
Abstract
We investigated the localization of polysialic acid (PSA), neural cell adhesion molecule (NCAM), and vesicular acetylcholine transporter (VAChT) in adult rat retina by using immunofluorescence with a confocal laser scanning microscope. Western blot analysis showed a typical broad smear of PSA and isoforms of NCAM (120, 140, and 180 kD). PSA immunofluorescence revealed multistratification in the inner plexiform layer (IPL). Dual immunostaining for PSA and NCAM exhibited the selective co-expression of PSA and NCAM on Müller cells. Moreover, dual immunolabeling for PSA and VAChT completely separated the five strata in the IPL. Strata 1, 3, and 5 were immunoreactive for PSA and Strata 2 and 4 for VAChT. These results suggest the possibility that PSA molecules on Müller cells are spatially related to ON and OFF retinal channels in the IPL.
Keywords
P
All vertebrate retinas consist of a neuronal network organized according to the same basic plan: three nuclear layers and two intercalated plexiform (synaptic) layers. The structural and neurochemical diversity at the level of the inner plexiform layer (IPL) is particularly perplexing and as yet not well understood. Cajal (1893) subdivided the IPL into five strata of equal thickness, with Stratum 1 beginning under the inner nuclear layer (INL) and Stratum 5 ending on the ganglion cell layer (GCL). Nelson et al. (1978) showed, by intracellular recordings and dye injections, that OFF-center ganglion cells branch closer to the INL, whereas ON-center cells branch closer to the GCL. Thus, in some cases the IPL can be subdivided into an OFF layer (Strata 1 and 2) and an ON layer (Strata 3, 4, and 5) (Famiglietti and Kolb 1976). In mammals, amacrine cells have been classified into 27 types on the basis of their dendritic field sizes and their differing stratification levels in the IPL (Kolb et al. 1992). In addition, a variety of neurotransmitters are found in the vertebrate retina, a great number of which are located in different amacrine cell types (for review see Brecha 1983). One of these transmitters is acetylcholine (Masland and Mills 1979). To date, these cholinergic amacrine cells, called “starburst” amacrine cells from the appearance of their dendrites, are the most extensively investigated of all amacrine cells. It is well established that cholinergic amacrine cells occur in mirror-symmetric pairs: the type with a normally placed cell body in the INL branches in Stratum 2, whereas the other, which has a displaced cell body in the GCL, branches typically in Stratum 4 (Famiglietti 1983; Kolb et al. 1992). Recently, vesicular acetylcholine transporter (VAChT) protein was demonstrated as a novel and unique marker for cholinergic neurons in the central and peripheral nervous systems (Gilmor et al. 1996; Arvidsson et al. 1997; Koulen 1997).
Müller cells are radial glial cells in the retina, which extend from the external to the internal limiting membrane. They are characterized by extensive cytoplasmic expansions, which fill all intercellular spaces and envelop the cell bodies of the neurons. As the principal glial cells, they conserve the structural alignment of its neuronal elements (Bron et al. 1997). Bartsch et al. (1990) reported the continuous expression of PSA on only Müller cells in the adult mouse retina and described three bands of PSA immunoreactivity in the IPL.
In this study we have investigated the expression of PSA in the adult rat retina using a monoclonal antibody against PSA. Immunofluorescence for PSA confirms the distinctive expression of PSA on Müller cells and demonstrates the multistratification in the IPL. To identify the PSA-immunoreactive strata, we have employed the dual immunostaining technique with a confocal laser scanning microscope. Dual staining for PSA and NCAM shows selective co-localization of PSA and NCAM on Müller cells. Interestingly, the five strata comprising the IPL are very clearly discernible by dual immunostaining for PSA and VAChT.
Materials and Methods
Antibodies
The mouse IgG monoclonal antibody (designated MAb 735) was a kind gift from Dr. D. Bitter-Suermann (Institute of Medical Microbiology, School of Medicine; Hannover, Germany) (Frosch et al. 1985). It recognizes homopolymers of α-2,8-linked polysialic acid at least more than eight residues in length (Roth et al. 1987). The polyclonal antibody against rat NCAM (120, 140, and 180 kD) was purchased from Chemicon International (Temecula, CA). The goat antiserum against the synthetic peptide CSPPGPFDGCEDDYNYYSRS, corresponding to amino acids 511-530 of the rat VAChT, was also purchased from Chemicon.
Immunohistochemistry for PSA
Male Wistar rats, 8-10 weeks old, were deeply anesthetized with diethylether and then perfused with 3% paraformaldehyde-0.1% glutaraldehyde in PBS. The eyes were removed and further fixed by immersion in the above fixative at 4C overnight. After washing in PBS for 2 hr, the eyes were cryoprotected in 15% sucrose in PBS for 2 hr and 30% sucrose in PBS at 4C overnight. They were then embedded in OCT compound (Miles; Elkhart, IN) and then cut into 12-μm sections with a cryostat. After several rinses in PBS, the sections were incubated with 50 mM NH4Cl in PBS for 30 min to block free aldehyde groups. After washing in PBS, the sections were treated with 5% normal goat serum (NGS) and 1% bovine serum albumin (BSA) in PBS for 10 min and then incubated with MAb 735 (diluted 1:2000 with 1% BSA in PBS) at 4C overnight. After washing in PBS, the sections were incubated with fluorescein isothiocyanate (FITC) conjugated to horse anti-mouse IgG (Vector Laboratories; Burlingame, CA; diluted 1:200 with 1% BSA in PBS) for 2 hr. After washing in PBS, the sections were briefly rinsed with water and mounted in Gel/mount (Biomeda; Foster City, CA). Immunofluorescence labelings were examined with a confocal laser scanning microscope (TCS4D; Leica, Welzlar, Germany). As controls, the primary antibody was omitted from the procedure or replaced by normal mouse IgG.
For immunoelectron microscopy using the pre-embedding staining procedure, vibratome sections (90 μm thick) were incubated with 5% NGS and 1% BSA in PBS for 30 min to block nonspecific binding and were then incubated with MAb 735 (1:2000) for 2 days at 4C. After washing in PBS, the sections were incubated with anti-mouse IgG-HRP (MBL, Nagoya, Japan; diluted 1:40) at 4C overnight. After washing in PBS, the sections were fixed with 1.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (CB, pH 7.4), containing 5% sucrose for 1 hr. After washing in 50 mM Tris-HCl buffer (TB, pH 7.4), the sections were incubated in 0.05% 3′,3′-diaminobenzidine (DAB) in TB for 20 min, then in the complete DAB solution containing 0.005% H2O2 for 10 min. The sections were postfixed with 1% OsO4 in CB containing 1% potassium ferrocyanide for 1 hr, dehydrated with a graded series of ethanol, and then embedded in epoxy resin mixture. Ultrathin sections were contrasted with lead citrate for 20 sec and examined with a JEOL 1200 EX electron microscope (JEOL; Tokyo, Japan).
Enzyme Digestion with Neuraminidase
To confirm the specificity of MAb 735, the sections were treated with neuraminidase to remove the sialic acid residues. After briefly rinsing in PBS, the cryostat sections were incubated with 0.1 M acetate buffer, pH 5.3, for 5 min at room temperature (RT), and then were digested with highly purified neuraminidase from Arthrobacter ureafaciens (Nacalai Tesque, Kyoto, Japan; 0.5 U/ml in 0.1 M acetate buffer, pH 5.3) at 37C overnight. After washing in PBS, the sections were stained with MAb 735 as described above.
Western Blot Analysis
Immunoblotting was done essentially as described by Bartsch et al. (1989). Briefly, retinas were homogenized in 25 mM Tris, pH 7.4, 1 mM CaCl2, 1 mM MgCl2, and protease inhibitors. Nuclei were pelleted at 600 X g. The supernatant was mixed with SDS sulfate sample buffer and boiled at 65C for 10 min. The sample (6 μg of protein) was loaded into each lane of a 5% stacking gel and an 8% separation gel. Separated proteins were transferred to polyvinylidene difluoride (PVDF) membranes (Millipore; Bedford, MA). After incubation with 10% NGS and 0.05% Tween-20 (v/v)/Trisbuffered saline (TBS) for 20 min to block nonspecific binding sites, the PVDF strips were incubated with MAb 735 diluted 1:2000 or anti-NCAM diluted 1:4000 in 1% BSA/0.05% Tween-20/TBS for 60 min. After washing, goat anti-mouse IgG-HRP (MBL, Nagoya, Japan; diluted 1:1000) or goat anti-rabbit IgG-HRP (Bio-Rad Laboratories, Hercules, CA; diluted 1:10000) was added for 60 min. The thoroughly rinsed blots were developed with an enhanced chemiluminescence system (Du Pont; Boston, MA).
Dual Immunostaining for PSA and NCAM
The sections were incubated with MAb 735 (1:1000) mixed with antibody against NCAM (1:1000) at 4C overnight. After washing in PBS, the sections were incubated with a mixture of FITC-conjugated horse anti-mouse IgG (Vector; diluted 1:100 with 1% BSA in PBS) and Texas Red-conjugated goat anti-rabbit IgG (Southern Biotechnology Associates, Birmingham, AL; diluted 1:50 with 1% BSA in PBS) for 2 hr at RT. After washing in PBS, the sections were briefly rinsed with water and mounted in Gel/mount. Dual immunofluorescence labelings were simultaneously scanned by a confocal laser scanning microscope.
Dual Immunostaining for PSA and VAChT
The sections were preincubated with 5% normal horse serum in PBS and then incubated with the mixture of MAb 735 (1:1000) and antiserum to rat VAChT (1:1000) at 4C overnight. After washing in PBS, the sections were incubated with a mixture of FITC-conjugated horse anti-mouse IgG (Vector; diluted 1:100 with 1% BSA in PBS) and biotinylated horse anti-goat IgG (Vector; diluted 1:100 with 1%
BSA in PBS) for 2 hr at RT. After washing in PBS, the sections were incubated with streptavidin-Texas Red conjugate (Bethesda Research Laboratories, Gaithersburg, MD; diluted 1:200 with 1% BSA in PBS) for 30 min at RT. After washing in PBS, the sections were briefly rinsed with water and mounted in Gel/mount. Dual immunofluorescence labelings were simultaneously scanned by a confocal laser scanning microscope.
Results
Light Microscopy for PSA Immunostaining
In the adult rat retina, immunoreactivity for MAb 735 was observed throughout the entire thickness of the neural retina, from the inner limiting membranes to the outer limiting membranes (Figures 1A and 1B). In particular, dense labeling occurred in the outer plexiform layer (OPL) and the IPL. In the latter, MAb 735 clearly demonstrated multistratified structures (Figure 1B. The staining pattern was composed of the dense network of fine processes in the IPL and in the OPL, the decoration of the cell bodies in the INL and in the ONL, and the outer and inner limiting membranes, probably representing microvilli and end-feet of Müller cells, respectively. In addition to the neural retina, MAb 735 immunoreactivity was also detected in the choroid, particularly in the larger vessels and loose connective tissue (Figure 1A. Intense labeling was concentrated in the perivascular regions. It appears that MAb 735 labels the axons of the vasomotor nerves in the choroidal stroma. As controls, omission of MAb 735 or normal mouse IgG instead of the primary antibody revealed no immunofluorescence (not shown).
Enzyme Digestion with Neuraminidase
To confirm the specificity of MAb 735, tissue sections were preincubated with neuraminidase. The extensive enzyme digestion completely abolished MAb 735 immunoreactivity (Figure 1C.
Western Blot Analysis
Western blots of the adult rat retina with MAb 735 revealed a broad smear from 150 to 260 kD (Figure 2, Lane 1). Such a typical broad smear and the molecular mass range corresponded to the profile of PSA reported previously. Immunoreactivity with anti-NCAM appeared as discrete bands at 120, 140, and 180 kD. (Figure 2, Lane 2).
Immunoelectron Microscopy
At the external limiting membrane portion of the retina, immunoreactivity was found only on Müller cell processes and their microvilli, which projected into the space between photoreceptor cell processes (Figure 3A. At the internal limiting membrane portion, immunoreactivity was found along the cell membrane of Müller cell processes and their expanded end-feet (Figure 3B. No immunoreactivity was found in any other neuronal elements in the retina. On the basis of these findings, we confirmed that Müller cells exhibited PSA immunoreactivity in the adult rat retina. In the choroid, strong immunoreactivity was also observed on the surface of unmyelinated nerve fibers (not shown). As control, normal mouse IgG instead of MAb 735 did not reveal any specific DAB reaction products.
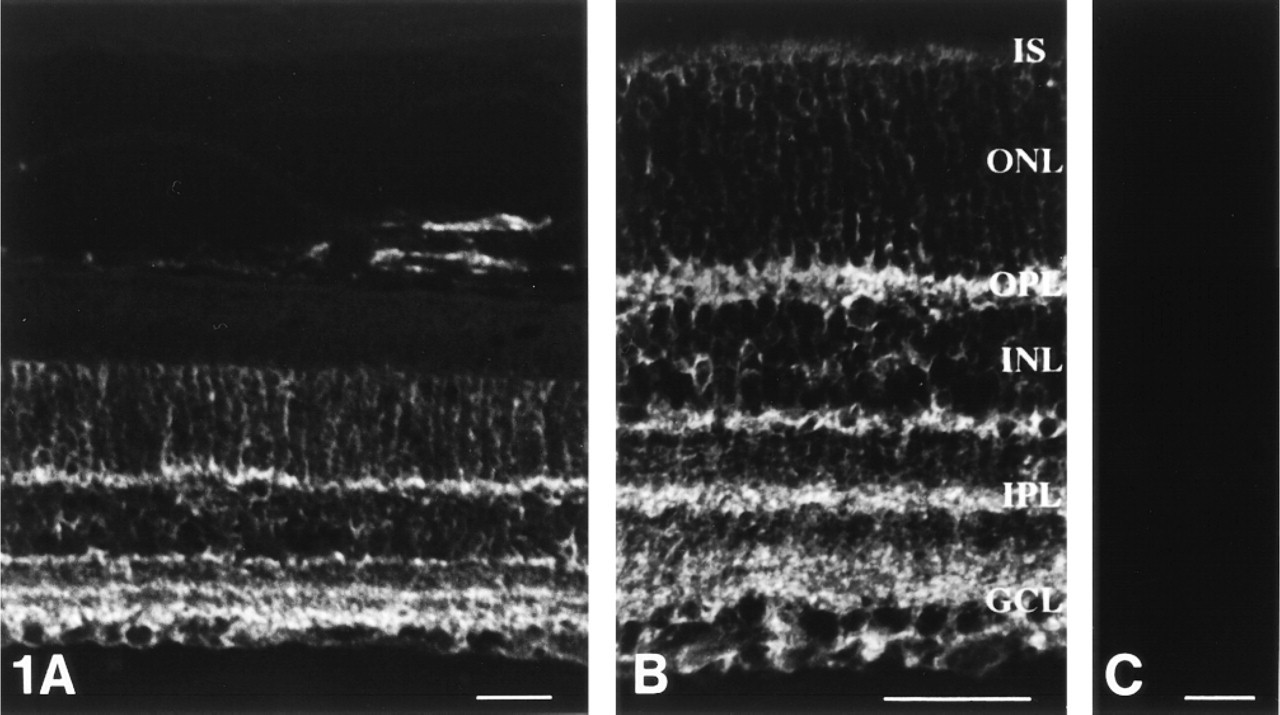
Representative single confocal tomographs of immunofluorescence of PSA in the adult rat retina and the choroid. (
Dual Immunostaining for PSA and NCAM
PSA immunoreactivity demonstrated the multistratified structures in the IPL (Figure 4A. In contrast, NCAM immunofluorescence showed diffuse labeling of the retina without stratification in the IPL (Figure 4B. To examine the co-localization of PSA and NCAM, we performed dual immunostaining for PSA and NCAM. Three yellow stratifications could be distinguished in the IPL (Figure 4C. These results indicated the selective expression of PSA in the adult rat retina, especially in the IPL.
Dual Immunostaining for PSA and VAChT
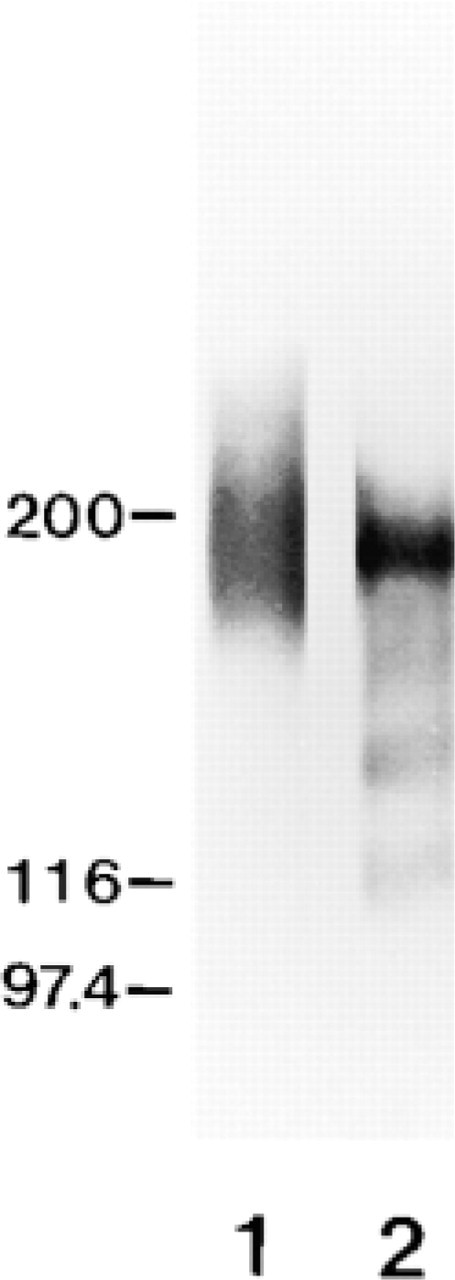
Western blot analysis of PSA and NCAM in the adult rat retina. Lane 1 is immunostained for PSA (with MAb 735) and Lane 2 for NCAM (with polyclonal antibody against rat NCAM). Positions of molecular weight markers are indicated at left.
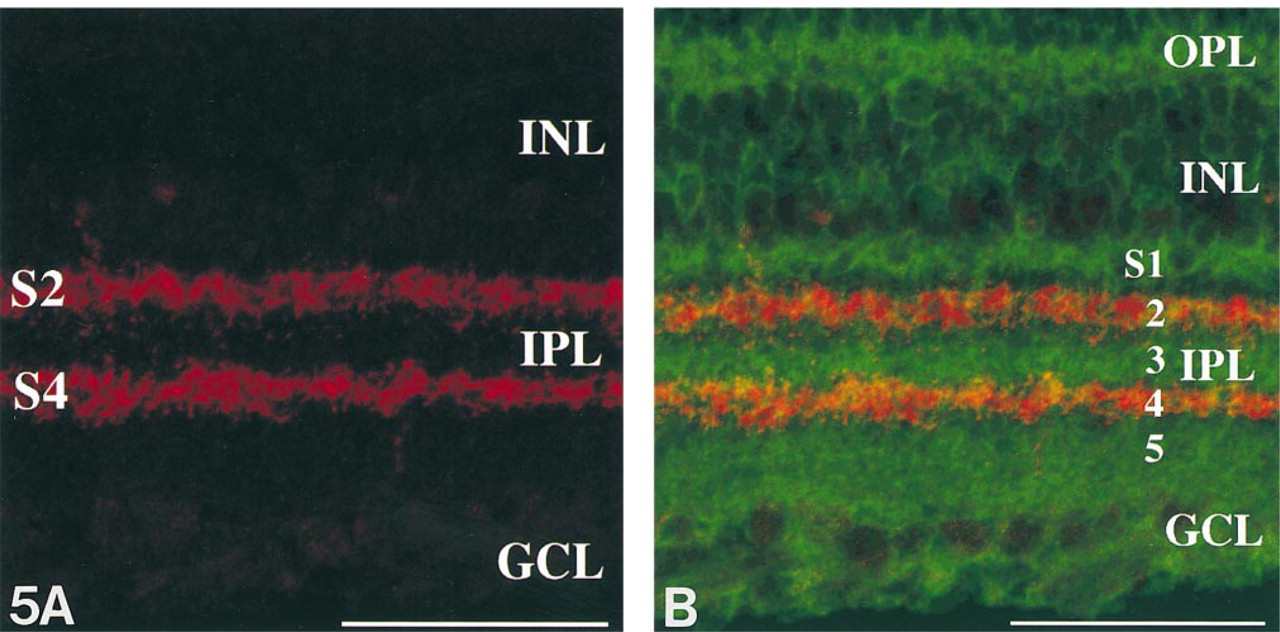
Immunostaining for VAChT, which has been recently reported as a novel marker for cholinergic neurons, demonstrated two prominent strata corresponding to Strata 2 and 4 in the IPL (Figure 5A. To identify which strata were immunoreactive for PSA, we performed dual immunostaining for PSA and VAChT. Figure 5B clearly demonstrates that the immunoreactive strata for PSA are completely distinguished from those for VAChT. It indicates that the PSA-immunoreactive multistrata were Strata 1, 3, and 5.
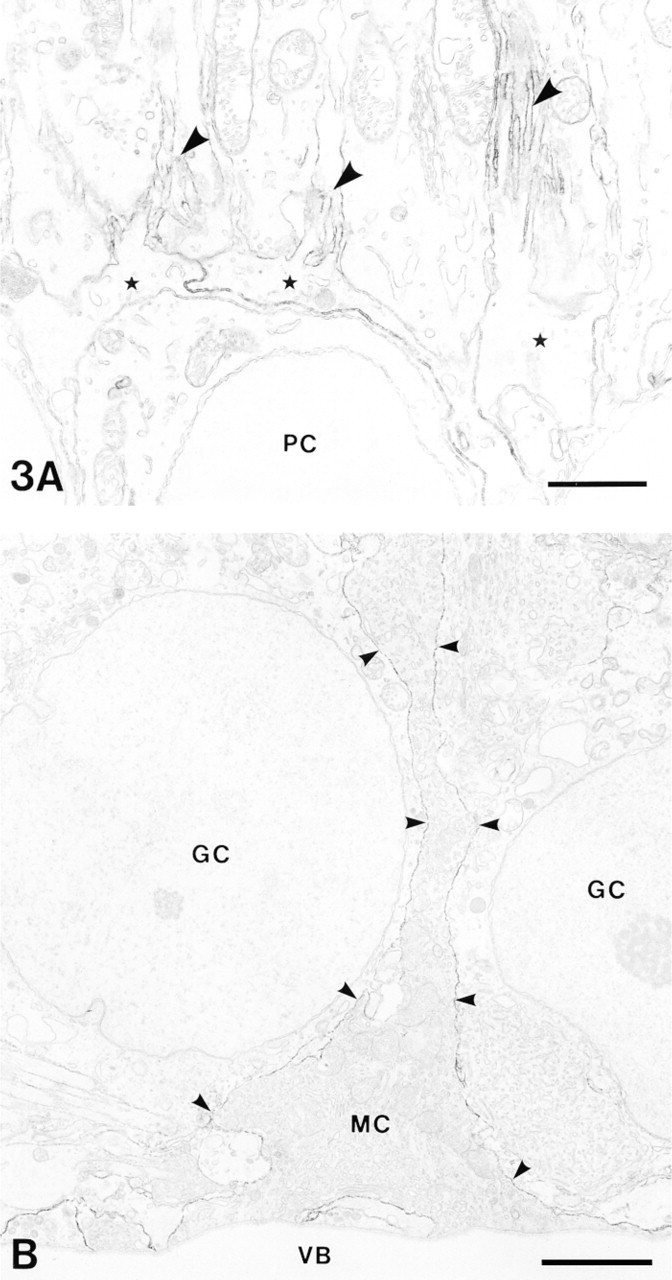
Immunoelectron microscopic localization of PSA in the adult rat retina. (
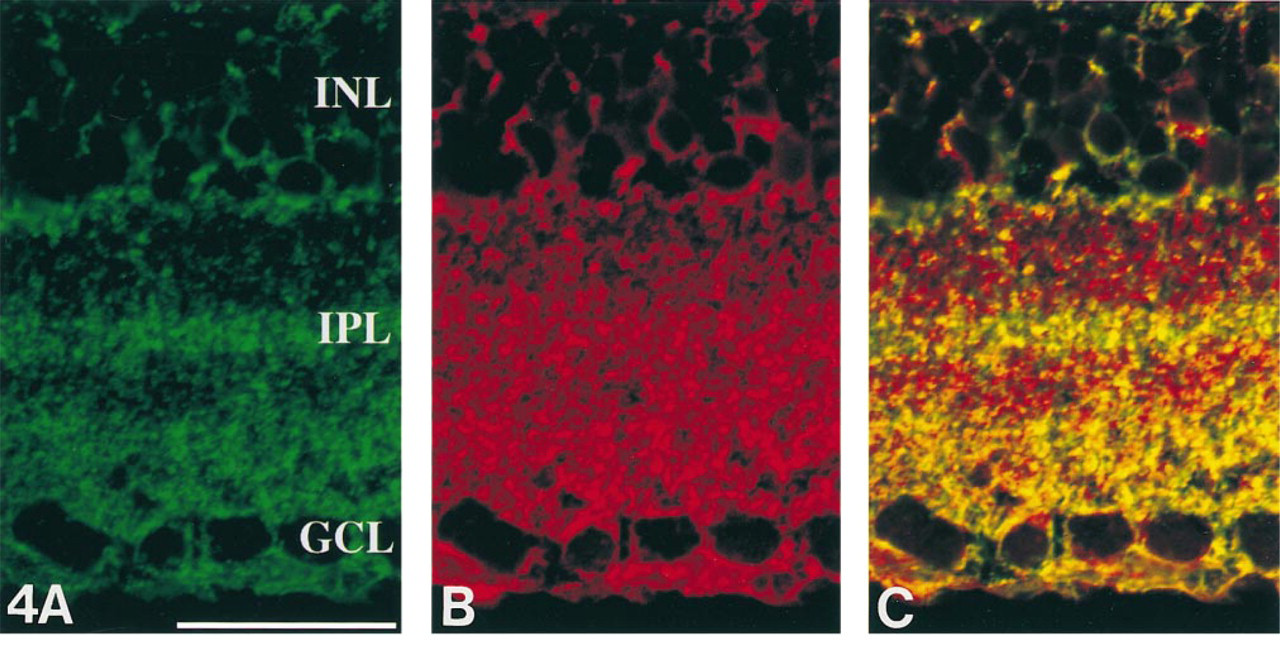
Representative single confocal images simultaneously scanned by a confocal laser scanning microscope. High magnification of the IPL stained for PSA and NCAM as shown by indirect immunofluorescence. (
(
Discussion
We have demonstrated the expression of PSA in the adult rat retina and choroid by using MAb 735 immunostaining. The pattern of the immunostaining is quite compatible with that of the adult mouse retina as reported previously (Bartsch et al. 1990). We have emphasized the multistratified expression of PSA in the IPL. Dual immunostaining for PSA and VAChT clearly separated the IPL into five strata, which presumably correspond with those of the IPL described by Cajal (1893). Strata 2 and 4 were immunoreactive for VAChT, and Strata 1, 3, and 5 for PSA. To the best of our knowledge, this is the first report that the five strata of the IPL can be so clearly discernible with dual immunostaining for PSA and VAChT.
Bartsch et al. (1990) first showed that Müller cells of the adult mouse retina continue to express PSA molecules, using immunohistochemistry at the light and electron microscopic levels. In the salamander retina, Pleurodeles waltl, only Müller cells continue to express PSA into adulthood (Becker et al. 1993). Interestingly, the rat brain L-glutamate/L-aspartate transporter 1 (GLAST-1) (Rauen et al. 1996) and the aquaporin-4 water channel protein (AQP4) (Nagelhus et al. 1998) are specifically expressed in Müller cells and fibrous astrocytes in the rat retina. This immunostaining pattern, especially the stratification of the IPL, is quite similar to that of PSA immunofluorescence.
Western blots analysis of the adult rat retina exhibited three major isoforms of NCAM (120, 140, and 180 kD), which are in agreement with previous reports (Lackie et al. 1991; Goridis and Brunet 1992; Zuber et al. 1992). NCAM, first reported by Brackenbury et al. (1977), is an abundant integral membrane glycoprotein that can promote cell-to-cell adhesion through a homophilic binding mechanism (Rutishauser et al. 1988; Doherty et al. 1990). In the mouse retina, NCAM-180 and NCAM-140 are present during development, whereas in the adult retina only NCAM-180 and NCAM-120 are expressed (Bartsch et al. 1989). In the goldfish retina, NCAM-180 is present during development but not in the adult retina (Bastmeyer et al. 1990). These isoform differences in species may be further elucidated elsewhere.
Dual immunostaining for PSA and NCAM revealed the selective co-localization of PSA and NCAM in the adult rat retina, especially in the IPL. Previously, it has been estimated that the degree of sialylation is less than 10% in the adult (Rothbard et al. 1982; Hoffman and Edelman 1983). Therefore, our results in the expression of PSA and NCAM in the adult closely correspond to previous observations.
VAChT has been recently shown to be a novel and unique marker for cholinergic neurons in the central and peripheral nervous systems (Gilmor et al. 1996; Arvidsson et al. 1997). Koulen (1977) demonstrated that VAChT was useful as a cellular marker in rat retinal development. VAChT immunoreactivity showed multistratification in two prominent bands in Strata 2 and 4 of the IPL of the rat retina, matching the cholinergic amacrine cell bounding patterns; one type has a normally placed cell body in the INL and branches in Stratum 2, and the other has a displaced cell body in the GCL and branches typically in Stratum 4 (Famiglietti, 1983; Kolb et al. 1992). Thus far, the other strata, which have no reactivity with cholinergic neuron markers, are not well described. We have easily identified all five strata by using dual immunostaining techniques and confocal laser scanning microscopy.
Acheson and Rutishauser (1988) have demonstrated that NCAM during development regulates cell contact-mediated changes in choline acetyltransferase (ChAT) activity of embryonic chick sympathetic neurons in vitro, and that the presence of a PSA moiety on NCAM blocks the cell contact-mediated increase in ChAT activity. The high level of PSA expression in the IPL at the peripheries of cholinergic neuron dendrites suggests influence on ChAT activity during development. This might further influence the functional development of strata of the IPL. Some authors suggest that removal of PSA enhances cell interactions (Rutishauser 1992) and leads to an increase in axon-axon fasciculation during development (Tang et al. 1994). Others, however (Cremer et al. 1997), suggest that PSA might contribute positively to fiber fasciculation, and therefore its absence would result in formation of smaller bundles. In the hippocampus, genetic or enzymatic removal of PSA induces aberrant innervation and ectopic synaptogenesis in general (Seki and Rutishauser 1998). We must await further experiments on the relationship of PSA and ChAT in the developing nervous system to understand these findings.
In conclusion, PSA molecules on Müller cells, which continue to be expressed in the adulthood, exhibit multistratification in the IPL. Dual immunostaining for PSA and VAChT clearly identify the five strata in the IPL. Strata 1, 3, and 5 are immunoreactive for PSA, and Strata 2 and 4 for VAChT. These results imply that Müller cell processes are spatially related to ON and OFF retinal channels in the IPL, and that PSA molecules on Müller cells and cholinergic dendritic development may play an important role in the establishment of retinal organization.
Footnotes
Acknowledgements
Supported in part by a Grant-in-Aid for Scientific Research (C) (08670024 to TS) from the Ministry of Education, Science, Sports and Culture, Japan.
We thank Dr D. Bitter-Suermann for providing MAb 735, Dr Hiroaki Kataoka (Department of Pathology, Miyazaki Medical College) for excellent technical advice, Yoshiteru Goto (Division of Electron Microscopy, Central Research Laboratories, Miyazaki Medical College) and Eiko Matsuura (Department of Anatomy, Miyazaki Medical College) for expert assistance.