). It is not unreasonable to speculate that glucose metabolism of gonadotropes controlled by GK may modify these processes in various ways including alterations of the phosphate-potential (ATP/ADPxPi), the intracellular pH, the redox-potential [NAD(P)H/NAD(P)], and level changes of other allosterically effective metabolites and cofactors, e.g., malonyl-CoA and 5′-AMP, all parameters thought to be critical for metabolic coupling in pancreatic β-cells (Prentki and Matschinsky 1987; Matschinsky 1995; Newgard and Matschinsky 2001; Matschinsky et al. 2006). It is also conceivable that glucose acts uniquely on pituicytes. One could speculate that GK regulates the pools of UDP-glucose and other sugar nucleotides in these cells and thereby influences the synthesis of the corresponding hormones, which are glycopeptides containing as much as 15% carbohydrate as one distinguishing feature of GK-positive pituitary cells. The apparent localization of GK in structures resembling the Golgi apparatus of monkey gonadotropes points in this direction.
In considering the potential physiological and pathological importance of direct regulation of pituicytes by glucose, we are struck by the lack of reported clinical symptoms of hypogonadism and hypothyroid-ism in patients with permanent neonatal diabetes linked to GK (PNDM-GK) (Njølstad et al. 2001, 2003). Patients with PNDM require insulin treatment for blood glucose control to survive because their β-cells are not functioning due to GK deficiency. The gonadotropes and thyrotropes of patients with homozygous GK deficiency can also be predicted to lack functional GK and should then show impaired function if the above speculation applies to human physiology or pathology. In view of the predominant hypothalamic regulation of pituitary cells, a direct modifier role of glucose as proposed here may not be readily apparent in routine clinical examinations, particularly when confronting a severe disease such as PNDM and may require specific and thorough testing of gonadotrope and thyrotrope function to manifest itself.
The presence of GK in gonadotropes and thyrotropes reported here could become a medical issue in the near future if the anticipated use of GKAs as anti-diabetic drugs becomes a reality (Grimsby et al. 2003; Matschinsky et al. 2006). It is likely that the drug would then be prescribed for use in large numbers of patients including also premenopausal women with type 2, and perhaps even with type 1, diabetes mellitus. In that event it will be possible and actually necessary to explore what effects GKAs may have on the physiology of gonadotropes and thyrotropes. It should also be obvious that the availability of GKAs affords an opportunity to assess the proposed glucose sensor role of GK in pituicytes experimentally in laboratory animals.
In summary, these immunohistochemical experiments support the hypothesis that GK is located in a discrete population of cells in the anterior pituitary. Most of the cells with GK immunoreactivity were gonadotropes. In addition, many thyrotropes also contained GK. The remaining GK-positive cells appear to be a very small subpopulation of corticotropes. With the exception of a very few cells in the monkey, GK was not detected in lactotropes or somatotropes. Even though the precise role that GK plays in the physiology of this selected group of pituitary cells remains to be elucidated, the presence of the enzyme suggests a direct influence of glucose at this level.
Footnotes
Acknowledgements
This work was supported by National Institutes of Health Grants DK-33655, DK-22122, and NS-04399.