
Abstract
The desert gerbil Psammomys obesus, an established model of type 2 diabetes (T2D), has previously been shown to lack pancreatic and duodenal homeobox gene 1 (Pdx-1) expression. Pdx-1 deficiency leads to pancreas agenesis in both mice and humans. We have therefore further examined the pancreas of P. obesus during embryonic development. Using Pdx-1 antisera raised against evolutionary conserved epitopes, we failed to detect Pdx-1 immunoreactivity at any time points. However, at E14.5, Nkx6.1 immunoreactivity marks the nuclei of all epithelial cells of the ventral and dorsal pancreatic buds and the only endocrine cell types found at this time point are glucagon and PYY. At E18.5 the pancreas is well branched and both glucagon- and ghrelin-positive cells are scattered or found in clusters, whereas insulin-positive cells are not found. At E22.5, the acini of the exocrine pancreas are starting to mature, and amylase and carboxypeptidase A immunoreactivity is found scattered and not in all acini. Ghrelin-, glucagon-, PYY-, gastrin-, somatostatin (SS)-, pancreatic polypeptide (PP)-, and insulin-immunoreactive cells are found scattered or in small groups within or lining the developing ductal epithelium as marked by cytokeratin 19. Using degenerate PCR, the P. obesus Neurogenin-3 (Ngn-3) gene was cloned. Nucleotide and amino acid sequences show high homology with known Ngn-3 sequences. Using specific antiserum, we can observe that Ngn-3-immunoreactive cells are rare at E14.5 but readily detectable at E18.5 and E22.5. In conclusion, despite the lack of detection of Pdx-1, the P. obesus pancreas develops similarly to Muridae species, and the Ngn-3 sequence and expression pattern is highly conserved in P. obesus.
T
Donev et al. (1989) found that the normoglycemic P. obesus pancreas expresses all the four major pancreatic hormones, and that two types of islets could be observed. These islets were called A islets or PP islets and could be distinguished by whether glucagon or pancreatic polypeptide (PP) was the prevalent cell type in the rims of the islets. PP islets were found to be most prevalent in the head of the pancreas, whereas A-type islets were mostly found in the tail portion of the pancreas (Donev et al. 1989). Interestingly, Pdx-1, a critical transcription factor for the formation of the pancreas and for the maintenance of β-cells, was shown to be deficient in the newborn P. obesus (Leibowitz et al. 2001). Several other well-characterized pancreatic factors, Nkx2.2 (NK2 transcription factor related, locus 2), Nkx6.1 (NK6 transcription factor related, locus 1), and ISL-1 (Lim-homeodomain protein Islet1) were all present in a normal distribution (Leibowitz et al. 2001). Our study is the first to investigate the hormone and transcription factor expression at multiple time points during development of P. obesus. In addition, we report on the cloning of the P. obesus Neurogenin-3 (Ngn-3) gene and localization of Ngn-3 immunoreactivity during development.
Materials and Methods
Animals
Animals were acquired from Hadassah Medical School (Jerusalem, Israel). All animals had free access to food (Koffolk 19560; Harlan Laboratories, Jerusalem, Israel) and water and were handled in accordance with the regulations specified under the Protection of Animals Act by the authorities in Denmark, European Union, and Novo Nordisk A/S. Several male and female P. obesus were housed together with one male and one female in each cage and with a barrier between the animals for the first 3 days. During the afternoon of the third day the barriers were removed, and on the fourth day the animals were separated. P. obesus has a gestation time of between 25 and 29 days in our housing environment, counting the day of separation as E0.5, and ∼25% of the females became pregnant. On the day of sacrifice (14.5, 18.5, and 22.5 days of gestation; n = 2, n = 4, n = 2, respectively), the animals were sedated with isoflurane, cervically dislocated, and the tissues removed. The time difference in embryonic days (E14.5 = E10.5, E18.5 = E12, and E22.5 = E17 in mice) in relation to the mouse differ due to the variable gestation period between the litters that we used in our studies.
Immunocytochemistry
The pancreas and parts of the intestines were removed from the embryos and fixed in 4% paraformaldehyde overnight. The tissue was then transferred to 70% ethanol, embedded in paraffin, and sectioned at 3-4 μm using the 2055 Autocut microtome from Leica Instruments GmbH (Wetzlar, Germany). In brief, the sections were deparaffinized and immunostained using the following protocol: 3% hydrogen peroxidase treatment for 5 min followed by microwave oven treatment at 650 W for 4 min and 160 W for 15 min in Tris-EGTA buffer, pH 9. The sections were then blocked in 10% normal donkey serum for 10 min and TNB blocking buffer (PerkinElmer; Hvidovre, Denmark) containing 0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl, 0.5% blocking reagent for 30 min and incubated with a mixture of two or three primary antibodies overnight (Table 1). Next, the sections were incubated with secondary biotinylated antibody (1:200) for 30 min and horseradish peroxidase-streptavidin conjugate for 15 min and developed with tyramide signal amplification substrate (Cy3, 1:100; PerkinElmer) for 10 min to visualize one of the antibodies. The one or two other primary antibodies were visualized with Cy2 (1:500) and Cy5 (1:500) using highly cross-absorbed secondary fluorescent antibodies for multiple labeling (Jackson ImmunoResearch Laboratories; West Chester, PA). Between each step the sections were rinsed in PBS. All pictures were taken using Zeiss LSM 510 META scanning laser microscope (Carl Zeiss; Oberkochen, Germany).
Antibody table
Gene Cloning
Embryonic pancreatic tissue was placed in RNA-later solution (Ambion; Austin, TX) overnight at 4C. RNA was prepared using the MicroRNA Isolation Kit (ZymoResearch; Hornby, Canada) for the E14.5 and the Qiagen Mini RNA Isolation kit (Qiagen Nordic; West Sussex, UK) for all other samples following the manufacturers' instructions. Cloning and sequencing of the Ngn-3 gene was done by Evrogen (Moscow, Russia). In brief, double-stranded cDNA was synthesized from total RNA extracted from P. obesus E18.5 tissue using the SMART (switching mechanism at the 5′ end of the RNA transcript) method (Clontech Laboratories; Mountain View, CA) (Zhu et al. 2001). Degenerate primers were designed on the basis of nucleotide and amino acid sequences from rat, mouse, and human and used for PCR amplification of the P. obesus Ngn-3 gene. The obtained fragment was cloned and sequenced, and new primers were designed for the subsequent 3′ and 5′ Step-Out rapid amplification of DNA ends (Matz et al. 1999). The resulting PCR products were cloned and sequenced. The Vector NTI (Invitrogen; Carlsbad, CA) software suite was used for sequence analysis and figure generation. The sequence of P. obesus Ngn-3 gene has been deposited in the GenBank database (accession number DQ790002).
In Situ Hybridization
Three-μm paraffin sections from E22.5 were deparaffinized and dehydrated in an ethanol series and rinsed in diethylpyrocarbonate-treated PBS. Sections were permeabilized with proteinase K (10 μg/ml) for 5 min followed by Triton X-100 for 30 min and rinsed in 5X sodium chloride sodium citrate (SSC) buffer. Slides were prehybridized at 60C for 1 hr followed by overnight incubation with antisense, sense, or a rat glucagon-positive control probe. Slides were rinsed with 5X, followed by 2X, followed by 1X SSC, and stored in TBS-Tween 20 (TBST). After 1 hr blocking in 10% normal sheep serum in TBST, the sheep anti-digoxigenin antiserum (1:2500) was added overnight at room temperature. Slides were rinsed in TBST followed by sodium chloride Tris-HCl magnesium chloride Triton X-100 (NTMT) and incubated with substrate nitrotetrazolium blue chloride/5-bromo-4-chloro-3-indolyl phosphate at 37C for 4 hr followed by overnight incubation at room temperature. The reaction was stopped in Tris-EGTA buffer, and the slides were rinsed in PBS and immunostained with guinea pig anti-insulin and rabbit anti-glucagon antibodies.
Results
Immunostainings of E14.5
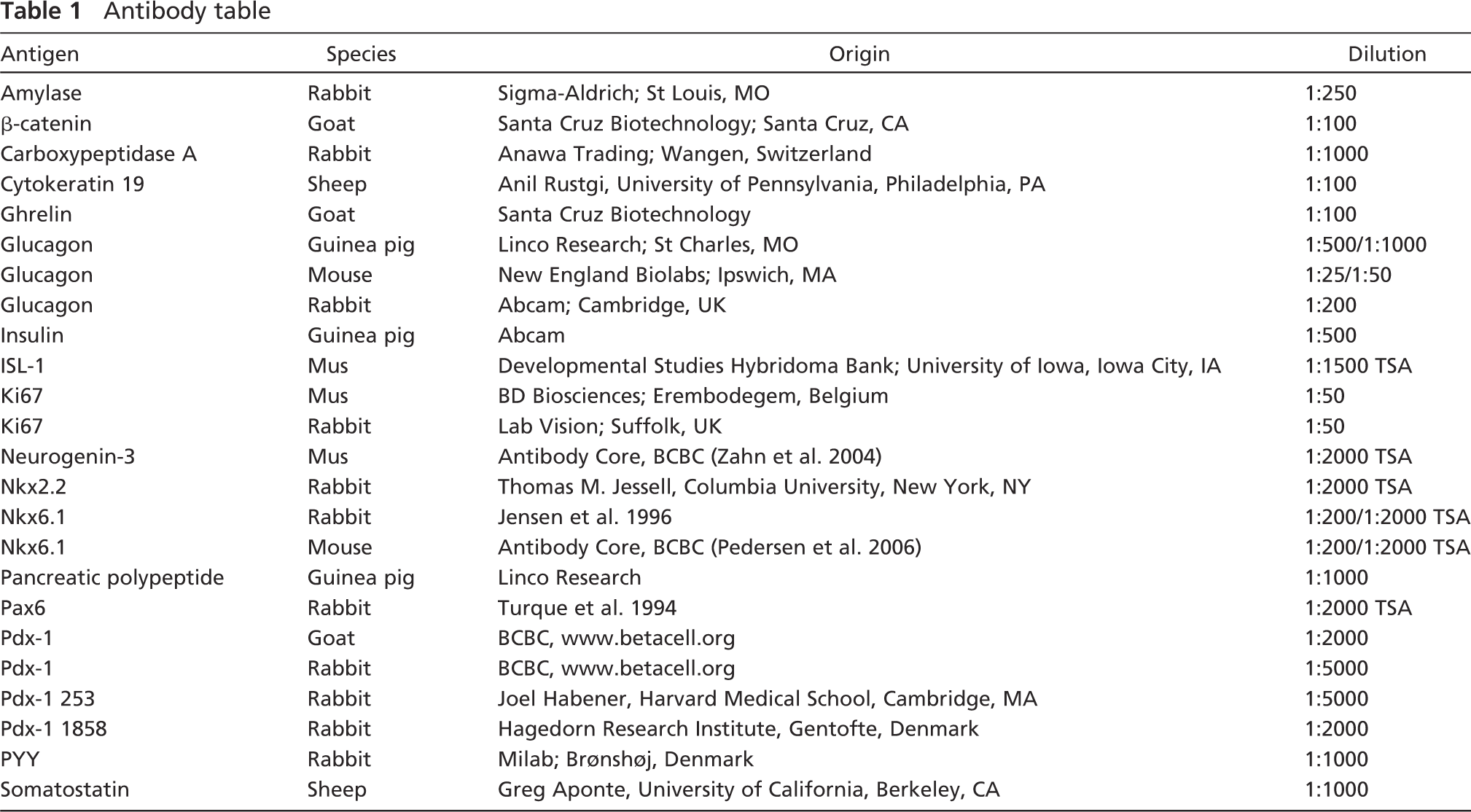
Embryonic tissue from day 14.5, which is approximately equivalent to mouse E10-10.5, was immunostained for the transcription factors Nkx6.1, ISL-1, and Ngn-3 and for β-catenin, glucagon, and PYY. The tissue did not stain with any Pdx-1 antibodies including those raised against N-terminally highly conserved epitopes (data not shown); hence, Nkx6.1 was used to identify the pancreatic epithelium. At E14.5, Nkx6.1 stains the entire pancreatic buds (Figures 1A, 1C, and 1E) including all the cells that will become endocrine, exocrine, and ductal. Nkx2.2 is broadly expressed in most of the dorsal bud and stains most strongly in the glucagon cells (Figure 1B). ISL-1 stains the mesenchymal nuclei surrounding the dorsal pancreatic bud (Figure 1C) as well as the nuclei of endocrine cells (only glucagon cells are detected at E14.5, data not shown), as expected. Most glucagon cells express PYY, but a few single hormone-expressing cells are also observed (Figure 1D). No other endocrine cells can be found at E14.5 (data not shown). β-catenin is involved in cell-cell adhesion and stains the pancreatic epithelium strongly and the surrounding mesenchyme more weakly (Figure 1E). Ngn-3 immunoreactive cells, which are the progenitors for all pancreatic endocrine cells, are quite rare at this time point and do not colocalize with Nkx6.1 (Figure 1E).
Immunostaining of E18.5
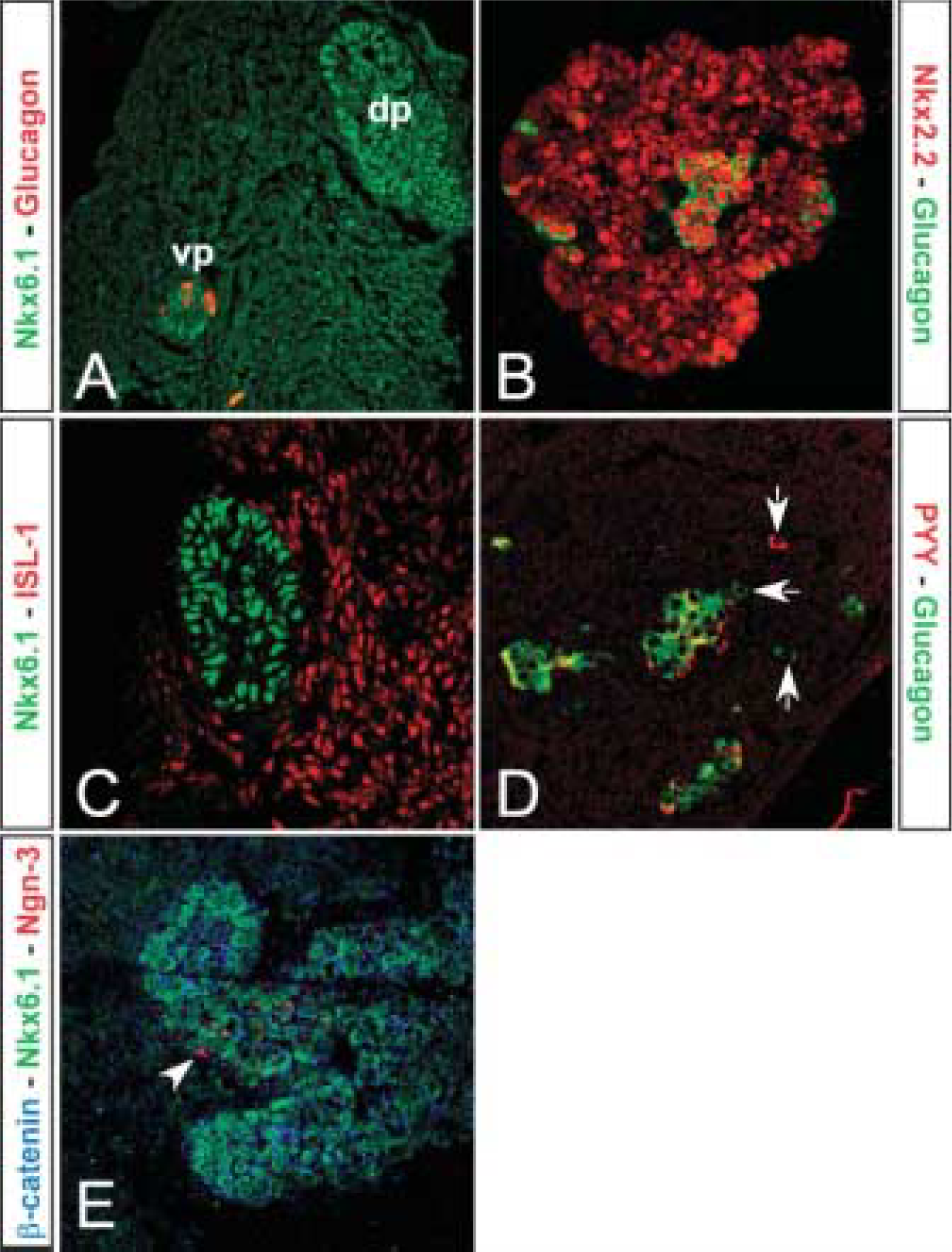
The pancreas is well branched at E18.5 (approximately equivalent to mouse E12.5), and the epithelium stains strongly for both cytokeratin 19 and β-catenin (Figures 2A and 2B). Pdx-1 immunoreactivity remained absent. Nkx6.1 cells are now scattered in the epithelium and do not colocalize with glucagon (Figure 2C). Ngn-3-positive cells were readily detectable at this time point (Figures 2A and 2B). Some Ngn-3-positive cells colocalize with Nkx6.1 and mark cells that have become restricted to an endocrine cell fate. The cells that are only Nkx6.1 positive are either less differentiated and earlier in development than the previously mentioned double-positive cells, or they are more differentiated cells that have lost Ngn-3 expression and are on their way to becoming β-cells (Figure 2B). Glucagon- (Figures 2C and 2D) and ghrelin-positive cells (data not shown) are scattered or found in clusters, whereas insulin-, somatostatin-, and PP-positive cells are not found (data not shown). Pax6 (paired box gene 6), an early endocrine cell marker, colocalizes with glucagon (Figure 2D).
Immunostainings of E22.5
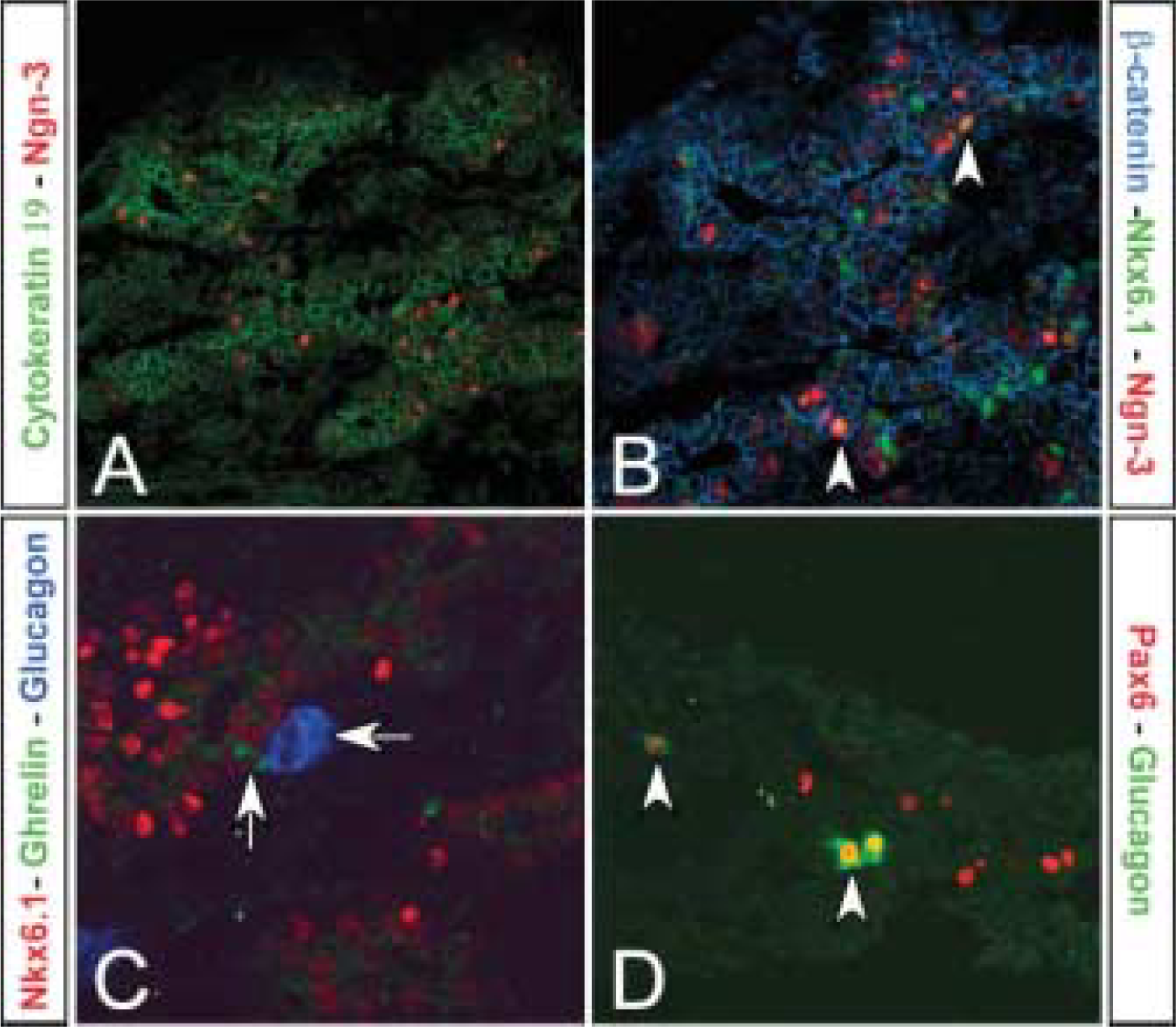
At E22.5, which is approximately equivalent to mouse E17, the acini of the exocrine pancreas are starting to mature, and carboxypeptidase A and amylase immunoreactivity are found scattered and not in all acini (Figures 3A and 3B). Insulin-, glucagon-, somatostatin-, PP-, and ghrelin-immunoreactive cells are now found scattered or in small groups within or lining the developing ductal epithelium (Figures 3C and 3D). PYY and gastrin are also observed at this time point (data not shown). Nkx2.2, Pax6, and ISL-1 are expressed in most endocrine cell types including insulin- and glucagon-positive cells (Figures 3E-3G). Nkx6.1 colocalizes now only with the insulin-immunoreactive cells (Figure 3H). Double-staining with Ki67 antibodies in insulin- or glucagon-positive cells shows that endocrine cells do not divide at this time point (Figure 3I).
Analysis of pancreatic markers at E14.5. (
Cloning and Expression of Ngn-3
The sequence of the P. obesus Ngn-3 gene was obtained using degenerate primers designed for conserved areas in the human, mouse, and rat Ngn-3 genes. By comparing the obtained sequence of Ngn-3 from the P. obesus with that of the human, mouse and rat, it is found that the P. obesus gene shows 78.7%, 85.7%, and 87.7% nucleotide homology to the previously mentioned species, respectively. The homology is also high examining the amino acid sequences, being 73.1% identical to human, 82.8% to mouse, and 85.6% to the rat proteins (Table 2).
Using in situ hybridization, we localized the endogenous Ngn-3 gene at E22.5 in scattered cells in the epithelium (Figures 4A and 4B). Double immunostaining on top of the in situ hybridization showed that the Ngn-3 gene did not colocalize with insulin and glucagon (Figure 4C) and showed the same pattern of scattered cells as was observed for the protein (Figure 4F). Using an antiserum directed against the mouse Ngn-3 protein, a more detailed analysis of Ngn-3 protein immunoreactivity was investigated. Ngn-3 cells are scattered in the epithelium at this time point and most do not express Nkx6.1 (Figure 4D), whose expression is becoming more restricted to the insulin-immunoreactive cells. Immunostaining for Ki67, an s-phase marker of dividing cells, and phosphohistone H3, also a marker of dividing cells, showed that the Ngn-3-immunoreactive cells and the endocrine cells are postmitotic at E22.5 (Figure 4E).
Analysis of pancreatic markers at E18.5. (
Discussion
Psammomys obesus belongs to a family of desert gerbils and is used as a model of human T2D. The absence of Pdx-1-like immunoreactivity remains a mystery and inspired us to further study how the pancreas develops in these animals compared with mice and rats. To our knowledge, this is the first comprehensive report on the development of the embryonic pancreas from any of the desert gerbil families (Meriones, Sekeetamys, Desmodillius, Pachyuromys, and Psammomys). We have demonstrated that during early development, E14.5 (equivalent to mouse E10-10.5), the P. obesus pancreas appears morphologically like wild-type mice and rats, and Nkx6.1 and β-catenin are good markers of the early pancreatic epithelium. At this time point, as in mice, we see very few hormone-positive cells, with only glucagon and PYY immunoreactive cells being present. This differs slightly from several mouse species where ghrelin-immunoreactive cells are observed at this time point (Prado et al. 2004; Heller et al. 2005). As previously reported in other rodent species, we also observed that most glucagon cells express PYY (Upchurch et al. 1994; Jackerott et al. 1996). In addition, in agreement with Leibowitz et al. (2001), we were unable to detect Pdx-1 immunoreactivity using several Pdx-1 antisera.
At E18.5 (approximately equivalent to E12.5 in mice), very few endocrine cells are present and most stain for glucagon. Nkx6.1 expression is now scattered in the epithelium as are other markers of endocrine cells, Nkx2.2 and Pax6. No exocrine pancreatic markers were observed. At embryonic day 22.5 (approximately equivalent to E17 in mice), we observed all the other endocrine cell types (insulin, somatostatin, PP, and ghrelin). In addition, we can observe cells immunoreactive for PYY and gastrin, hormones that are transiently expressed in the pancreas during development and are thought to be growth promoting (Gittes et al. 1993; Upchurch et al. 1994). This is the first time when amylase and carboxypeptidase A immunoreactivity is observed in the developing acini, but the pattern is quite heterogeneous, suggesting that the expression is just beginning and is slightly delayed as compared with mice and rats. This difference could also be reflected in the extremely variable gestation time of P. obesus.
Ngn-3 is the prerequisite gene for endocrine cell development (Gradwohl et al. 2000; Schwitzgebel et al. 2000). We have successfully cloned and characterized the expression of the P. obesus gene. The gene is highly homologous to all published Ngn-3 sequences. Using in situ hybridization, we could demonstrate that the Ngn-3 mRNA appeared in scattered cells in the epithelium and did not colocalize with either insulin or glucagon as has also been observed in mice (Jensen et al. 2000). Using immunohistochemistry, the Ngn-3 protein appears in a scattered pattern throughout the developing epithelium, in agreement with the in situ hybridization. At E14.5 Ngn-3-immunoreactive cells were very rare, but beginning at E18.5 we observed quite a few cells and these never colocalized with hormone-producing cells. Using two different markers of proliferation we could observe that the Ngn-3-positive cells were not proliferating, which is also what is observed in mice (Gradwohl et al. 2000; Jensen et al. 2000; Schwitzgebel et al. 2000).
Analysis of pancreatic markers at E22.5. (
Percentage similarity among different species at the protein level

In conclusion, the P. obesus pancreas shows a strikingly normal development of the endocrine, exocrine, and ductal tissue in the pancreas despite the apparent absence of Pdx-1. All the normal transcription factors we analyzed, except for Pdx-1, are expressed at a similar timing as has been reported in mice and rats. Because the lack of Pdx-1 in mouse (Jonsson et al. 1994; Offield et al. 1996) and human (Stoffers et al. 1997) leads to pancreas agenesis, we hypothesize that either Pdx-1 exists in P. obesus but is sufficiently mutated to escape immunological detection, or alternatively a related homeodomain protein may compensate in P. obesus for a functional deletion of Pdx-1 activity. Such compensation may be partial and restricted primarily to cover Pdx-1 function in pancreas development—and thus in part also explain the diabetic phenotype that may have been instrumental in selective survival in desert-like habitats.
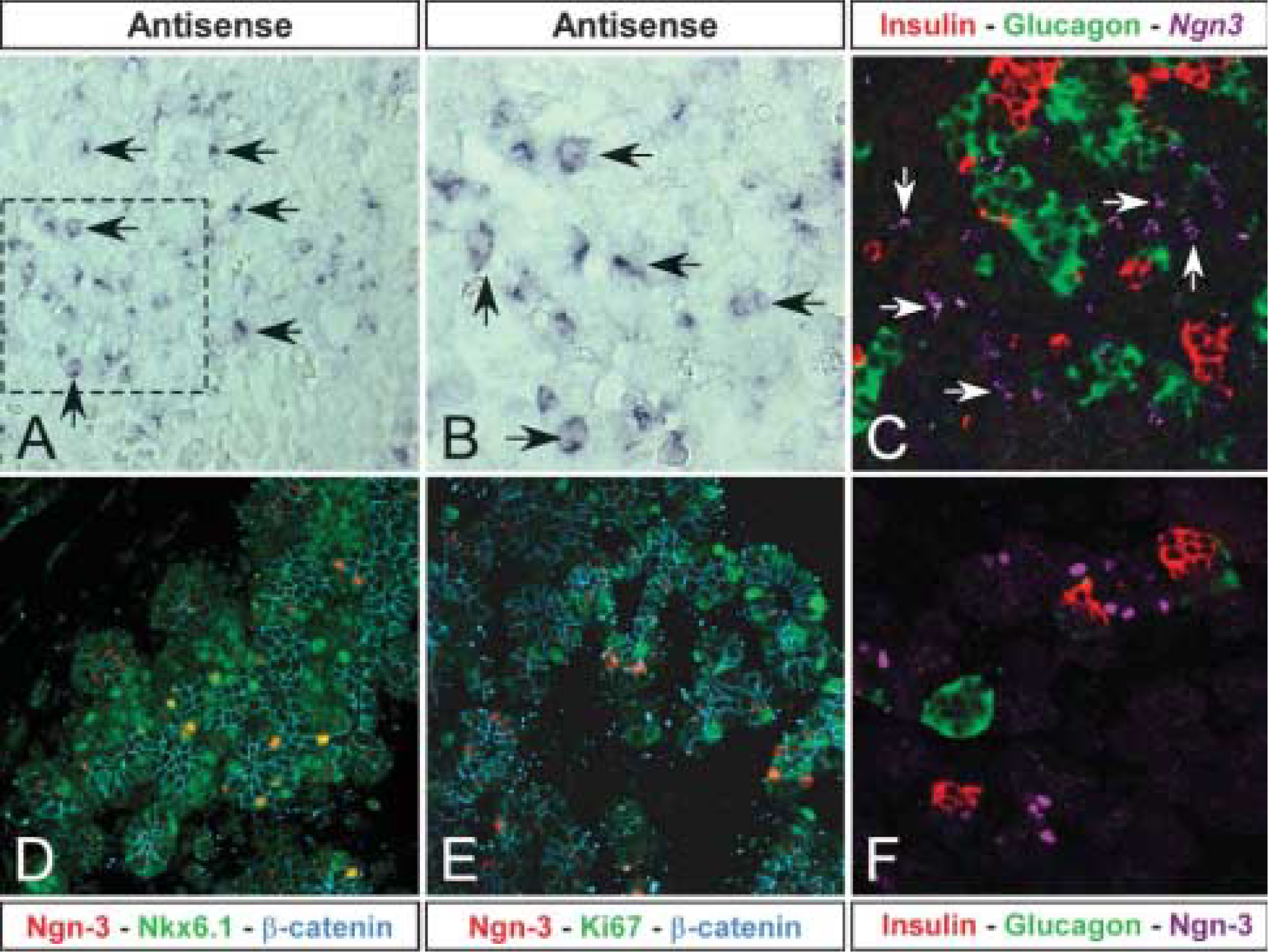
Expression of the Ngn-3 mRNA and protein at E22.5. (
Footnotes
Acknowledgements
This work is supported by an ErhvervsPhD award to Louise Vedtofte by Ministeriet for Videnskab, Teknologi og Udvikling, Denmark.
The authors thank Hanne Jensen-Holm, Thorhallur Tomas Buchholz, and Mikkel Schrøder Jørgensen for technical assistance with obtaining time-mated P. obesus, and Susanne Primdahl for assistance with tissue processing and embedding. The authors also thank Dr. Ole D. Madsen for critical feedback on this manuscript. The ISL-1 monoclonal antibody developed by Thomas M. Jessell was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biological Sciences, Iowa City, IA.