
Abstract
We investigated carbonic anhydrase IV (CA IV) in rat and human heart with immunohistochemical methods by both light and electron microscopy. In cryosections that were incubated with anti-CA IV/FITC, the capillaries showed a strong reaction for CA IV. In paraffin and semithin sections treated with anti-CA IV/ABC (avidin-biotin-peroxidase complex) blood vessels, capillaries, and sarcolemma (SL) were positively stained. By staining ultrathin sections with anti-CA IV/immunogold, CA IV could also be demonstrated at the latter two locations, including the specialized sarcolemmal structures intercalated discs, and T-tubules. In addition, by this method CA IV was seen to be associated with the sarcoplasmic reticulum (SR). The absence of immunostaining in SR and/or SL with some techniques probably indicates a problem of accessibility of the antigenic sites. In line with the immunohistochemical results, CA IV mRNA expression was visualized in both endothelial and muscle cells by in situ hybridization histochemistry.
Keywords
S
A membrane-bound CA, CA IV, has been shown by immunohistochemistry to be present in capillary endothelium of rat and human skeletal muscles (Sender et al. 1994) and in sarcolemma (SL) and sarcoplasmic reticulum (SR) of rat muscle (Decker et al. 1996). CA IV was also detected by immunoblotting in rat skeletal and heart muscle (Waheed et al. 1992). With a modified Hansson technique, Vandenberg et al. (1996) showed CA to be present in capillary endothelium and SL of ferret heart and, from perfusion experiments, inferred a significant role for CA in cardiac pH regulation.
The purpose of this study was to investigate in heart muscle the localization of membrane-bound CA IV to different cells and subcellular structures. Anti-sera against CA IV were used in various techniques to immunostain sections of rat and human heart for visualization of CA IV at light microscopic and electron microscopic levels. Furthermore, in situ hybridization (ISH) histochemistry was applied to determine the presence of CA IV mRNA. This technique was carried out on human heart muscle sections and, in view of the immunohistochemical results for CA IV in skeletal muscle mentioned above, also on human skeletal muscle sections.
Materials and Methods
Tissue Preparation
Pieces of rat and human heart muscle were dissected and frozen in liquid nitrogen. Human heart muscle samples were obtained from explanted hearts of patients undergoing transplantation. The samples were taken from unaffected tissue portions immediately after explantation. Cryosections were screened to exclude tissue specimens with visible degradation. Other sections 5-7μm thick intended for further processing were fixed and stored as decribed by Sender et al. (1994).
In addition, pieces of heart muscle were fixed for 4–6 hr in one of the following fixatives: (a) 2.5% glutaraldehyde/2% paraformaldehyde in 0.1 M sodium cacodylate-HCl buffer, pH 7.3; (b) 0.1% glutaraldehyde/4% paraformaldehyde in the same buffer; (c) 3% glutaraldehyde in the same buffer; or (d) paraformaldehyde/lysine/periodate (PLP) in 0.1 M phosphate buffer as described by Yu (1993). For semithin and ultrathin sections, the tissue specimens were postfixed for 1 hr with 1% OsO4 and 1.5% K4[Fe(CN)6] dissolved in 0.1 M sodium cacodylate-HCl buffer, pH 7.3 (Karnovsky 1971). After dehydration the samples were embedded in Epon 812 (Serva; Heidelberg, Germany) or in LR White resin (LRW), medium grade (Plano; Marburg, Germany). Some of the heart muscle specimens that were fixed as described in d were embedded in paraffin.
Immunohistochemistry
Preparation and specificity of the applied antisera, anti-rat lung CA IV and anti-human lung CA IV, were previously described (Brown et al. 1990; Zhu and Sly 1990).
Light Microscopy
The cryosections were stained by immunofluorescence according to Sender et al. (1994). Paraffin sections 2–3μm thick were mounted on poly-
Electron Microscopy
Ultrathin sections about 80 nm thick were etched with a saturated solution of sodium metaperiodate for 30 min. They were incubated overnight at 4C with a solution of 0.1% BSA/5% goat nonimmune serum/0.05 M Tris-HCl, pH 7.6, and anti-CA IV (diluted 1:200-1:500). Sections were further treated according to the protocol given by Decker et al. (1996). The size of the immunogold particles was 10–15 nm. Control sections were incubated either with rabbit non-immune serum (Sigma), with the same dilutions as for the antisera, or with antigen-blocked antisera according to Sender et al. (1994).
Generation of Single-stranded Riboprobes
Full-length single-stranded riboprobes (sense and anti-sense) were synthesized from full-length 1105-base pair (
In Situ Hybridization
Human skeletal muscle samples were frozen in liquid nitrogen. Cryosections 10μm thick were mounted on TESPA-coated slides (Sigma). The sections were fixed in 4% paraformaldehyde in PBS for 20 min. They were then incubated with 25 μg/ml pronase E (Sigma) in 50 mM Tris/5mM EDTA, pH 7.4, for 10 min at 37C. The sections were refixed in 4% paraformaldehyde in PBS for 10 min at room temperature, washed in PBS, and dehydrated in 70% ethanol. After prehybridization at 37C for 2 hr, the sections were hybridized overnight at 37C with the [35S]-UTP-labeled human CA IV riboprobes. Experimental tissue sections were incubated with the anti-sense CA IV riboprobe and control sections with the sense riboprobe. After stringent washing (Kelly et al. 1991), the sections were dehydrated, air-dried, and coated with Ilford K5 photographic emulsion for autoradiography. After exposure for 18 hr the slides were developed and fixed. Counterstaining was performed with hematoxylin/eosin.
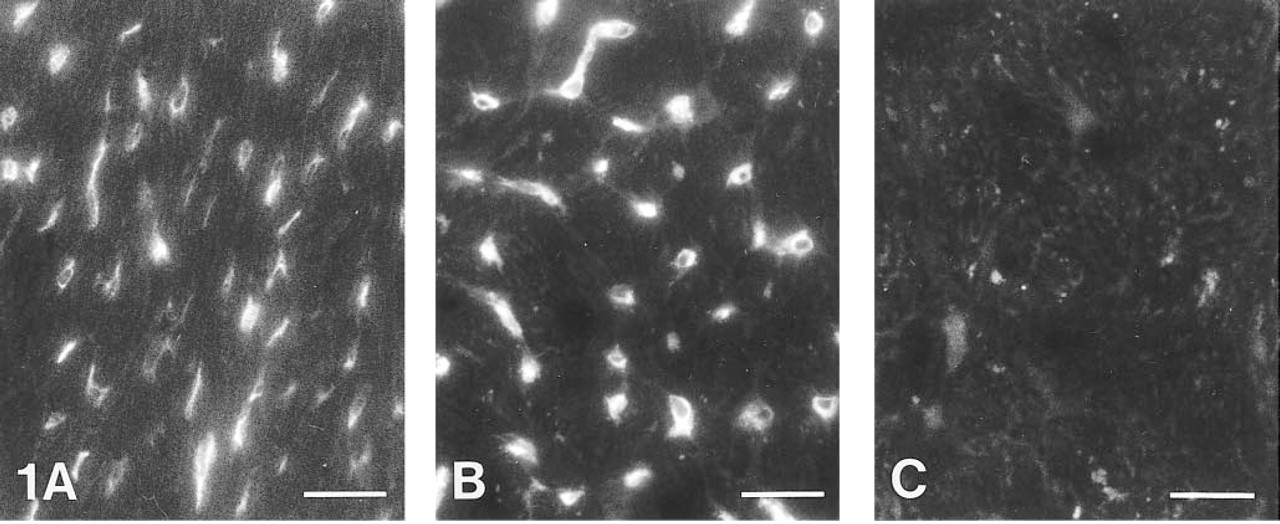
Cryosections from rat heart
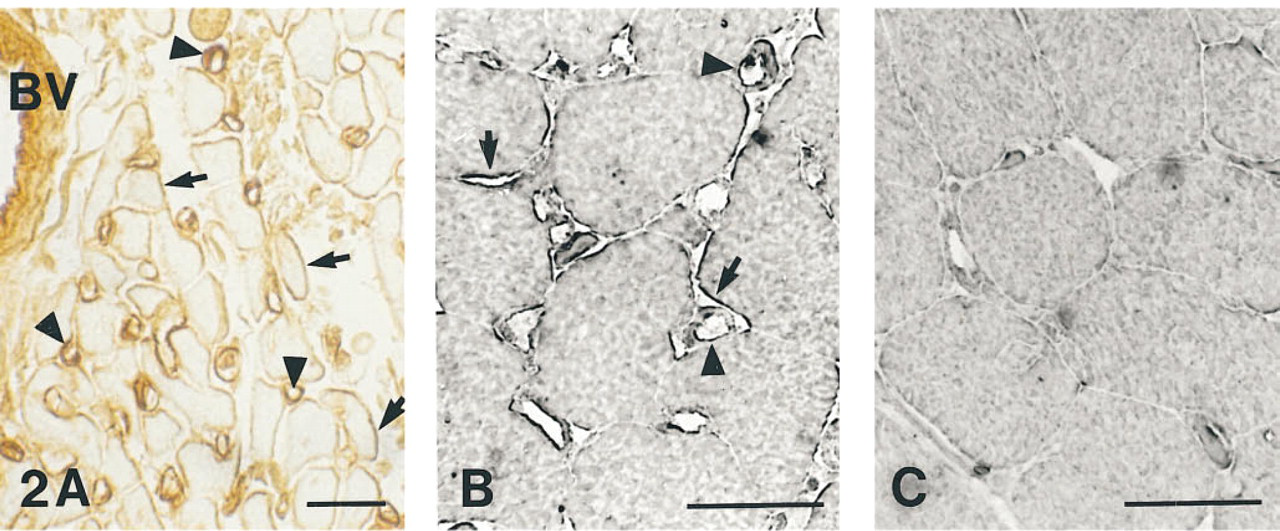
(A) Paraffin section of PLP-fixed rat heart, immunostained with anti-CA IV/avidin-biotin-peroxidase complex. Capillaries (arrow-heads), SL (arrows), and the endothelium of blood vessels (BV) are positively stained.
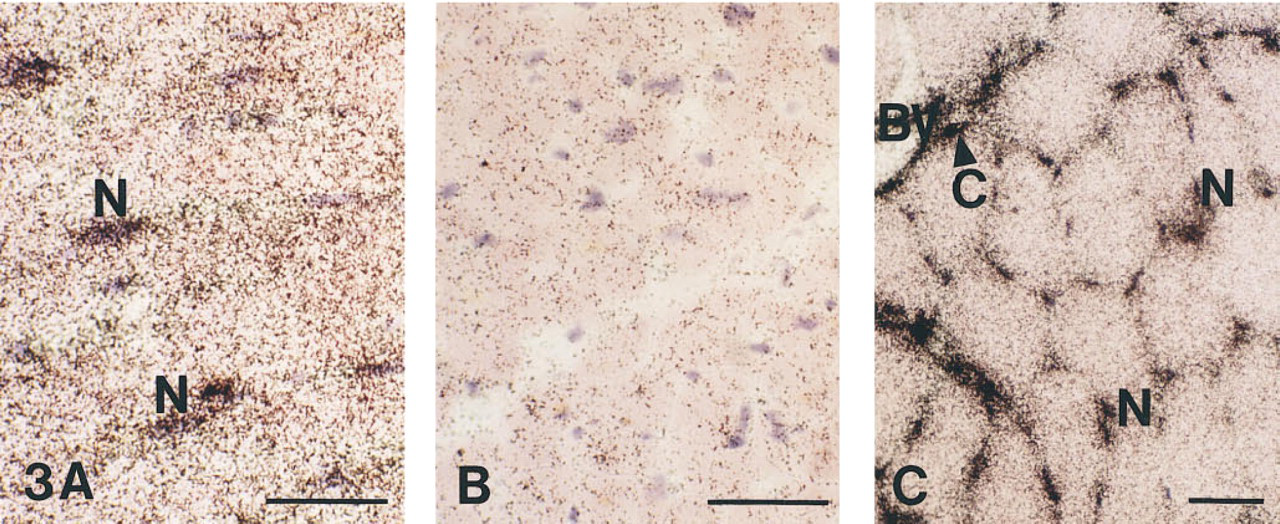
In situ hybridization for CA IV mRNA in cryosections from human heart
Results
Light Microscopy
Immunofluorescence Method: CA IV in Capillary Endothelium. Incubation of heart muscle cryosections with anti-CA IV/FITC resulted in positive staining of the capillary endothelium in rat (Figure 1A) and human heart (Figure 1B). The muscle cells showed no apparent staining. Control sections incubated with non-immune serum/FITC showed only background fluorescence (Figure 1C). Therefore, the immunostaining indicates the association of CA IV with heart capillaries.
Avidin-Biotin-Peroxidase Complex Method: CA IV in Vascular Endothelium and Sarcolemma. After incubation with anti-CA IV/ABC, paraffin sections of rat heart showed intense brown staining in the capillary endothelium and the endothelium of blood vessels (Figure 2A). A positive reaction was also seen at the SL, which exhibited a brown color of variable intensity. The cytoplasm in only some of the cardiac muscle cells showed yellowish staining, which was also seen in control sections (not shown). We conclude, therefore, that CA IV is present in the vascular endothelium and in the SL. This result is confirmed by semithin sections of rat cardiac muscle incubated with anti-CA IV/ABC or nonimmune serum/ABC, respectively. There was strong SL and capillary staining in parts of the antiserum-treated sections (Figure 2B). With this technique, the capillary staining appeared not only on the luminal but also on the external surface of the capillary wall. As in the cryosections (Figures 1A and 1B) and in the ABC-treated paraffin sections (Figure 2A), the cytoplasm showed no specific staining. In the control sections, almost no immunoreactivity was visible throughout the sections (Figure 2C).
In Situ Hybridization to CA IV-specific mRNA. The hybridization of isotope-labeled riboprobe with tissue intrinsic CA IV mRNA transcripts was shown in heart (Figure 3A) and skeletal muscle (Figure 3C). In cardiac muscle sections, an intense hybridization signal was seen in many nuclei and, to a lesser extent in the cytoplasm. In control sections (Figure 3B) in which a sense RNA probe was used, the amount of silver grains was significantly reduced. On the basis of the number and distribution of the positive nuclei, one can conclude that both cardiomyocytes and capillary endothelial cells express CA IV. In addition, in both heart and skeletal muscle sections there was a prominent signal in larger blood vessels. This is shown in Figure 3C for skeletal muscle, together with a high signal intensity over nuclei and a lower signal intensity in the cytoplasm. Because the nuclei of skeletal muscle cells are often close to capillaries, it is sometimes difficult to unequivocally attribute the signal to one of these cell types. However, some capillaries showing accumulation of siver grains can definitely be identified. Therefore, we conclude that there may be an identical pattern of CA IV mRNA expression in heart and skeletal muscle.
Electron Microscopy
The applied fixatives and embedding media gave similar results regarding intensity and localization of the CA IV immunoreaction. Figure 4F shows a section from human heart; the other sections were taken from rat cardiac muscle.
CA IV in Capillary Endothelium. After incubation with anti-CA IV/immunogold, capillary endothelial cells showed an accumulation of gold particles, predominantly in the proximity of the luminal membrane (Figures 4A and 4B).
CA IV in the Sarcolemma. The SL, too, exhibited strong immunoreaction for CA IV (Figure 4C). Furthermore, CA IV appeared to be associated with specialized sarcolemmal structures, intercalated discs and T-tubules. As shown in Figures 4D and 4F, patches of gold particles were found next to the intercalated discs. Very few particles were observed within the cells. An accumulation of gold particles could also be seen associated with membranous structures close to the Z discs (Figure 4E). We interpret this staining pattern as a positive reaction of T-tubules, which extend inward from the cell surface at this site, and/or as staining of junctional SR.
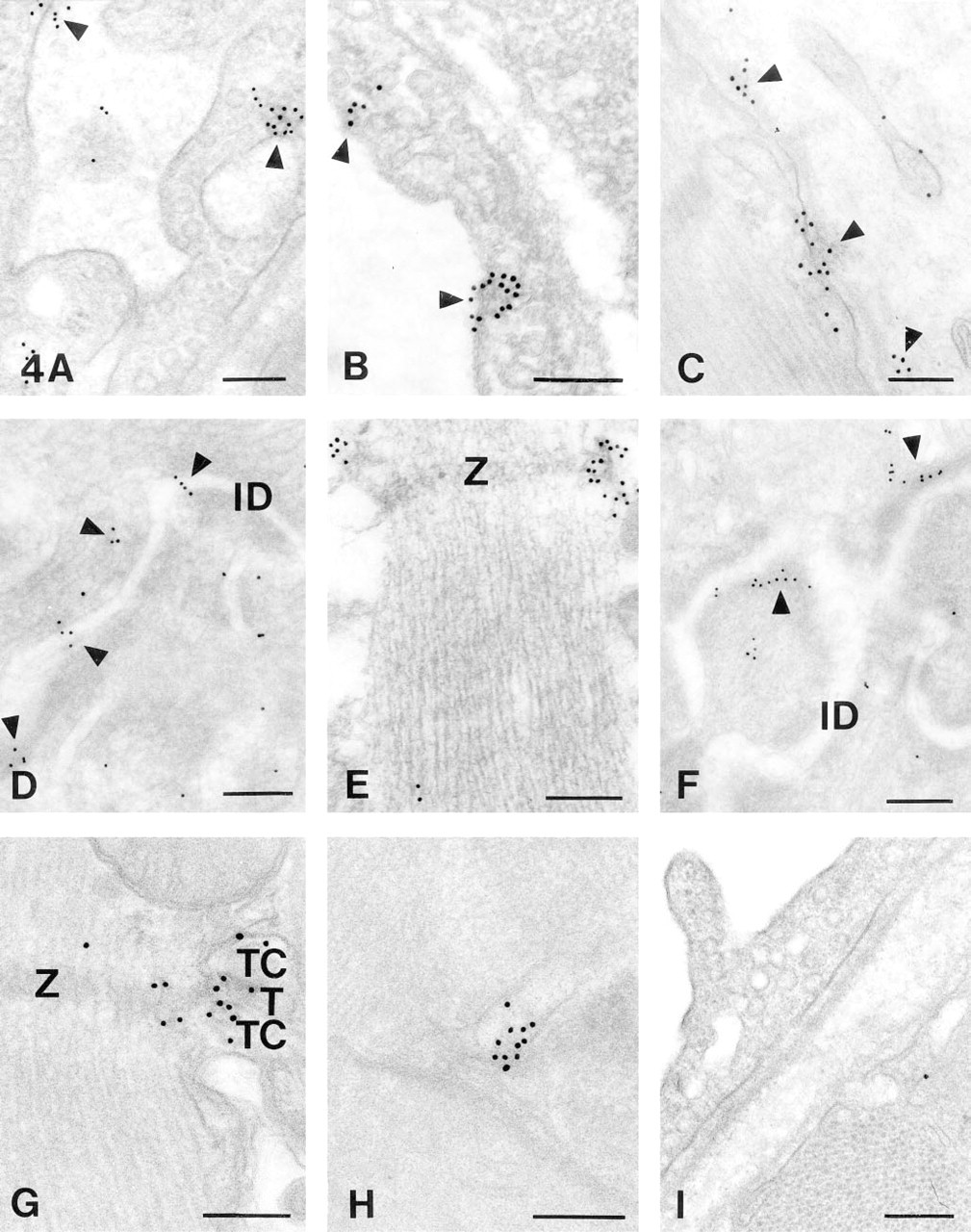
Ultrathin sections of heart tissue fixed with 2.5% glutaraldehyde/2% paraformaldehyde and embedded in Epon
CA IV in Sarcoplasmic Reticulum. CA IV inside the muscle cells themselves is shown in Figures 4G and 4H. Clusters of gold particles were associated with junctional complexes between sarcolemma and SR membrane, situated at the level of Z discs. In Figure 4G, both junctional SR and T-tubules are positively stained, whereas mitochondria remain unstained. Figure 4G shows an example of a triadic junctional complex, although the majority of junctional complexes in heart are diadic. What are called “terminal cisternae” in Figure 4G are much smaller in heart than in skeletal muscle and may not be wider than the L-system in general. In the longitudinal system, seen to be located between the myofibrils, we also found CA IV (Figure 4H). Control sections incubated with antigen-blocked antiserum/immunogold showed almost no background staining throughout the whole section (Figure 4I).
The ultrathin sections therefore support and extend the light microscopic results mentioned above. CA IV is located on the luminal membrane of capillary endothelial cells and is associated with SL and SR of cardiac muscle cells.
Discussion
CA IV in Capillary Endothelium
The localization of CA IV in capillary endothelium of heart muscle is consistent with our earlier demonstration of CA IV in skeletal muscle capillaries (Sender et al. 1994; Decker et al. 1996). As in skeletal muscle, electron microscopy shows CA IV to be associated with the endothelial luminal membrane both in human and rat heart. Using the modified Hansson technique, Vandenberg et al. (1996) have shown CA to be located in capillary endothelium of ferret heart. In their sections, as in our semithin sections (Figure 2B), staining for CA activity was apparent both at the luminal and the external capillary surface. The modified Hansson technique does not allow the identification of the specific CA isozyme involved. From Western blots, showing a reaction of crude ferret heart homogenates with anti-bovine CA IV, the authors concluded that the CA, stained by the Hansson technique, is likely to be CA IV.
Concerning the physiological role of this CA, Forster and Crandall (1975) have shown that a capillary CA can accelerate CO2 uptake by the blood during capillary transit. The significance of endothelial CA IV in larger blood vessels remains to be ascertained.
CA IV in the Sarcolemma
As in skeletal muscle (Sender et al. 1994; Decker et al. 1996), CA IV could be detected in heart muscle SL in semithin and ultrathin sections but not in cryosections. On the one hand, the higher sensitivity of the ABC technique compared with the immunofluorescence method may contribute to this finding. On the other hand, as discussed for skeletal muscle sections, this probably also indicates a problem of accessibility (Decker et al. 1996). As can be seen in Figures 2A and 2B, SL staining becomes apparent in locations at which the antibodies have easiest access to the outer surface of the SL. This access may be facilitated by the ABC technique, which uses fixation media that lead to considerable cell shrinkage and hence to a widening of the intercellular spaces. The present immunohistochemical results showing CA IV associated with the SL and with specific sarcolemmal structures, the intercalated discs and the T-tubules, agree with the histochemical localization of CA as described by Vandenberg et al. (1996). With the Hansson technique, staining for CA in addition to capillaries was apparent at the SL in longitudinal sections and was most prominent in the region of the intercalated discs. As the authors pointed out, it would require electron microscopy to unequivocally demonstrate that this CA is indeed associated with endothelial as well as sarcolemmal membranes. In the present study we confirm at the electron microscopic level that CA IV is associated with the capillary luminal membrane and with the SL. CA at the latter location may have a function in pH regulation (De Hemptinne et al. 1987; Lagadic-Gossmann et al. 1992; Vandenberg et al. 1996).
CA IV in the Sarcoplasmic Reticulum
In heart muscle, SR CA IV could be demonstrated at the electron microscopic but not at the light microscopic level, a phenomenon that has also been observed in skeletal muscle sections (Sender et al. 1994; Decker et al. 1996). However, in cryosections of rabbit heart, Bruns and Gros (1992) did report an intracellular staining for CA with dansylsulfonamide (DNSA), a fluorescent CA inhibitor. The staining appeared to be associated with intracellular structures and was interpreted as a positive reaction of SR and T-tubules. On the other hand, Vandenberg et al. (1996) found no intracellular staining for CA activity with the modified Hansson method, consistent with our present light microscopic results. As has been discussed with regard to skeletal muscle (Decker et al. 1996), a problem of accessibility of CA IV to the histochemical reagent appears to exist. Supposing that CA is mainly oriented towards the SR interior, as was shown for skeletal muscle (Geers et al. 1991; Gros et al. 1993), only lipo-philic substances such as DNSA, but not macromolecules, such as antibodies, may be able to penetrate the SR membranes and react with an intra-SR enzyme epitope. Only in ultrathin sections of 80 nm is the section thickness less than the thickness of longitudinal tubules, so that free access of antibodies to the intra-SR CA is expected. In all thicker sections, a major part of the SR may be present as an intact, closed membranous network into which antibodies cannot enter. The same problem may exist for the reagents used in the Hansson technique, which may not gain access to the active center of the enzyme inside the SR.
It has been postulated that CA in skeletal muscle SR is necessary to rapidly produce and buffer H+ that move across the SR membrane in exchange for Ca++ (Wetzel et al. 1990). CA IV in heart muscle SR may play an anologous role, as has been indicated by results of Geers and Gros (1995).
CA IV mRNA Expression in Endothelium and Muscle Cells
The ISH histochemistry clearly shows that CA IV mRNA is present in heart muscle and skeletal muscle cells as well as in capillaries and other vessels. This is in accordance with the immunohistochemical results presented above, indicating the synthesis of CA IV in the same cell types that exhibit CA IV immunoreactivity.
The percentage similarities between human CA IV and other human CAs present in skeletal muscle cells and/or erythrocytes are 31% for CA I, 36% for CA II, and 30% for CA III at the amino acid level (Okuyama et al. 1992). Considering that the percentage homology at the nucleotide level will be even lower, we believe that nonspecific hybridization of the riboprobe to mRNAs of other CA isozymes is eliminated by the use of stringent washing procedures.
Footnotes
Acknowledgements
Supported by the Deutsche Forschungsgemeinschaft, grant Gr 489/10.
The excellent technical assistance of Iris Albers and Elke Mallon and the expert photographic assistance of Angelika Hundt, are gratefully acknowledged.