
Abstract
Androgen has been shown to promote the proliferation of prostate cancer through the action of the androgen receptor (AR). Mutation (T877A) of the AR gene found in an androgen-sensitive prostate cancer cell line, LNCaP, has been postulated to be involved in hypersensitivity and loss of specificity for androgen. In the present study, trafficking of AR and AR (T877A) in living prostate and non-prostate cancer cell lines under high and low concentrations of androgen and antiandrogen was investigated by tagging green fluorescent protein (GFP) to the receptors. In the presence of a high concentration of androgen, AR-GFP localized in the nucleus by forming discrete clusters in all cell lines. AR (T877A)-GFP was also translocated to the nucleus in LNCaP and COS-1 cells by the addition of a high concentration of androgen. In contrast, in the presence of a low concentration of androgen, the translocation of AR-GFP and AR (T877A)-GFP was observed in LNCaP cells, but not in COS-1 cells. Upon the addition of antiandrogen, AR-GFP was translocated to the nucleus but did not form subnuclear foci in both COS-1 and LNCaP cells, whereas AR (T877A)-GFP in both cells was translocated to the nucleus with subnuclear foci. The present study demonstrates the differential response of nuclear trafficking of AR and its mutant in prostate cancer cell lines and COS cells, and the subcellular and subnuclear compartmentalization provide important information on the sensitivity of the AR mutation.
Hormone architectonics of the tissue and cell have evolved from autoradiographic techniques. Tritiumlabeled steroids were systemically administered, and uptake cells were visualized by immersion of the emulsion (Eisenfeld 1975). Although it was useful for the demonstration of ligand distributions, it did not provide information on the receptor itself or precise sub-cellular distributions due to its limited resolution (Stumpf and Roth 1966; Stumpf 1983). Purification of steroid/nuclear receptors and the development of specific antibodies in conjunction with immunocytochemistry enabled us to investigate their distribution at the cellular level (Press and Greene 1988; Walsh et al. 1990). There is still controversy regarding the distribution of the receptors because sections were required for fixation and permeabilization. Recent studies have used green fluorescent protein (GFP), a 27-kDa protein from the jellyfish, Aequorea victoria (Shimomura et al. 1962), allowing us to directly detect its chimera protein without fixing and staining the cells. It also permits the real-time imaging of the subcellular localization of the chimera protein in live cells (Nordeen et al. 2001). This GFP-tagging method has clearly shown that steroid/nuclear receptors labeled with GFP retain their normal transcriptional activity and ligand-binding specificity (Hager et al. 2000; Kawata et al. 2001). Studies including ours on the subcellular localization of AR in living cells using GFP have been reported (Georget et al. 1997; Tyagi et al. 2000; Ochiai et al. 2004). After agonist binding, GFP-tagged ARs translocated from the cytoplasm to the nucleus and concentrated transiently in a subnuclear compartment that has the appearance of foci in a boundary region between euchromatin and heterochromatin (Tomura et al. 2001).
AR mutations with the substitution of amino acids in each domain have been implicated in the pathogenesis of a number of clinical disorders including prostate cancer (Heinlein and Chang 2004). The presence of AR mutations is generally found to increase with cancer stage and may contribute to the progression of prostate cancer and the failure of endocrine therapy by allowing AR transcriptional activation in response to antiandrogens or other endogenous hormones. In prostate cancer cells, replacement of threonine with alanine at the position of 877 of AR in the ligand-binding domain has been shown to be involved in hypersensitivity and the loss of specificity for androgen (Gaddipati et al. 1994; Suzuki et al. 1996). The molecular basis of AR function underlying these disorders remains unknown, and there are few reports on the trafficking of AR mutations in prostate cancer cell lines using GFP imaging analysis.
In the present study, real-time imaging of wild-type AR-GFP and mutated AR (T877A)-GFP was examined by comparing prostate cancer cells, which include LNCaP cells (androgen-sensitive prostate cancer cell line), DU 145 cells, PC-3 cells (hormone-refractory prostate cancer cell lines) and non-prostate cancer cells, and COS-1 cells at different concentrations of androgen and antiandrogen, with special emphasis on the trafficking in subcellular and subnuclear compartmentalization. Here we present the differential ligand-mediated response of GFP-labeled AR and its mutation in living cells.
Materials and Methods
Cell Culture
LNCaP, DU 145, and PC-3 cells were originally obtained from the American Type Culture Collection (Manassas, VA). LNCaP and PC-3 cells were maintained in RPMI 1640 (Nacalai Tesque; Kyoto, Japan) with 6% and 10% fetal bovine serum, respectively. DU 145 and COS-1 cells were maintained in DMEM (Invitrogen; Carlsbad, CA) with 10% fetal bovine serum.
Western Blot Analysis
For Western blot analysis, COS-1 cells were transiently transfected with pAR-GFP (see Plasmids and Transfection). The cells were solubilized in lysis buffer. Proteins were separated with SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride (PVDF) membrane (Immunobilon-P; Millipore, Billerica, MA) using a semi-dry blotting apparatus (Transblot-SD; Bio-Rad Laboratories, Hercules, CA). Immunoblot was performed with anti-AR antibody (PG-21; provided by Dr. G. Prins) diluted 1:1000 using chemiluminescent detection (ECL; Amersham Pharmacia Biotech, Piscataway, NJ). Characterization of the anti-AR antibody has been published elsewhere (Prins et al. 1991; Lu et al. 1998). Polyclonal rabbit antibodies were raised against amino acids 1–21 of the rat AR, and this PG-21 recognized both unliganded and liganded AR.
Immunocytochemistry and Immunofluorescence
Cultured cells were fixed for 15 min at 37C in 4% paraformaldehyde in PBS. Fixed cells were incubated with anti-AR antibody for 24 hr at 4C. Cultured cells were then reacted with biotinylated goat anti-rabbit IgG antibody for 1 hr at room temperature. Cultures were reacted with the streptavidin-biotin peroxidase complex [Histofine SAB-PO (R) kit; Nichirei, Tokyo, Japan] for 1 hr at room temperature. Cells were then visualized with 0.02% 3,3-diaminobenzidine (Sigma; St Louis, MO) and 0.006% H2O2 in Tris-HCl-buffered saline (pH 7.6). For immunofluorescence detection, Alexa Fluor 488-linked anti-rabbit IgG antibody (1:1000 dilution; Molecular Probes, Eugene, OR) was used as a secondary antibody.
Plasmids and Transfection
Vectors expressing AR-GFP and AR (T877A)-GFP were provided by Dr. H. Nawata (Taplin et al. 1995; Tomura et al. 2001). The N terminus of the GFP sequence was fused to the C terminus of the human AR sequence. Both nucleotide sequencing and Western blot confirmed the validity of the plasmid constructs. Cells were maintained overnight in a poly-
Confocal Laser-scanning Microscopy for Real-time Imaging of Living Cells
Cells transfected with the GFP chimera construct were plated on a poly-
Results
Characterization and Localization of Endogenous AR of Prostate Cancer Cell Lines
Because the presence of endogenous AR in prostate cancer cells has been controversial (Tilley et al. 1995; Alimirah et al. 2006), we investigated whether prostate cancer cell lines that include DU 145, PC-3, and LNCaP cells had endogenous AR by Western blot analysis using anti-AR antibody (Figure 1). AR protein was not expressed in DU 145 or in PC-3 cells but was expressed in LNCaP cells at the same level of expression as in COS-1 cells transfected with rAR.
Immunocytochemistry following the streptavidinbiotin peroxidase method with anti-AR antibody showed that AR immunoreactivity was observed only in LNCaP cells and not in DU 145 or PC-3 cells in prostate cancer cell lines. The result was consistent with Western blot analysis (Figure 2). Brown-colored reaction products showing endogenous AR were detected in both the cytoplasm and nucleus of LNCaP cells without ligand, and it was translocated completely to the nucleus in the presence of 10−6 M testosterone. As a control, we examined AR immunoreactivity in COS-1 cells transfected with plasmid vector expressing rAR. Immunoperoxidase reaction products showing AR were observed in the cytoplasm of COS-1 cells in the absence of testosterone, whereas AR immunoreactivity was seen in the nucleus after the addition of 10−6 M testosterone. We also performed a fluorescent method using Alexa 468-labeled IgG as a secondary antibody. The same result as with the immunoperoxidase method was observed: fluorescence was observed in both the cytoplasm and nucleus of LNCaP cells in the absence of testosterone, but bright immunofluorescence was seen only in the nucleus after the addition of 10−6 M testosterone. In addition, immunofluorescence of ligand-activated AR was observed as a discrete non-uniform pattern in comparison with a diffuse distribution in the absence of ligand.
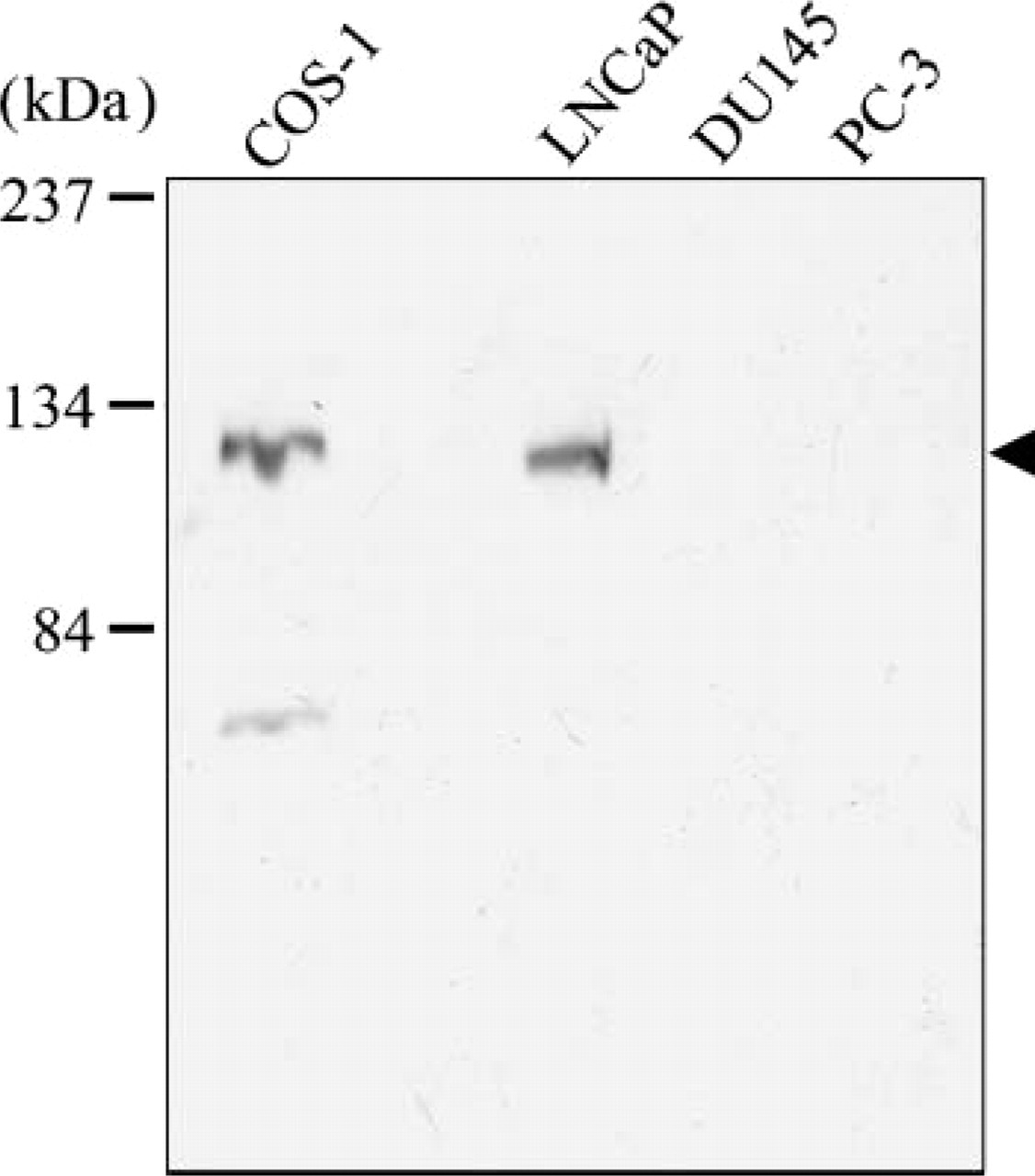
Expression of androgen receptor (AR) in prostate cancer cell lines. Cell lysates of prostate cancer cell lines, LNCaP, DU145, and PC-3, and COS-1 transfected with an expression plasmid of AR (COS-1) were applied to SDS-PAGE and blotted with anti-AR antibody. Specific bands were detected at the predicted molecular mass of 110 kDa in COS-1 and LNCaP lanes (arrowhead).
Subcellular Localization and Trafficking of AR-GFP With High Concentration of Androgen
We transfected plasmids expressing AR-GFP to COS-1, LNCaP, DU 145, and PC-3 cells, and fluorescent images were scanned of single living cells by confocal laser microscopy after the addition of 10−6 M testosterone (Figure 3). In cell lines of COS-1, DU 145, and PC-3, the fusion protein of AR-GFP was detected in the cytoplasm of these cells in the absence of androgens, whereas in LNCaP cells, AR-GFP was detected not only in the cytoplasm but also in the nucleus without nucleoli. The primary localization of AR-GFP in LNCaP cells was similar to the localization of endogenous AR. Upon the addition of testosterone, AR-GFP in all cell lines was time-dependently translocated into the nucleus and showed a discrete pattern that formed subnuclear foci. It took <20 min to form nuclear foci in all investigated cells after treatment with testosterone. Intranuclear foci were distributed evenly throughout the nucleus without nucleoli. Intensity of foci formation was not changed after overnight treatment with the ligand (data not shown). No significant difference in the localization and trafficking pattern of the AR-GFP was detected between LNCaP cells (androgen sensitive) and DU 145/PC-3 cells (androgen refractory).
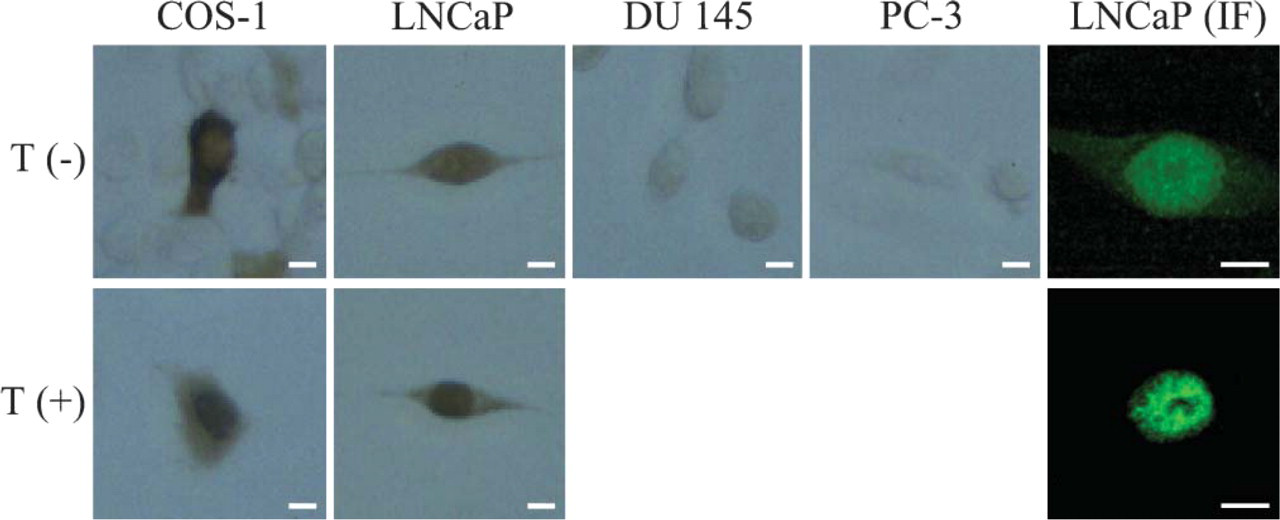
Localization of androgen receptors in prostate cancer cell lines. Immunocytochemical staining of prostate cancer cell lines, LNCaP, DU145, and PC-3, and COS-1 transfected with the expression plasmid of AR (COS-1) was performed with anti-AR antibody. Immunofluorescence staining for AR was performed with LNCaP cells (LNCaP IF). Upper: cells were cultured in the absence of ligand. Lower: cells were cultured in the presence of 10−6 M testosterone. Bar = 10 μm.
Subcellular Localization and Trafficking of AR (T877A)-GFP With High Concentration of Androgen
We transfected plasmids expressing AR (T877A)-GFP to COS-1 and LNCaP cells (Figure 4). In both cells, the primary localization of AR (T877A)-GFP was not only in the cytoplasm but also in the nucleus in the absence of testosterone. However, a predominance of AR-GFP was observed in the nucleus. Upon the addition of 10−6 M testosterone, AR (T877A)-GFP in both cell lines was time-dependently translocated into the nucleus and formed subnuclear foci. Trafficking patterns and distribution in the nucleus of both cells were the same as those observed in the case of wild-type AR-GFP.
Trafficking Pattern of AR-GFP and AR (T877A)-GFP With Low Concentration of Androgen
We also transfected plasmids expressing AR-GFP and AR (T877A)-GFP to COS-1 and LNCaP cells and observed the effect of a low concentration of androgen (Figure 5). With 10−9 M DHT treatment, the distribution of AR-GFP and AR (T877A)-GFP in COS-1 cells remained cytoplasmic, whereas those in LNCaP cells were translocated to the nucleus and formed subnuclear foci, showing the same pattern as in the high androgen concentration with regard to the time course, distributional pattern, and their size.
Trafficking Pattern of AR-GFP and AR (T877A)-GFP With Antiandrogen Treatment
We used bicalutamide (CAS), a competitive androgen antagonist, as a ligand at 10−5 M, almost the same as the plasma concentration of the clinical dose (Figure 6). Upon the addition of CAS, AR-GFP that was transfected in both COS-1 and LNCaP cells was translocated into the nucleus but did not form subnuclear foci, whereas AR (T877A)-GFP in both cells was translocated to the nucleus and formed typical subnuclear foci. The distribution and trafficking pattern of AR (T877A)-GFP in LNCaP cells and COS-1 cells were the same as those observed in the case of AR-GFP.
Discussion
The present study using a real-time imaging method clearly showed that wild-type AR-GFP and mutated AR (T877A)-GFP in prostate cancer cells, LNCaP, and non-prostate cancer cells, COS-1, translocated to their nucleus and formed subnuclear foci at a high concentration of androgen, whereas a low concentration of androgen led to AR-GFP and AR (T877A)-GFP forming nuclear foci only in the LNCaP cells and not in the COS-1 cells. In addition, it was demonstrated that anti-androgen treatment induced subnuclear foci in LNCaP and COS-1 cells that were transfected with AR (T877A)-GFP, but not in those with AR-GFP (Table 1).
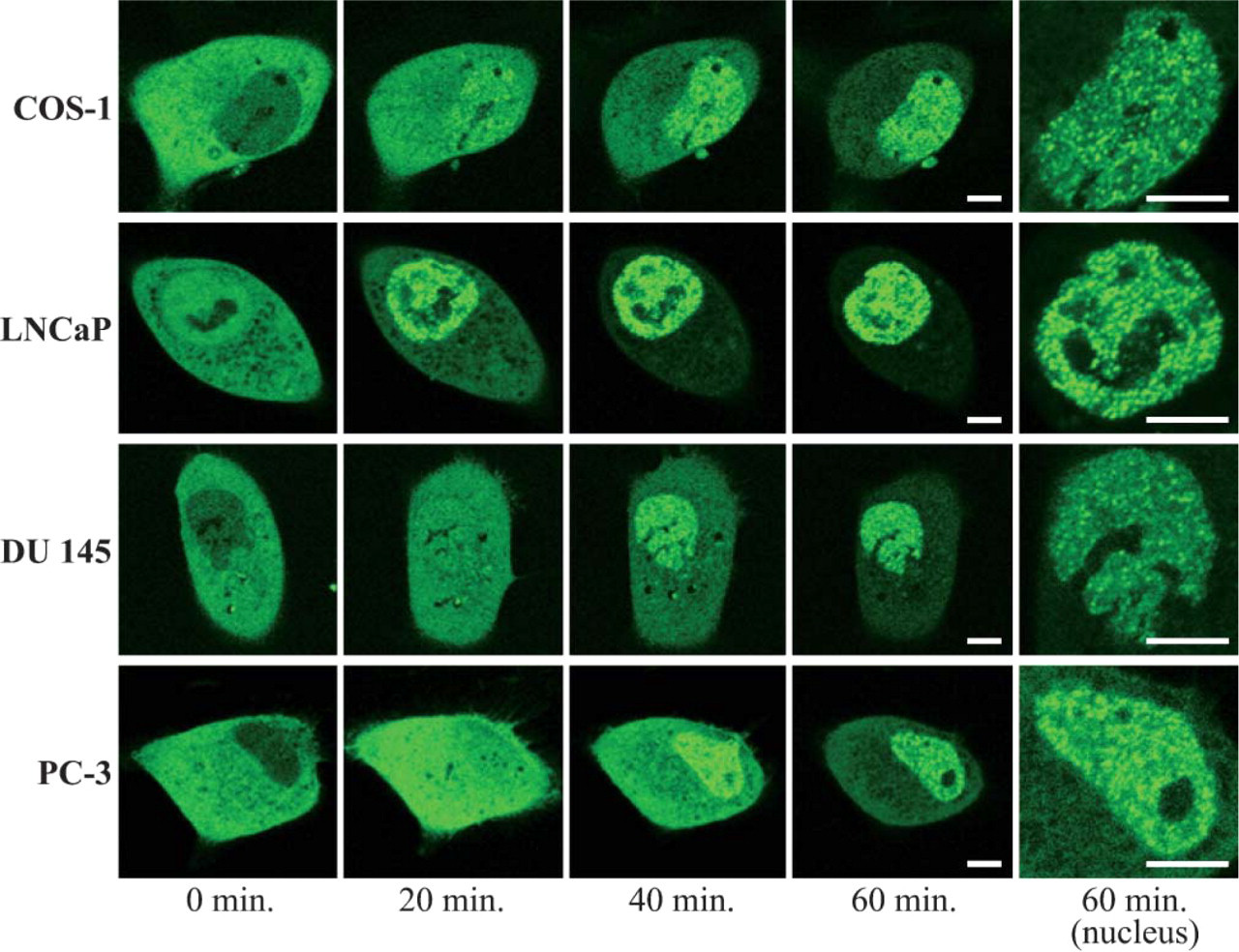
Real-time imaging of AR-green fluorescent protein (AR-GFP) in prostate cancer cell lines and COS-1 cells treated with 10−6 M testosterone. COS-1, LNCaP, DU 145, and PC-3 cells transfected with the expression plasmid of AR-GFP were cultured in the absence of ligand for 14 hr before observation. Fluorescent images were captured by confocal laser scanning microscopy every 20 min after the addition of 10−6 M testosterone. Bar = 5 μm.
AR activation by androgen is a multistep process. AR in the cytoplasm forms a complex with chaperone proteins such as heat-shock protein 90 (HSP90), HSP70, and HSP56 (Picard and Yamamoto 1987; Hager et al. 2000; Pratt et al. 2004). Ligand binding leads to a conformational change of its receptor followed by revealing the NLS (Simental et al. 1991; Zhou et al. 1994; Gelmann 2002). Many studies have suggested that exposed NLS of the cytoplasmic cargo protein is recognized by the importin family that mediates translocation to the nucleus through the nuclear complex, and the direct binding of Ran GTP to importin-cargo complex releases the cargo into the nucleoplasm (Picard and Yamamoto 1987; Poukka et al. 2000). There has been, however, no direct evidence of an interaction between AR and the importin family. Further studies are needed to elucidate the subcellular compartmentalization of AR.
With the use of the three-dimensional construction of confocal microscopic images with AR-GFP, it was demonstrated that after binding with the agonist, AR-GFP is concentrated in a subnuclear compartment that has the appearance of foci in a boundary region of the nucleus between euchromatin and heterochromatin (Tomura et al. 2001). The functional significance of these subnuclear foci has been discussed at the molecular level. The interaction of AR with this compartment is strongly biased for the agonist-bound form of AR (Tomura et al. 2001; Karvonen et al. 2002; Black et al. 2004). Several studies have shown that the interaction between AR and SRC1, GRIP-1, and the p160 coactivator family in these subnuclear foci is an agonist-dependent recruitment and/or release step that occurs before AR engages with chromatin (Karvonen et al. 2002; Black et al. 2004).
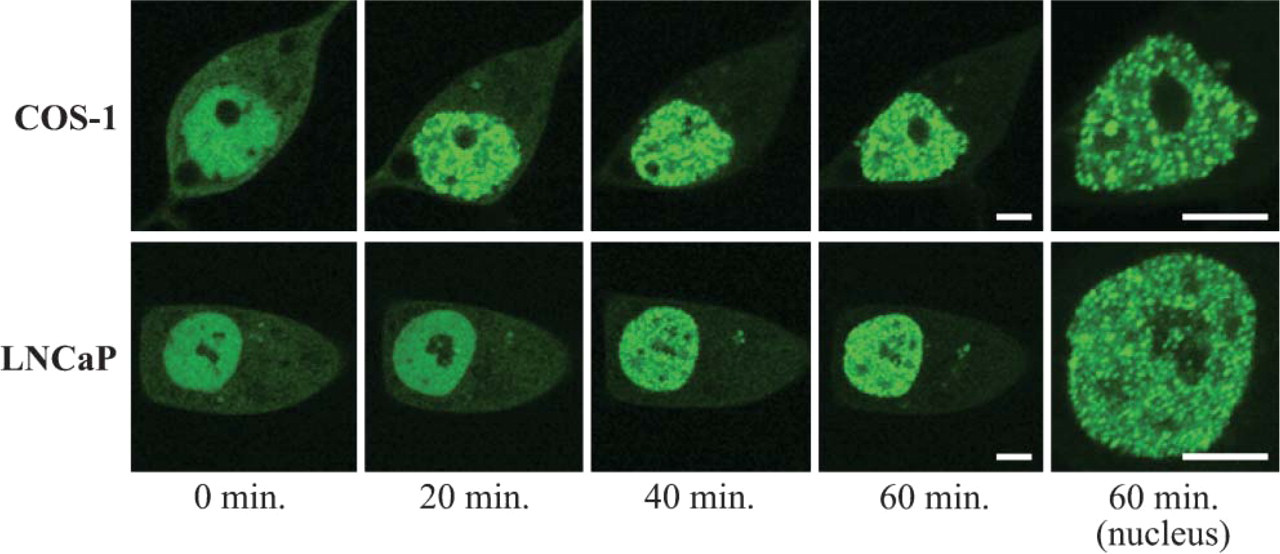
Real-time imaging of AR (T877A)-GFP in COS-1 and LNCaP cells treated with 10−6 M testosterone. COS-1 and LNCaP cells transfected with the expression plasmid of AR (T877A)-GFP were cultured in the absence of ligand for 14 hr before observation. Fluorescent images were captured by confocal laser scanning microscopy every 20 min after the addition of 10−6 M testosterone. Bar = 5 μm.
Most prostate cancer patients initially respond to androgen ablation therapy (Huggins and Hodges 1941); therefore, it is considered that androgen plays pivotal roles in the growth promotion of epithelial tissue of prostate cancer. The LH-RH analog excludes testicular androgen production, and antiandrogen excludes adrenal androgens. This combination of the two medicines is the first-line hormone therapy of prostate cancer. However, the disease eventually progresses to what is defined as hormone-refractory prostate cancer (HRPC), at which point the tumor is no longer responsive to androgen ablation, and uncontrolled progression of the disease is inevitable (Palmberg et al. 1999). Several possible mechanisms have been proposed (Grossmann et al. 2001). Mutations in the ligand-binding domain of the AR gene or amplification of the AR gene would increase AR sensitivity to a low concentration of androgen or allow it to respond to other steroids or anti-androgens. Many mutation sites of the AR gene in prostate cancer cells have been reported (Gottlieb et al. 2004), and codon 877 mutation (threonine to alanine) is the most frequently detected. This mutation is also found in LNCaP cell lines (Veldscholte et al. 1990). It was previously reported that mutations of the AR gene result in a higher affinity to androgen (Tilley et al. 1996). Moreover, it is an accepted theory that this results in the broadening ligand specificity of AR (Gaddipati et al. 1994; Taplin et al. 1995; Suzuki et al. 1996). Codon 877 of AR is located in the LBD and contacts the ligand directly. Mutation at this site alters the stereochemistry of helix 11 that forms part of the ligand-binding pocket. The present study showed that antiandrogen treatment did not induce the formation of subnuclear foci of AR-GFP in the COS-1 and LNCaP cells but apparently produced those foci of AR (T877A)-GFP in COS-1 and LNCaP cells. However, Farla et al. (2005) demonstrated that AR (T877A)-GFP did not form foci in CAS-treated Hep3B cells. These results indicate that T877A mutation broadens the specificity of ligand recognition (McDonald et al. 2000), but subnuclear foci formation of antiandrogen-bound AR (T877A) appears to depend on the intracellular environment, e.g., levels of interacting regulatory factors in the cellular milieu.
The present study demonstrated that AR-GFP and AR (T877A)-GFP in LNCaP cells were translocated to the nucleus and formed subnuclear foci at both low and high concentrations of androgen, and that those in COS-1 cells were translocated at the high concentration but not at the low concentration. These results suggest that T877A mutation of the AR gene does not account for AR hypersensitivity. It is supposed that hypersensitivity of AR might be due to the existence of some other intracellular factors such as chaperone proteins, signaling cascade, and cofactors in LNCap cells. Some cofactors, for example, the 160-kDa nuclear receptor coactivator (p160) family, interact directly with AF-1 of AR and can also bind to LBD (Ma et al. 1999). Consequently, they enhance the AR-mediated transcription of target genes in a ligand-dependent manner (Alen et al. 1999). One possibility is that an increase of the p160 family, such as glucocorticoid receptor-interacting protein 1 (GRIP-1) and the steroid receptor coactivator-1 (SRC1), may allow low concentrations of androgen to function more efficiently as an AR ligand. However, this is still controversial because expression levels of SRC1 protein in HRPC cells varied in experiments (Gregory et al. 2001; Linja et al. 2004).
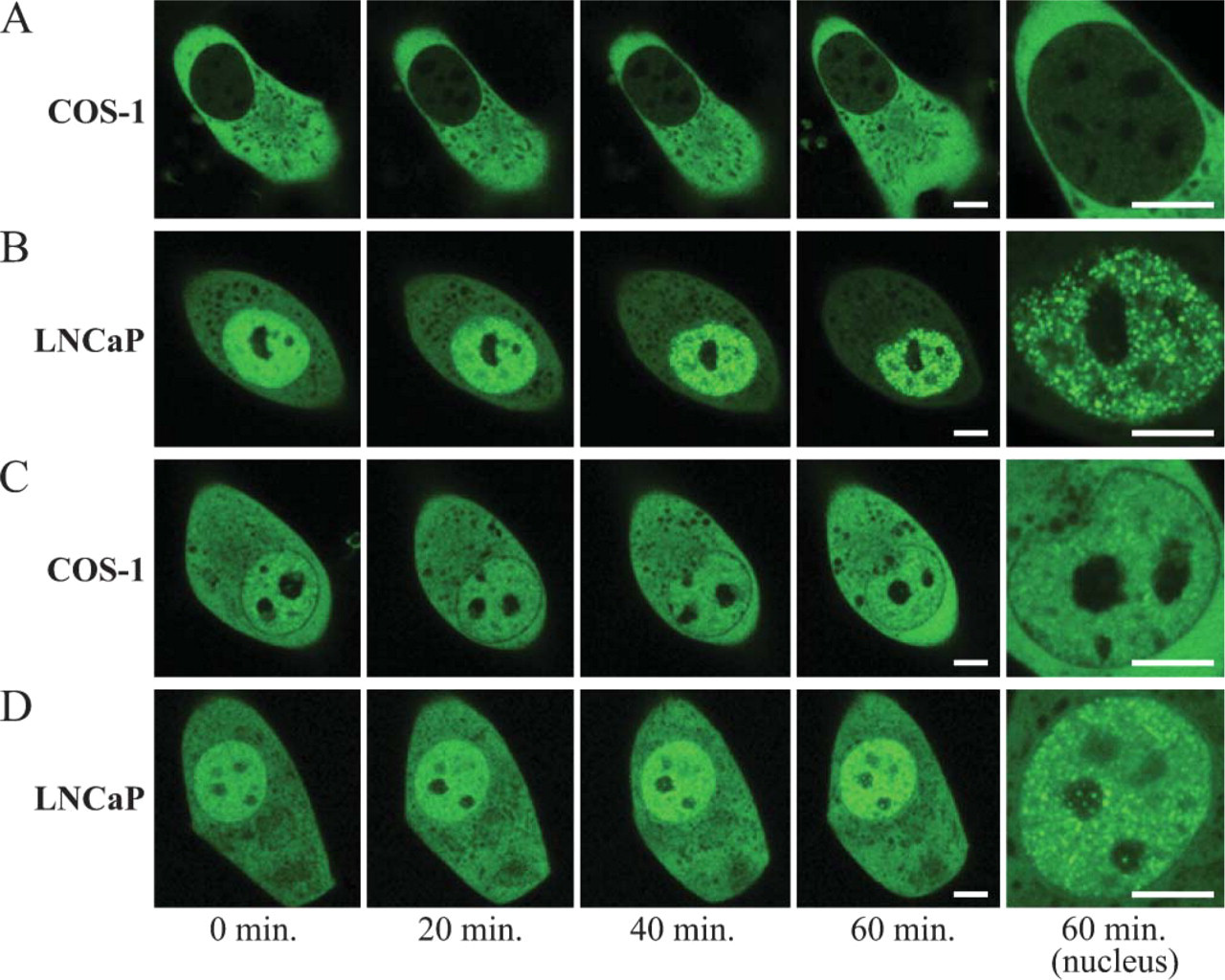
Real-time imaging of AR-GFP and AR (T877A)-GFP in COS-1 and LNCaP cells treated with 10−9 M dihydrotestosterone (DHT). COS-1 (
The fusion protein with AR was detected in the cytoplasm without androgen treatment in non-prostate cells; however, AR-GFP in LNCaP cells localizes not only in the cytoplasm but also in the nucleus. Subcellular factors in prostate cancer may activate the AR-signaling pathway itself or another one besides AR, having an interdependence on AR. Another possibility is that prostate cancer may have specific docking proteins for substrate binding in a nuclear import pathway. Furthermore, AR(T877A)-GFP in non-prostate cancer cells was detected in both the cytoplasm and nucleus. Alterations in the AR gene may lead to AR having an affinity for the nucleus.
The presence of endogenous AR in prostate cancer cells has been controversial (Tilley et al. 1995). The present study clearly showed that AR protein was not expressed in DU 145 or PC-3 cells, but was expressed in LNCaP cells at the same level of expression as in COS-1 cells transfected with rAR by Western blot analysis using anti-AR antibody. Immunocytochemistry by the streptavidin-biotin peroxidase method with anti-AR antibody was consistent with Western blot analysis, showing that AR immunoreactivity was observed only in LNCaP cells and not in DU 145 or PC-3 cells. A recent report demonstrated the expression of AR protein in DU 145 and PC-3 cells (Alimirah et al. 2006). This contradictory result might be due to the different antibodies used.
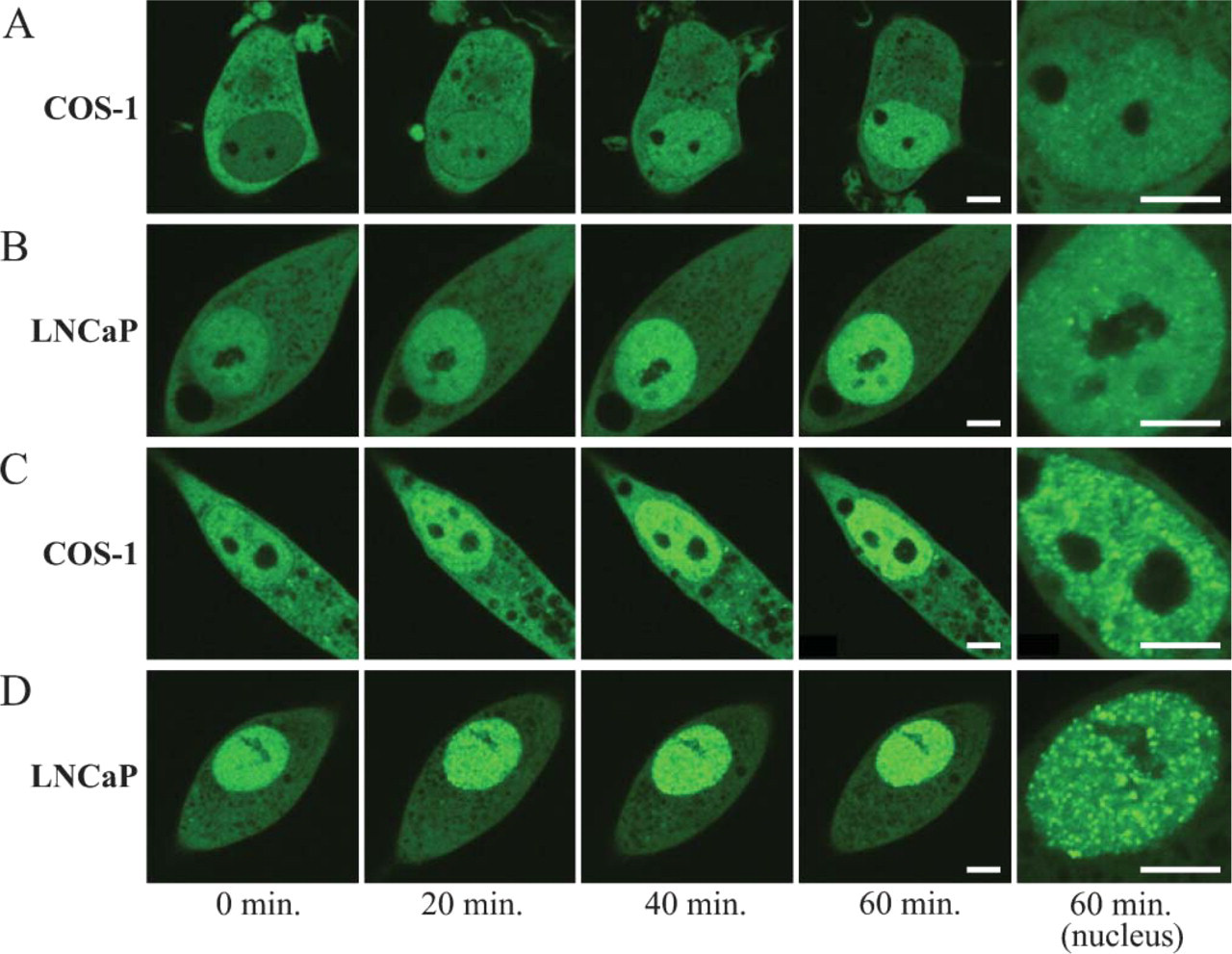
Real-time imaging of AR-GFP and AR (T877A)-GFP in COS-1 and LNCaP cells treated with 10−5 M bicalutamide (CAS). COS-1 (
The concepts of compartmentalization of AR in the cell and nuclear import of AR to the nucleus in response to androgen and antagonist have extended our understanding of the molecular mechanisms of prostate cancer progression. The present study of live imaging using GFP may facilitate further study on the relationship between AR and prostate cancer.
Summary of differential response of AR and AR (T877A)
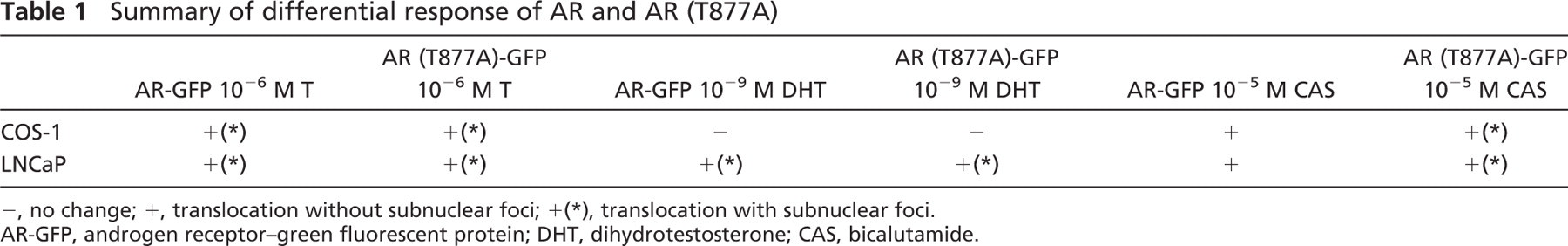
−, no change; +, translocation without subnuclear foci; +(∗), translocation with subnuclear foci.
AR-GFP, androgen receptor-green fluorescent protein; DHT, dihydrotestosterone; CAS, bicalutamide.
Footnotes
Acknowledgements
This work was supported by Grants-in-Aid for Scientific Research from the Ministry of Education, Science, Sports, Culture and Technology, Japan, grant numbers 16700306, 16200026, and 14571514 (to KM, TM, MK).
We thank Dr. Hajime Nawata for the kind gift of AR-GFP and AR (T877A)-GFP.