
Abstract
The high intraluminal concentrations of HCO3
− in the human pancreatic ducts have suggested the existence of a membrane protein supplying the Cl−/HCO3
− exchanger. Membrane-bound carbonic anhydrase IV (CA IV) is one of the potential candidates for this protein. The difficulties in isolating human pancreatic ducts have led the authors to study the molecular mechanisms of HCO3
− secretion in cancerous cell lines. In this work, we have characterized the CA IV expressed in Capan-1 cells. A 35-kDa CA IV was detected in cell homogenates and purified plasma membranes. Treatment of purified plasma membranes with phosphatidylinositol-phospholipase-C indicated that this CA IV was not anchored by a glycosylphosphatidylinositol (GPI). In contrast, its detection on purified plasma membranes by an antibody specifically directed against the carboxyl terminus of human immature GPI-anchored CA IV indicated that it was anchored by a C-terminal hydrophobic segment. Immunoelectron microscopy and double-labeling immunofluorescence revealed that this CA IV was present on apical plasma membranes, and in the rough endoplasmic reticulum, the endoplasmic reticulum-Golgi intermediate compartment, the Golgi complex, and secretory granules, suggesting its transport via the classical biosynthesis/secretory pathway. The expression in Capan-1 cells of a 35-kDa CA IV anchored in the apical plasma membrane through a hydrophobic segment, as is the case in the healthy human pancreas, should make the study of its role in pancreatic HCO3
− secretion easier.
T
After stimulation with secretin, the ductal cells of the pancreas secrete a solution rich in HCO3 −. Several models have been proposed for the mechanisms involved in this secretion. These take into account the concentration of HCO3 − in the pancreatic juice. In the rat, where the intraluminal concentrations of HCO3 − ions are of the order of 70 mM after stimulation, it is accepted that HCO3 − ions accumulate in the cell, in part due to the activity of CA II, and partly due to their entry mediated by a basolateral Na+/HCO3 − cotransporter (Zhao et al. 1994). The HCO3 − ions are then secreted at the apical poles of the cells via a Cl−/HCO3 − exchanger (Novak and Greger 1988), which functions in parallel with the cystic fibrosis transmembrane conductance regulator (CFTR). However, this model cannot be applied to species such as the guinea pig, the cat, or man, in whom the concentration of HCO3 − in the pancreatic juice may reach 140–150 mM after secretin stimulation. In fact, in the presence of such high luminal concentrations, a Cl−/HCO3 − exchanger would ensure the absorption of HCO3 − ions rather than their secretion. This phenomenon is avoided due to the inhibition of the exchanger by high concentrations of HCO3 − (Ishiguro et al. 2000). It seems reasonable that another protein is implicated in the exit of HCO3 −. This hypothetical protein may be an HCO3 − channel (Ishiguro et al. 1996b, 2000, 2002) or a carbonic anhydrase (Hyde et al. 1999), more specifically, a CA IV (Fanjul et al. 2004). Most of these results have been obtained using pancreatic ducts isolated from guinea pigs (Ishiguro et al. 1996a, b, 2000, 2002). In man, given the difficulty of obtaining intact pancreatic ducts, different studies have been made using cancerous cell lines derived from pancreatic ductal tissue. Among these, the Capan-1 line, of ductal origin (Kyriazis et al. 1982), is considered to be a good experimental model of the human pancreatic ductal cell. Despite their neoplastic transformation, Capan-1 cells have retained numerous properties of normal ductal pancreatic epithelial cells (Levrat et al. 1988; Cheng et al. 1998). They polarize spontaneously and form domes, evidence of transepithelial exchange of H2O and electrolytes (Levrat et al. 1988). They express the CFTR Cl− channel (Becq et al. 1993; Hollande et al. 1998), an anionic outward-rectifying channel (Becq et al. 1992), a high-conductance anionic channel permeable to HCO3 − ions (Becq et al. 1992; Mahieu et al. 1994), an Na+/HCO3 − cotransporter (Shumaker et al. 1999), and a plasma membrane–associated CA II (Alvarez et al. 2001). They also possess a 55-kDa CA IV-immunoreactive protein that has been suggested to be involved in the secretion of HCO3 − ions (Mahieu et al. 1994).
With the aim of validating the Capan-1 cell line as a useful model for studying human pancreatic HCO3 − secretion, we investigated the expression of a 35-kDa CA IV and its subcellular distribution. We show that Capan-1 cells express a 35-kDa CA IV that is anchored in membranes by its carboxyl terminus, as is the case in the healthy human pancreas, and is targeted to the apical plasma membranes via the classical biosynthesis/secretory pathway.
Materials and Methods
Cell Culture
The human cancerous pancreatic ductal cell line Capan-1 (American Type Culture Collection, Rockville, MD) was established from a hepatic metastasis taken from a patient with a pancreatic adenocarcinoma. The cells were cultured in RPMI 1640 medium supplemented with 15% fetal calf serum, penicillin (100 U/ml), streptomycin (100 μg/ml), and fungizone (0.25 μg/ml) (all from Gibco BRL; Grand Island, NY). They were maintained by successive passages after trypsinization with 0.05% trypsin-0.02% EDTA (Gibco BRL) and seeded at 2.5 × 105 cells/ml in 25-cm2 and 80-cm2 flasks (Nunc; Roskilde, Denmark) or on glass coverslips in Leighton tubes. The media were renewed every 2 days. Possible contamination with mycoplasma was checked routinely on cells cultured without antibiotics by a PCR method (Mycoplasma PCR kits, Stratagene; La Jolla, CA) or by seeding on specific medium.
Antibodies
To characterize the CA IV expressed in Capan-1 cells, two polyclonal antibodies were generated: (1) a CA IV NH2 antibody directed against a synthetic peptide purified by HPLC and corresponding to a sequence of 18 amino acids at the NH2 terminal (residues 1–18: AESHWCYEVQAESSNYPC) of human mature GPI-anchored CA IV; and (2) a CA IV COOH antibody directed against a synthetic peptide purified by HPLC corresponding to a transmembrane region of 19 amino acids at the COOH terminal of human immature CAIV (residues 276–294: ALPALLGPMLACLLAGFLR). This region is cleaved in the rough endoplasmic reticulum (RER) during anchorage of human CA IV to GPI (Okuyama et al. 1992). After binding to ovalbumin, the peptides were injected into rabbits in a standard immunization procedure.
Double-labeling reactions were carried out using antibodies directed against markers of intracellular compartments: (1) a monoclonal antibody directed against the calnexin, an RER marker (Chemicon International Inc.; Temecula, CA) (Wada et al. 1991); (2) the monoclonal ERGIC 53 antibody directed against the ERGIC 53 protein, an endoplasmic reticulum-Golgi intermediate compartment (ERGIC) marker (a gift from Dr. P. Hauri, Department of Pharmacology, Biocenter of the University of Basel, Basel, Switzerland) (Schweizer et al. 1988); (3) a monoclonal antibody directed against the 58K protein, a membrane Golgi marker (clone 58K-9, Sigma Chemical Co.; St. Louis, MO) (Bloom and Brashear 1989); (4) a monoclonal antibody directed against γ-adaptin, a trans-Golgi network (TGN) marker (Transduction Laboratories; Lexington, KY) (Robinson 1990); and (5) a monoclonal 1.13M1 antibody directed against MUC5AC mucin, a secretory granule marker (a gift from Dr. J. Bara, INSERM U.55, Hôpital St Antoine, Paris, France) (Bara et al. 1998).
Immunocytochemistry
CA IV was detected in Capan-1 cells by immunofluorescence, immunogold staining, and immunoperoxidase reactions. To specify its subcellular distribution, double-labeling reactions of CA IV and intracellular compartments were also conducted.
Immunoelectron Microscopy Labeling
Cells fixed in situ with paraformaldehyde (PFA) (3%)-glutaraldehyde (GA) (0.1%) (30 min) were scraped and pelleted by centrifugation (10 min, 5000 × g). The pellets were dehydrated in increasing ethanol concentrations. This was followed by impregnation (2 hr and overnight) with LR White resin (EMS; Fort Washington, PA), then polymerization at 50C for 24 hr. Ultrathin sections collected on nickel grids were pretreated for 2 × 5 min on 20-μl droplets of Tris-HCl buffer (0.05 M, pH 7.6) and 15 min on 0.6% BSA-0.5% NaCl in Tris-HCl buffer. They were then incubated with the CA IV COOH antibody (1:50, 2 hr) in Tris-HCl-BSA (0.6%)-NaCl (0.5%) buffer. After washes (3 × 10 min), they were incubated with 10-nm gold-labeled protein A (1:20, 1 hr) (British BioCell International; Cardiff, UK), then rinsed with Tris-HCl-NaCl buffer (3 × 10 min). Sections were stained with aqueous 4% uranyl acetate (40 min) and examined using a Hitachi H600 electron microscope.
Transmission Electron Microscopy Immunoperoxidase
Cells were fixed (20 min, 4C) with PFA (2%)-GA (2.5%), permeabilized or not with Triton X-100 (0.1%, 1 min), and incubated with the CA IV COOH (1:100) antibody in Tris-HCl (0.05 M, pH 7.6)-BSA (0.1%) buffer (1 hr), followed by goat anti-rabbit IgG coupled with peroxidase (Sigma Chemical Co.) (1:400, 45 min). Peroxidase activity was revealed by 3,3′-diaminobenzidine (Sigma Chemical Co.). After washing, cells were treated with OsO4 (1%)-FeCNK (2.5%) (45 min, 4C), dehydrated, and embedded in Epon-Araldite resin. Ultrathin sections were examined using a Hitachi H600 electron microscope.
CA IV-Intracellular Compartment Double Labeling
Confluent cells, fixed with PFA (3%) and permeabilized with ethanol, were stained immunocytochemically using the CA IV COOH (1:100) antibody and subcellular marker antibodies to either calnexin (1:50), 58K (1:50), ERGIC 53 (1:800), γ-adaptin (1:50), or MUC5AC (1:500). Antigen–antibody complexes were revealed with goat anti-rabbit IgG immune serum coupled with FITC (1:400) (Nordic Immunological Laboratories; Tilburg, The Netherlands) for CA IV, and goat anti-mouse IgG coupled with tetramethyl rhodamine isothiocyanate (TRITC) (1:100) (Nordic Immunological Laboratories) for the markers of various intracellular compartments.
The immunofluorescence preparations were observed using a confocal laser scanning microscope (Carl Zeiss LSM 410; Jena, Germany) equipped with argon (wavelength 488 nm) and helium (wavelength 543 nm) lasers.
Controls
The specificity of the immunocytochemical reactions was checked: (1) by using the CA IV COOH antibody fully saturated by an excess of the corresponding synthetic peptide (10−4 M); and (2) by incubating solely with the goat anti-rabbit IgG coupled with peroxidase, or with the gold-labeled protein A, or with a mixture of goat anti-rabbit IgG coupled with FITC and goat anti-mouse IgG coupled with TRITC.
Immunoblotting
Cellular Homogenates. Confluent cultures were rinsed, then scraped off into PBS. After centrifugation (5 min, 1500 × g), cells were lysed in 1 ml of buffer (HEPES 20 mM, NaCl 150 mM, EDTA 1 mM, aprotinin 25 μg/ml, leupeptin 10 μg/ml, pepstatin 15 μg/ml, PMSF 1 mM, DNase 2 μg/ml) containing 1% octyl-β-D-glucopyranoside (OTG) (Sigma Chemical Co.) for 30 min, 24 hr, or 48 hr at 4C. After centrifugation (5 min, 14,000 × g), the supernatants were considered as cell homogenates. Proteins (50 μg) were separated on a 10% SDS PAGE then electrotransfered onto a nitrocellulose membrane. After blocking with Tris-buffered saline (TBS)-Tween 20 (0.1%)-skim milk (5%), CA IV was detected by incubating the nitrocellulose membranes successively with the CA IV NH2 (1:3000) antibody or the CA IV COOH (1:2000) antibody in TBS-Tween-milk overnight at 4C, then with a goat anti-rabbit IgG immune serum coupled to peroxidase (1:10,000) (Pierce; Rockord, IL). Antigen–antibody complexes were revealed by enhanced chemiluminescence (ECL; Amersham, Buckinghamshire, UK).
To check the specificity of the CA IV NH2 and CA IV COOH antibodies, we performed immunoblots using each antibody on cell homogenates (50 μg of protein per lane) with 1 μg of purified bovine CA II (Sigma Chemical Co.), and 1 μg of a recombinant form of CA IV corresponding to the secreted form of human CA IV, which is normally GPI-anchored (a gift from Dr. Sly, Department of Biochemistry and Molecular Biology, St. Louis University School of Medicine, St. Louis, MO), and 1 μg of a purified human CA IX (a gift from Dr. Zavada, Institute of Molecular Genetics, Academy of Sciences of the Czech Republic, Prague) as migration controls. In addition, we carried out the following controls: (1) incubation with the CA IV antibody fully saturated with the relevant synthetic peptides (10−4 M), and (2) incubation in the presence of the peroxidase-linked anti-IgG antibody alone.
Determination of the Mode of Anchorage of CA IV in Plasma Membranes
Isolation of Plasma Membranes. Plasma membranes of confluent Capan-1 cells were purified on a continuous Percoll gradient according to the method of Record et al. (1982). Briefly, 8 × 107 cells, placed in Tris buffer (25 mM, pH 9.6, KCl 100 mM, MgCl2 5 mM, ATP 1 mM) were sonicated, then centrifuged (1000 × g, 10 min). The supernatant was taken up in 18 ml of a mixture of Percoll (11 ml), H2O (2.2 ml), Tris (100 mM), pH 7.4, KCl (400 mM), and MgCl2 (20 mM), adjusted to pH 9.6, then centrifuged (160,000 × g, 10 min) (60 Ti rotor, Beckman). The Percoll was removed from the membrane fraction by two successive centrifugations at 130,000 × g for 30 min each. All buffers contained leupeptin (10 μg/ml), aprotinin (25 μg/ml), pepstatin (15 μg/ml), PMSF (1 mM), and DNase (1 mM). Immunoblots of fractions of the purified plasma membranes were performed as described above.
To determine whether CA IV was bound to the plasma membranes of Capan-1 cells by a phosphatidylinositol (PI) residue, extracts of the purified plasma membranes were treated with phosphatidylinositol phospholipase C (PI-PLC). The fraction containing 50 μg of proteins was incubated in the presence of PI-PLC (0.2 mUE, B. thurigiensis, ICN Pharmaceuticals; Costa Mesa, CA) in Tris-SO4 − buffer (100 mM, pH 7.5, sodium deoxycholate 0.1%, 2 hr, 37C) with stirring, then centrifuged (30,000 rpm, 90 min; 60 Ti rotor, Beckman). After concentration by ultrafiltration (Ultra-free-L, Millipore Co.; Bedford, MA). the pellet and supernatant fractions were taken up in the loading buffer. The immunoblots were revealed using the CA IV NH2 (1:3000) antibody. For a control of the PI-PLC activity, we carried out immunoblots using an antibody directed against placental alkaline phosphatase (pAP) (1:2000), a GPI-anchored protein that is found on the plasma membranes of Capan-1 cells (Fanjul et al. 1991).
In addition, immunocytochemical reactions were carried out on cells treated with PI-PLC. Briefly, Capan-1 cells fixed with PFA (3%), were incubated at 37C with PI-PLC (0.2 UE) in Tris-SO4 − buffer, 100 mM (pH 7.5), and then incubated with either the CA IV NH2 antibody or the pAP antibody to verify the activity of the PI-PLC. Immunocytochemical reactions on cells preincubated in buffer without PI-PLC served as a control.
Results
Immunoblots of homogenates of Capan-1 cells lysed in the presence of OTG for 30 min (Figures 1A and 1B, Lanes 1), 24 hr (Figures 1A and 1B, Lanes 2) or 48 hr (Figures 1A and 1B, Lanes 3) showed the presence of a 35-kDa CA IV-immunoreactive band, which was detected by both the antibody directed against the N terminus of CA IV (Figure 1A) and the antibody directed against its carboxyl terminus (Figure 1B). A slight reduction in the intensity of this band was observed in extracts from cells lysed for 48 hr (Figures 1A and 1B, Lanes 3). A second band at 55 kDa was also revealed by both antibodies. This band was intense in the extracts of cells lysed for 30 min (Figures 1A and 1B, Lanes 1), was diminished in extracts prepared by lysis for 24 hr (Figures 1A and 1B, Lanes 2), and was totally absent in extracts of cells lysed for 48 hr (Figures 1A and 1B, Lanes 3).
The specificity of the two anti-CA IV antibodies was verified by the absence of cross-reactivity with CA II (Figures 1C and 1D, Lanes 3) and CA IX (Figures 1C and 1D, Lanes 4), two other CA isoforms expressed by Capan-1 cells. Furthermore, a recombinant CA IV corresponding to the human secreted 32-kDa CA IV was recognized by the CA IV NH2 antibody (Figure 1C, Lane 2). Two other bands, of 18 and 20 kDa, attributed to cleavage products of CA IV (Waheed et al. 1996), were also seen (Figure 1C, Lane 2). On the other hand, the CA IV COOH antibody did not recognize the re-combinant form of secreted CA IV (Figure 1D, Lane 2).
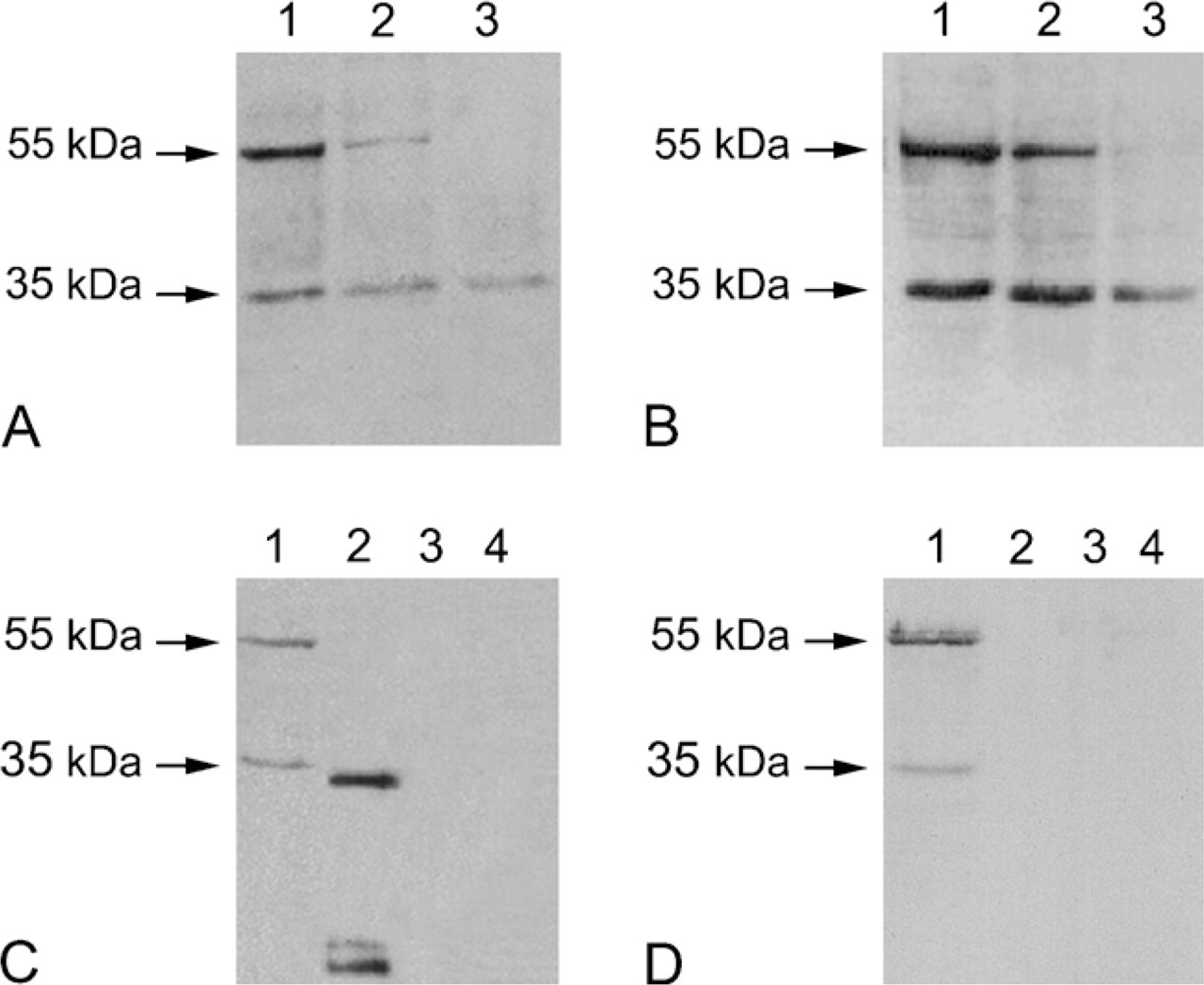
Characterization of the carbonic anhydrase IV (CA IV) expressed in Capan-1 cell immunoblots revealed with the CA IV NH2 antibody (
Anchorage of CA IV in the Plasma Membrane
In an attempt to ascertain whether the 35-kDa CA IV expressed in Capan-1 cells was anchored in the plasma membrane by GPI linkage, purified extracts of plasma membranes were treated with PI-PLC. Immunoblots were then made using the CA IV NH2 antibody (Figure 2A). Without treatment with PI-PLC, the 35-kDa CA IV was observed in the membrane fraction (Figure 2A, Lane −, M), and no protein was detected in the soluble fraction (Figure 2A, Lane −, S). After the action of PI-PLC, the 35-kDa CA IV was found to be still associated with plasma membranes (Figure 2A, Lane +, M), and no immunoreactive CA IV protein was detected in the soluble fraction (Figure 2A, Lane +, S). To check the activity of PI-PLC, similar procedures were conducted on the same samples for pAP, which is known to be GPI-anchored and expressed by Capan-1 cells (Fanjul et al 1991). Without treatment with PI-PLC, a band of 70 kDa was observed in the membrane fraction (Figure 2B, Lane +, M), and no protein was detected in the soluble fraction (Figure 2B, Lane +, S). After treatment with PI-PLC, although a fraction of pAP remained associated with the membrane fraction (Figure 2B, Lane +, M), a great part was seen in the soluble fraction (Figure 2B, Lane +, S). These observations were further supported by immunofluorescence labeling with both the CA IV NH2 and anti-pAP antibodies after treatment of fixed, non-permeabilized cells with PI-PLC. No decrease in CA IV immunoreactivity was observed after PI-PLC treatment, in contrast to the reduction in pAP immunoreactivity (data not shown).
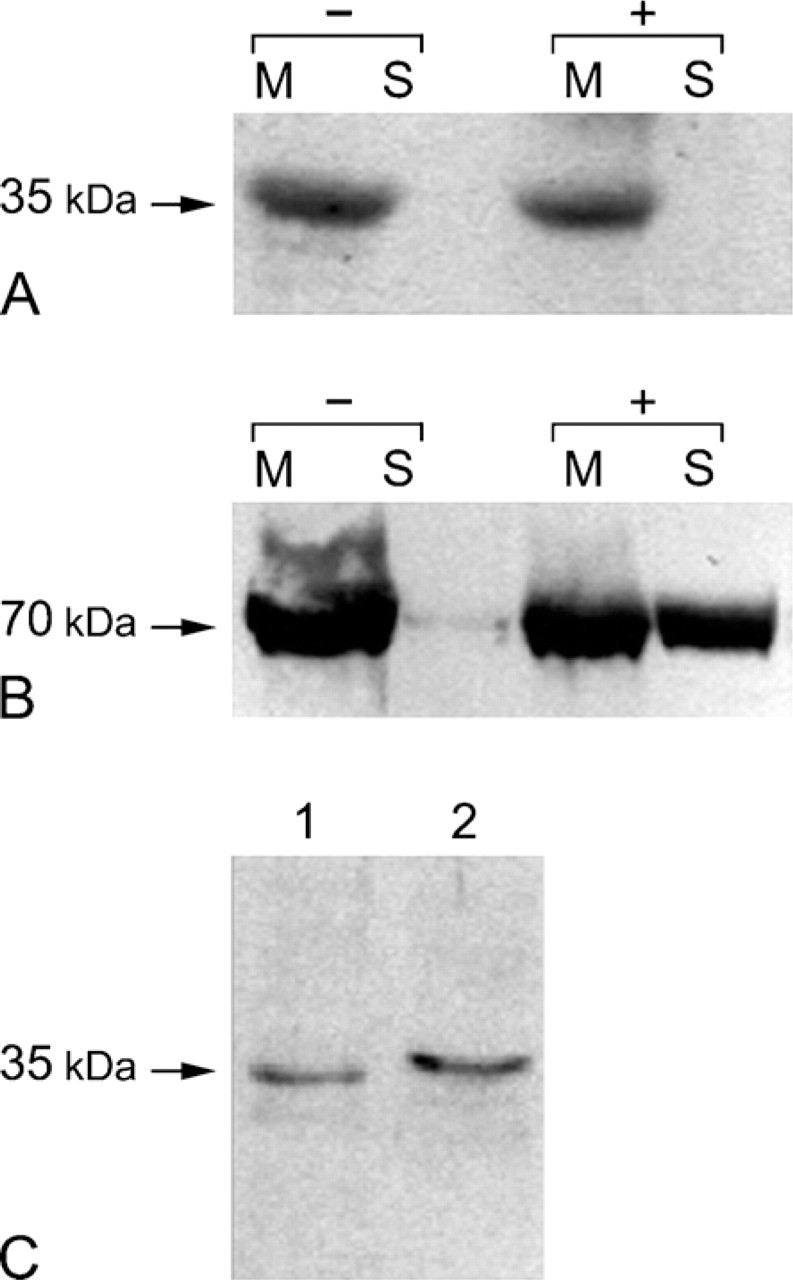
Determination of the mode of anchorage of the CA IV in plasma membranes. (
Localization of CA IV on apical plasma membranes of Capan-1 cells. (
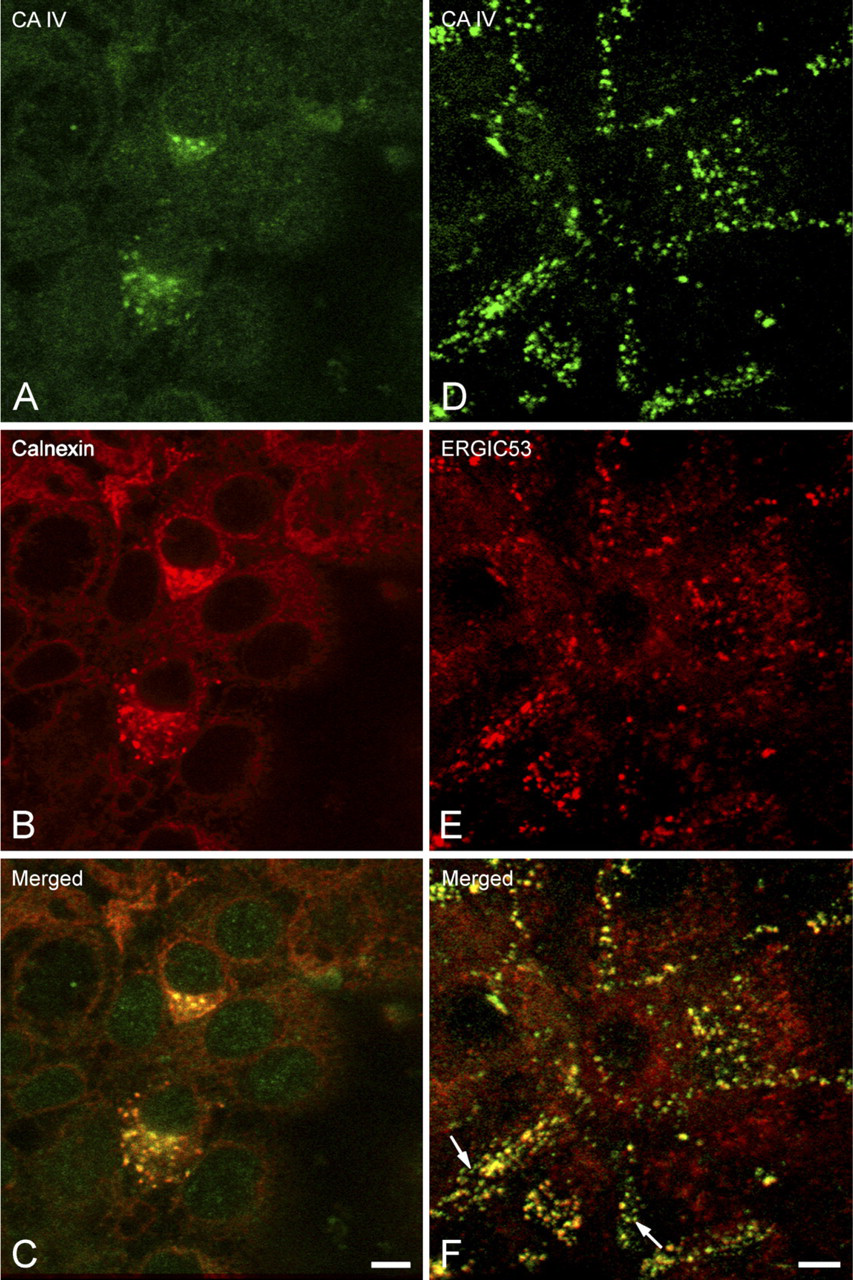
Confocal microscopy examination of the distribution of CA IV detected with the CA IV COOH antibody, in the rough endoplasmic reticulum (RER) and endoplasmic reticulum-Golgi intermediate compartment (ERGIC) of polarized Capan-1 cells. (
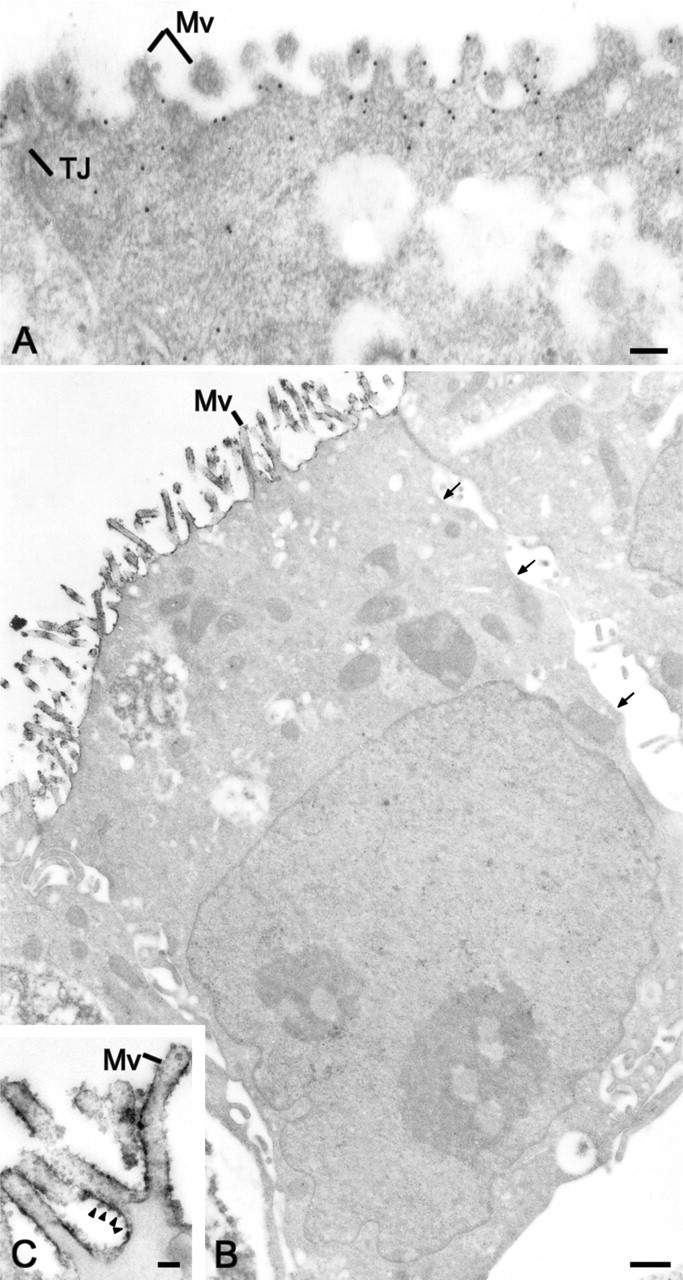
The use of the CA IV COOH antibody allowed the determination of the type of anchorage of the 35-kDa CA IV expressed in Capan-1 cells. Immunoblots of purified plasma membranes probed with the CA IV COOH antibody showed a 35-kDa band (Figure 2C, Lane 1) similar to that observed in the total protein extracts (Figure 2C, Lane 2). Immunogold staining (Figure 3A) and immunoperoxidase (Figures 3B and 3C) using the CA IV COOH antibody confirmed the presence of a CA IV anchored by its COOH terminus in the plasma membranes of polarized Capan-1 cells. CA IV immunoreactivity was seen on the surface of apical plasma membranes (Figures 3A and 3B), whereas no staining was observed along basolateral membranes (Figure 3B, small arrows). With immunoperoxidase reactions, the staining pattern was either a fine edge or dots spaced unevenly along the plasma membrane (Figure 3C, arrowheads).
Intracellular Distribution of the CA IV
Double-labeling reactions examined by confocal microscopy allowed the colocalization of the CA IV anchored by its carboxy terminal with markers of the RER (Figures 4A–4C), the ERGIC (Figures 4D–4F), the Golgi apparatus (Figures 5A–5C), the TGN (Figures 5D–5F), and the secretory granules (Figures 6A–6C). In polarized cells, observation of focal planes passing through the medial regions showed a CA IV immunoreactivity that presented as small punctate or elongated structures, preferentially grouped in a perinuclear cytoplasmic zone (Figures 4A, 4D, 5A, and 5D). These structures were also revealed by the antibodies directed against the calnexin (Figure 4B), the ERGIC 53 protein (Figure 4E), the 58K protein (Figure 5B), and the γ-adaptin (Figure 5E). Superimposing the staining patterns of these different markers with that of the CA IV immunoreactivity showed that most of immunofluorescent structures were colocalized (Figures 4C, 4F, 5C, and 5F). Some spots with CA IV immunofluorescence alone were observed (Figures 4F, 5C, and 5F, arrows). In planes passing through the supranuclear cytoplasm, MUC5AC staining of the secretory granules (Figure 6B) was colocalized (Figure 6C) with a part of the CA IV immunoreactivity (Figure 6A). In some cells, only CA IV immunofluorescence was observed (Figure 6C, arrows).
Transmission electron microscopy immunostaining with the CA IV COOH antibody showed labeling of the different membrane compartments involved in the trafficking of membrane and secretory proteins. Staining was observed on the membranes of the RER cisternae by immunoperoxidase reaction (Figure 7A, arrows and 7C, arrowheads) or immunogold staining (Figure 7B, arrows), as well as on Golgi complex membranes (Figure 7C, arrow). CA IV immunoreactivity was also observed on the membranes of secretory granules (Figure 7D, arrows) and vesicles localized in apical cytoplasms of polarized Capan-1 cells. Some stages of fusion between the secretory granule membrane and the CA IV-stained apical plasma membrane were observed (Figure 7D, arrowheads).
Discussion
In this work, we show that Capan-1 cells express a 35-kDa CA IV that is anchored in the apical plasma membranes by a hydrophobic segment at its carboxyl terminus, and targeted via the classical biosynthesis/secretory pathway.
The CA IV expressed in Capan-1 cells was characterized using two antibodies: one directed against a peptide sequence within the N terminus of the human GPI-anchored CA IV, and the other against the C-terminal transmembrane segment that anchors CA IV in the RER before cleavage and association with a GPI. The specificity of these two antibodies was established by the absence of cross-reaction with two CA isoforms expressed in Capan-1 cells, CA II (Mahieu et al. 1994; Alvarez et al. 2001) and CA IX (unpublished data), and by the recognition of a recombinant secreted form of human CA IV, which is normally GPI-anchored, by the CA IV NH2 antibody and not the CA IV COOH antibody. These two antibodies revealed the presence of two CA IV-immunoreactive proteins with apparent molecular mass of 55 kDa and 35 kDa in Capan-1 cells. The molecular mass of the latter protein corresponds to that of the CA IV previously described in different human tissues (Zhu and Sly 1990; Parkkila et al. 1996; Christie et al. 1997), including the ductal epithelium of the normal human pancreas (Fanjul et al. 2004). With regard to the 55-kDa CA IV-immunoreactive protein, the question arises as to whether it represents a genuine CA IV. This 55-kDa protein was the only form previously detected in Capan-1 cell homogenates (Mahieu et al. 1994) and human pancreatic samples (Carter et al. 1990) prepared without the use of detergent. In the present study, the use of detergent has allowed the demonstration of a 35-kDa CA IV-immunoreactive protein in addition to the 55-kDa form. These results are in agreement with those previously described in another human pancreatic ductal cell line (CFPAC-1 line) (Fanjul et al. 2002). In the same way, the use of detergent has permitted the evidence of the 35-kDa protein in healthy human pancreas (Fanjul et al. 2004). It is notable that increasing the lysis time up to 48 hr leads to a reduction in the level, and ultimately the disappearance, of this 55-kDa protein in Capan-1 cell homogenates. On the other hand, in extraction protocols designed to isolate the plasma membranes of Capan-1 cells, we did not detect the 55-kDa band. The sensitivity of this 55-kDa protein to detergents suggests that it may correspond to the formation of a protein complex including the 35-kDa CA IV and not a 55-kDa CA IV. These results are in agreement with those described in retinal cells (Wolfensberger et al. 1994) and in erythrocytes (Wistrand et al. 1999). In these models, under certain lysis conditions, the authors observed the disappearance of a 55-kDa CA IV-immunoreactive protein and the appearance of a 35-kDa or 45-kDa band. In other tissues, such as the esophagus, Christie et al. (1997) have also detected a CA IV-immunoreactive protein with a molecular mass above 35 kDa (60 to 74 kDa), which they have considered as a CA IV-cross-reactive protein. However, the fact that the 55-kDa protein was detected in Capan-1 cells by antibodies directed against different regions of human CA IV argues against a cross-reaction. Further studies are needed to better characterize this 55-kDa CA IV-immunoreactive protein.
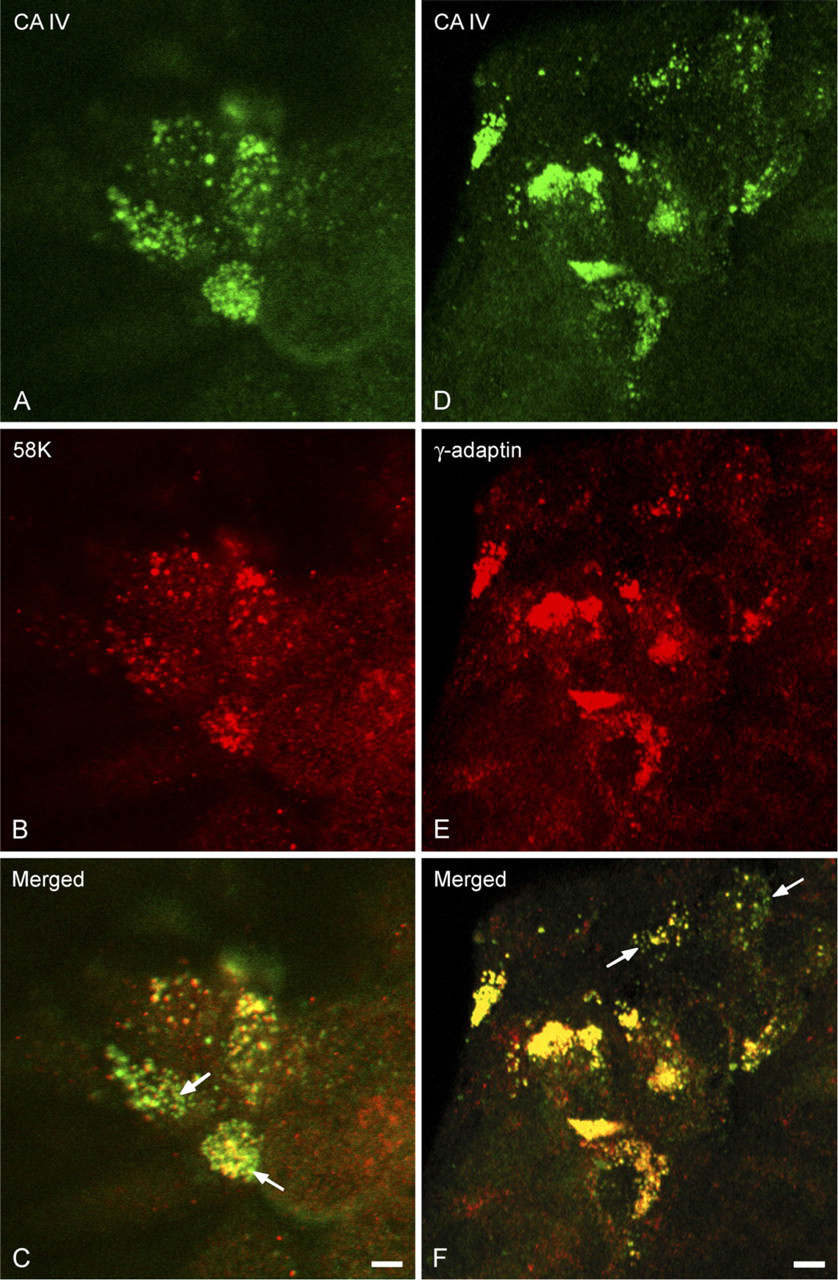
Confocal microscopy examination of the distribution of CA IV detected with the CA IV COOH antibody, in the Golgi complex and trans-Golgi network (TGN) of polarized Capan-1 cells. CA IV immunoreactivity (
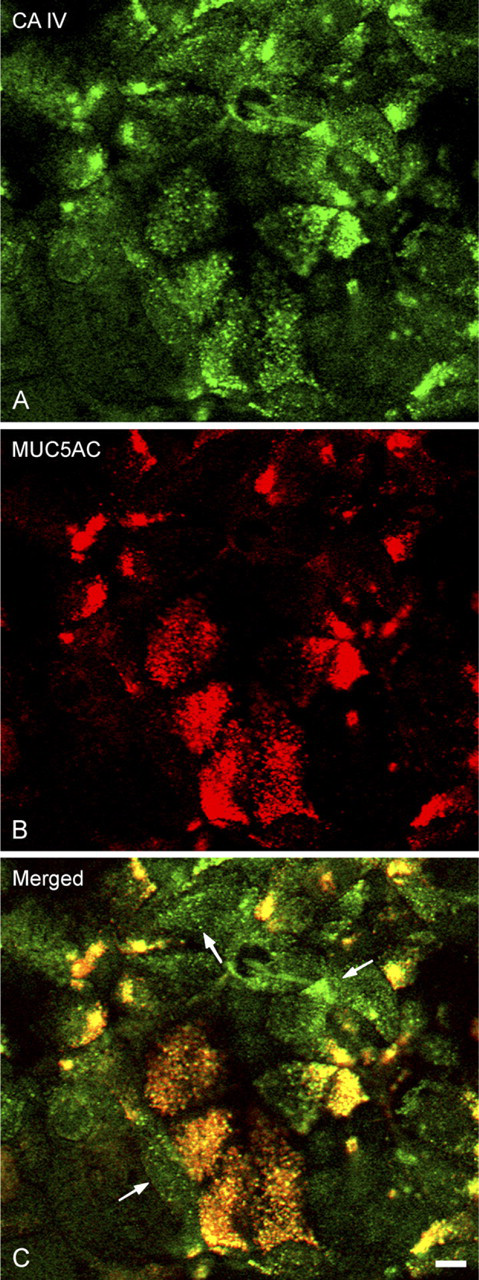
Confocal microscopy examination of the distribution of CA IV detected with the CA IV COOH antibody in secretory granules. Focal plane passing through supranuclear cytoplasms and showing colocalization (
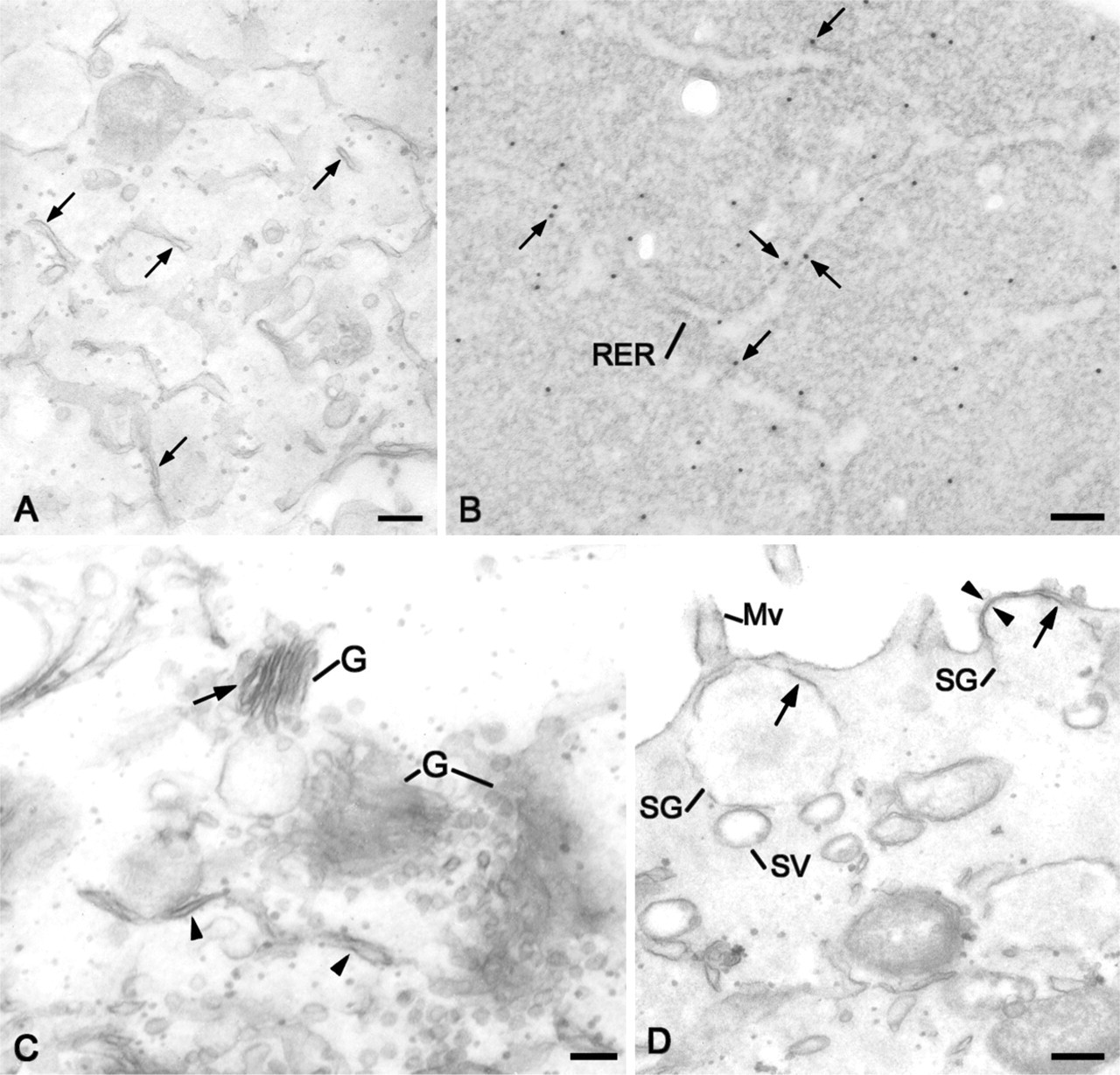
Distribution of CA IV in subcellular compartments by transmission electron microscopy immunoreactions using the CA IV COOH antibody. (
The human 35-kDa CA IV expressed in various tissues has generally been described as a GPI-anchored protein (Zhu and Sly 1990; Sender et al. 1994). With regard to CA IV expressed in Capan-1 cells, the experiments conducted either by immunocytochemistry or by immunoblotting after treatment with PI-PLC failed to show a GPI anchorage. The use of an antibody directed against the hydrophobic C-terminal segment of human immature CA IV allowed us to specify the mode of anchorage of CA IV in Capan-1 cells. By immunoblotting, this antibody revealed a 35-kDa protein in plasma membrane fractions. Moreover, electron microscopy immunolabeling showed CA IV COOH immunoreactivity mainly localized on apical plasma membranes. These findings suggest that CA IV expressed in Capan-1 cells is anchored in plasma membranes via the hydrophobic segment within its carboxyl terminal. The absence of GPI anchorage may be due to the cancerous nature of Capan-1 cells. It is known that colon cancer cells are not capable of anchoring proteins to a GPI (Chen et al. 2001). This defect results from a mutation in the PIGA gene that encodes a protein required for the first step in GPI anchor assembly. This leads to immature proteins, which remain anchored through the carboxyl terminal domain. In Capan-1 cells, this is unlikely, because we have demonstrated a GPI anchorage for pAP. Furthermore, we have described a 35-kDa CA IV anchored via its hydrophobic C-terminal segment in ductal cells from human healthy pancreas (Fanjul et al. 2004). In Capan-1 cells, the immunoreactivity visualized with the CA IV COOH antibody in the RER, and also in Golgi compartments and secretory granules, provides further evidence that this isoform of CA IV conserves its hydrophobic C-terminal peptide after passing the RER. Moreover, this subcellular distribution of CA IV is indicative of its targeting to apical plasma membranes by the classical secretory pathway. The expression of CA IV anchored in plasma membranes by a hydrophobic sequence has been reported in human eryrthrocytes, where it was considered to be a GPI-anchored CA IV isoform (Wistrand et al. 1999). It is known that certain proteins, such as N-CAM (Powell et al. 1991), or decay-accelerating factor (DAF) (Nonaka et al. 1995) can exist as several isoforms, including a GPI-linked form and a transmembrane form. These isoforms are thought to derive from alternative splicing (Cunningham et al. 1987; Nonaka et al. 1995). With regard to the CA IV expressed in pancreatic ductal cells, further experiments will be necessary to determine its exact origin.
The expression in Capan-1 cells of a CA IV anchored in membrane by a hydrophobic sequence similar to that described in healthy human pancreas (Fanjul et al. 2004) provides further support for considering this cell line a useful model for the study of pancreatic HCO3 − secretion. However, the physiological role of this CA IV has yet to be elucidated. In view of its localization on the outer leaflet of apical plasma membranes of ductal cells, it probably participates in the regulation of intra-luminal HCO3 − concentrations by catalyzing the reversible reaction H2O + CO2 HCO3 − + H+. It may provide locally and rapidly the production of HCO3 − to attain concentrations approaching 150 mM. We propose that this CA IV could be the extracellular component of an HCO3 − transport metabolon in association with cytosolic CA II and HCO3 −/Cl− exchanger. Indeed, we have described in Capan-1 cells the presence of a CA II associated with the cytosolic leaflet of apical plasma membranes (Alvarez et al. 2001), and Cheng et al. (1998) have proposed the involvement of an apical Cl−/HCO3 − exchanger in Cl− and HCO3 − secretion in these cells. The functionality of such a complex has been demonstrated in transfected HEK293 cells (Sterling et al. 2002). Another possibility is that this CA IV could be associated with an HCO3 − channel, in particular a high-conductance (350 pS) HCO3 − channel, that we have detected in the apical plasma membrane of Capan-1 cells (Becq et al. 1992; Mahieu et al. 1994). Further experiments are necessary to verify these hypotheses.
Taken together, the results presented in this study using the Capan-1 cell line are in agreement with the recent demonstration of the existence of a 35-kDa CA IV in normal human pancreatic ductal cells. These observations will facilitate the study of the role of CA IV in pancreatic HCO3 − secretion in man, and confirm the Capan-1 line as a good experimental model of the normal pancreatic ductal cell.
Footnotes
Acknowledgements
This investigation was partly supported by the French Association “Vaincre la Mucoviscidose.”