
Abstract
There is a lack of agreement on the distribution of islet amyloid polypeptide (IAPP) in the pancreases of healthy and diabetic subjects. Therefore, a detailed morphometrical and immunohistochemical study was performed to obtain information on the distribution of cells expressing insulin, glucagon, somatostatin, pancreatic polypeptide (PP), and IAPP in the pancreases of non-diabetic (n=4) and diabetic individuals (n=6). In the non-diabetic cases, β-cells contributed to ∼64%, α-cells to 26%, δ-cells to 8%, PP cells to 0.3%, and IAPP cells to 34% of the islet cell population. The ratio of IAPP/insulin was ∼1:2. In diabetic cases, β-cells were decreased by 24%, and IAPP was decreased by 57%. The α- and δ-cells were increased by 40% and 58%, respectively. IAPP/insulin ratio was decreased by 41%. Thus, only 50% of the β-cells in non-diabetics and only 30% in diabetics coexpressed IAPP. In diabetics, more δ-cells coexpressed IAPP than in non-diabetics. The results seem to argue against the notion that the secretion of IAPP is increased in diabetics. It is possible that an increase in somatostatin and glucagon plays a greater role in diabetes than IAPP.
T
Materials and Methods
Tissues
Eleven pancreases from donors, all due to accidental deaths, were examined (Table 1). Six of the donors had type 2 diabetes (DM); only one required insulin (case 6). The remaining cases (two males and three females) had no history of pancreatic disease. Average age of donors was 44.8 years (range 18–70 years). Two of the type 2 diabetic patients were female and four were male, with an average age of 55 years (range 27–84 years). We do not know which region of the pancreas the samples came from. Based on immunohistochemical (IHC) results, however, all tissues were from the glucagon-rich region. All tissues were fixed in formalin and processed for histology by conventional methods. One section of each sample was stained with hematoxylin and eosin (H&E) and evaluated histopathologically for suitability for the study using the following criteria: (1) intact tissue with no evidence of autolysis, (2) sample diameter of at least 1.0 × 1.0 cm, and (3) an adequate number of islets (at least five islets per section). Selected tissues were cut in 30 serial sections.
IHC
Immunostaining was carried out using an avidin-biotinperoxidase complex (ABC) method with mouse monoclonal anti-IAPP antibody (clone R10/99; Lab Vision, Fremont, CA); rabbit polyclonal IAPP antibody (Abcam; Cambridge, MA); mouse monoclonal anti-insulin antibody; and polyclonal antibodies bodies against glucagon, somatostatin, and pancreatic polypeptide (PP) (Zymed Laboratories; South San Francisco, CA). For IAPP, IHC staining antigen retrieval was performed by incubating the sections with 0.1 N citrate buffer (pH 6.2) at 100C for 10 min. For polyclonal antibody, protinase K was also used. Several methods were used for visualization of islet hormones. Consecutive serial sections were processed with individual antibodies, and microphotographs were taken from each section. Sections stained with IAPP and another antibody (insulin, glucagon, somatostatin, or PP) were superimposed in Adobe Photoshop (Adobe Systems; Mountain View, CA), and the stained cells in each layer of Photoshop were counted. Only nucleated stained cells were counted. This technique allowed us to identify cells that coexpressed both hormones (Figure 1). In addition, slides were also processed by a multi-labeling method (Pour et al. 1994) to demonstrate the ratio of immunoreactive cells in each islet and for the detection of cells costained with IAPP and other endocrine cells. One of the antibodies was processed either with Histomark red (Kirkegaard and Perry Laboratories; Gaithersburg, MD) to obtain a red color, with Histomark blue giving a blue color, or with DAB producing a brown color (Kirkegaard and Perry Laboratories). The second antibody was processed with a different color than the first antibody (Figure 1). In a series of other serial sections, the reverse color combination was used. For example, in the first set, β-cells were stained red and IAPP were stained blue. In the second set, β-cells were stained blue and IAPP were stained red. Coexpression of the hormones was judged by the change of color hue [for example, brownish red (red and brown) or purple (red and blue)]. In the multi-labeling method, insulin was represented in red and IAPP in blue in one slide. In the next slide, insulin was represented in blue and IAPP in red. This method allowed us to more accurately differentiate the cell type as well as determine numbers and the ratio within each islet. For proper evaluation of color intensity, no counterstaining was performed. Negative control slides were processed similarly except that either no primary antibody was used (for monoclonal antibody) or a non-immunized serum was used instead of the primary antibody (for polyclonal antibody). In tissues from diabetics only, islets unaffected or slightly affected by amyloid deposits were evaluated.
Relative distribution of pancreatic endocrine cells within the islets of normal and diabetic cases a
Values expressed as mean ± SD.
Significant difference (p<0.001) compared to NP.
Significant difference (p<0.01) compared to NP.
NP, normal pancreas; DM, pancreas from diabetic cases.
Determination of Islet Size
In the H&E-stained slides, diameters of ∼200 randomly selected islets/pancreas were measured by a microscale using a Zeiss Axiomat microscope (Carl Zeiss; Oberkochen, Germany). The average value was considered to be the size of the islets (area: μm2 = π × length a/2 × length b/2).
Statistical Evaluation
Results are presented as mean ± SD. Differences in significance between groups was tested using ANOVA with Bonferroni correction for pairwise comparisons.
Results
Normal Pancreas
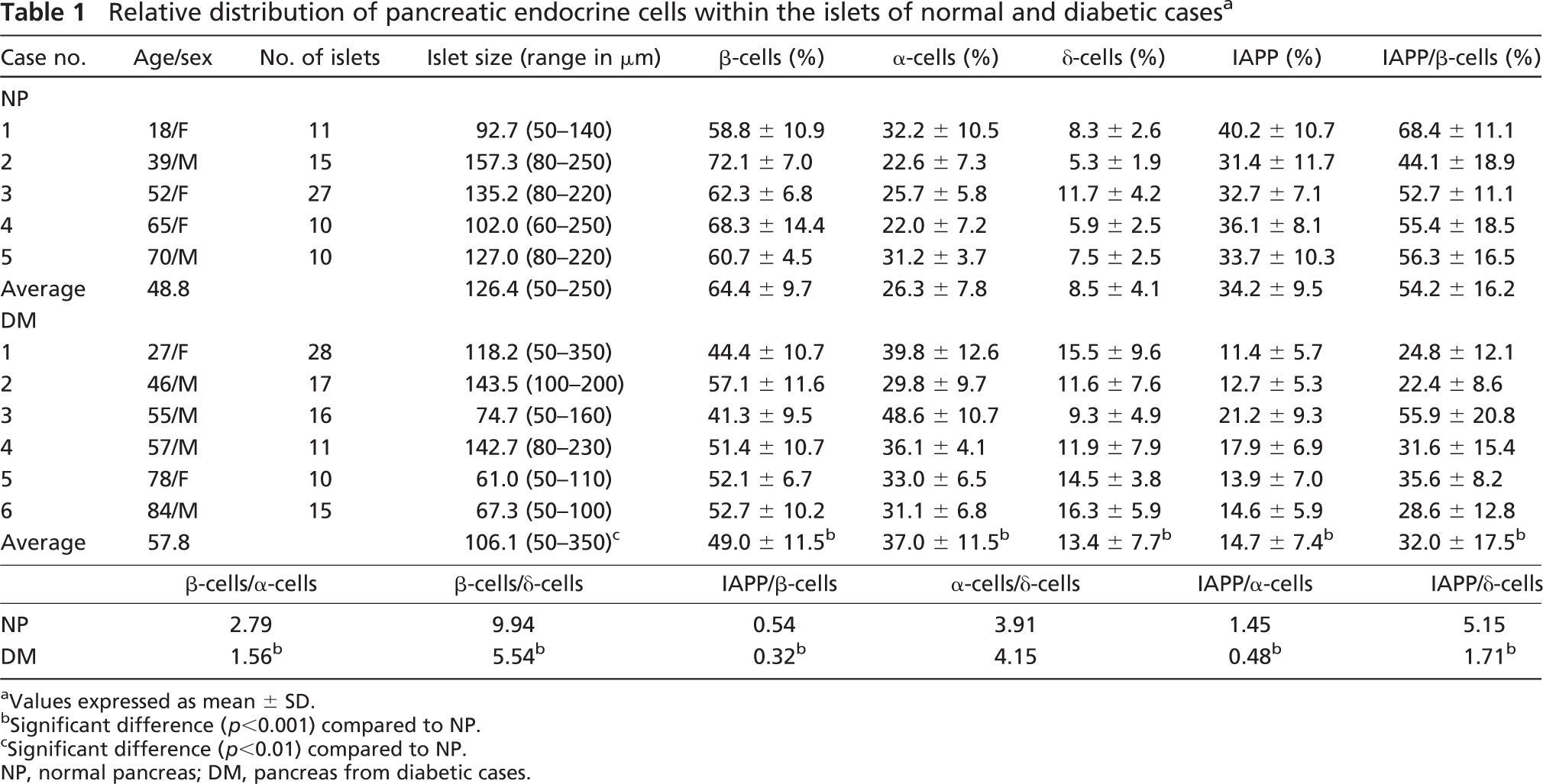
The number of islets per specimen and the size and percentage of the endocrine cells are summarized in Table 1. Islet size varied between 50 and 250 μm and was smaller in the younger case (case 1) than in the others (Table 1). The number of individual islet cells varied among the cases and even within the same tissue. The number of islet cells did not differ significantly among cases and was independent of the size of the islets. In each case and in every islet, the number of IAPP cells was about one-half the number of β-cells, except in case 1, which had the highest number of IAPP cells. The difference, however, was not significant. Although expression of IAPP coincided with the β-cells in each case, a few cells in the islet periphery were expressed in cells unstained with anti-insulin (Figure 1).
Demonstration of IAPP-expressing cells in the islets of non-diabetic and type 2 diabetic cases by immunohistochemistry (IHC). Avidinbiotin-peroxidase complex (ABC) method.
Results of the monoclonal and polyclonal antibodies did not differ except for the quality of the staining, which was stronger with the monoclonal antibody. This was also the case in the pancreases from the diabetic cases.
Pancreases in Type 2 Diabetic Cases
Most islets in these cases were fragmented and presented aggregates of β-cells with ill-defined borders.
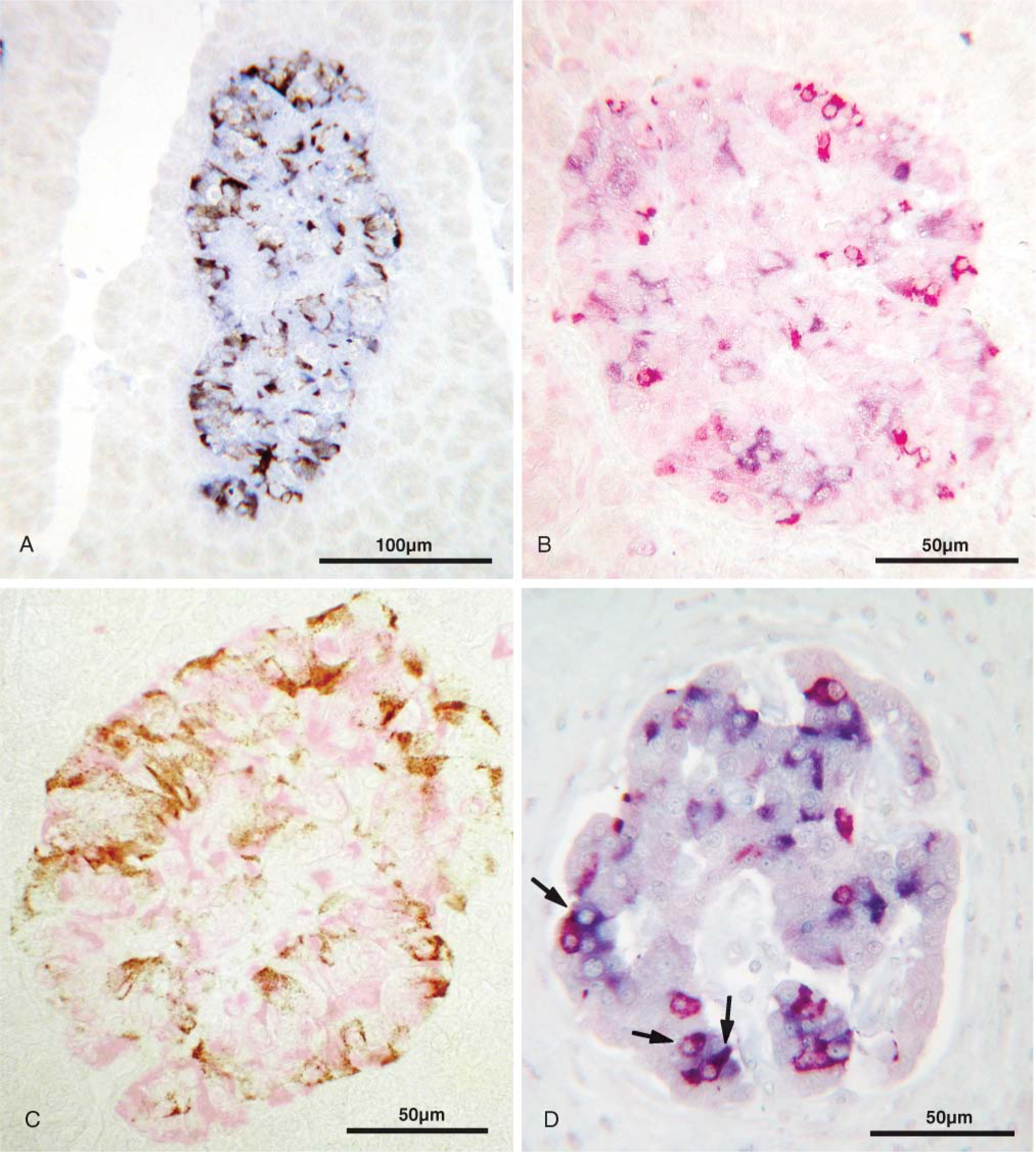
Some islets appeared unusually large because of amyloid deposition and edema, measuring between 200 and 350 μm. Amyloid deposits were strongly stained with Congo red but showed no reactivity or only a weak reactivity to anti-IAPP. Excluding these large islets, the size of intact islets was significantly smaller in all six cases. The numbers of IAPP and β-cells were lower and the numbers of δ and α cells were higher than in the controls (normal pancreases). Compared with the normal pancreas, percentage of insulin cells within the islets of all six cases was significantly lower as was the number of IAPP cells, which were predominantly coexpressed in β-cells. The IAPP/insulin ratio was also significantly lower than in the control cases (Table 1).
Remarkably, there was a marked proliferation of centroacinar cells in many areas close to or remote from the distorted islets in all six cases (Figure 2D). These cells, generally located within the acini, had a large semi-transparent cytoplasm (Figure 2) and reacted with both IAPP antibodies (Figure 3). Between these cells there were a few or single cells immunoreactive with insulin, glucagon, somatostatin, or PP antibodies (Figure 3B).
No differences were found in the number of PP cells among the cases and between the pancreases of normal cases and diabetic cases.
Discussion
IAPP has been shown to inhibit the secretion of a variety of hormones including insulin (Stridsberg and Wilander 1991; Stridsberg et al. 1993). Its hypersecretion has been implicated in the development of diabetes (Johnson et al. 1989; Tokuyama et al. 1997). There are, however, no convincing data to support this view, and the available reports are contradictory. Early studies indicated that IAPP is coproduced and cosecreted with insulin (Hanabusa et al. 1992). Recent investigations have shown that this is true for the normal pancreas; however, in the disease states, especially in diabetes, insulin and IAPP can be secreted independently (Mulder et al. 1995a,b,1996; Ahren and Gutniak 1997). The manifestation of type 2 diabetes has been claimed to influence the capacity of β-cells to synthesize and secrete IAPP (Takada et al. 1996). IAPP causes β-cell death and impaired insulin secretion (Janson et al. 1996). It was also found that the expression of IAPP is not restricted to β-cells. It is also coexpressed in non-β-cells (Madsen et al. 1991) and predominantly in somatostatin cells (De Vroede et al. 1992). Differences in IAPP expression have also been reported in obesity, impaired glucose tolerance, and non-insulin-dependent diabetes (Enoki et al. 1992). The opinions about the role of amyloid deposition within islets as the source of β-cell toxicity vary considerably (Janson et al. 1996; Marzban et al. 2003). IAPP-deficient mice develop a more severe form of alloxan-induced diabetes, indicating a beneficial feature of IAPP in β-cell damage (Mulder et al. 2000b). On the other hand, administration of exogenous insulin has been shown to lower the secretion of IAPP, suggesting that insulin treatment may delay β-cell damage (Lindstrom et al. 1997).
In the present study, some undisclosed patterns of pancreatic endocrine cells in non-diabetics and type 2 diabetics were noted. Although it is generally believed that IAPP is coproduced, costored, and cosecreted with insulin, in our series only ∼50% of pancreatic β-cells in non-diabetic cases coexpressed IAPP. Although the number of cases was limited, the remarkable similarities in the findings among the cases point to the general pattern of IAPP distribution in the normal and diabetic cases. Because the intensity of the staining with the IAPP antibody was consistently strong (indicating its high concentration), it is unlikely that IAPP was completely discharged from the cells without leaving traces behind. Although variations in the number of endocrine cells among the non-diabetic cases could be due to differences in the digestive state, the fixed IAPP/insulin ratio in all non-diabetic cases strongly suggests that this ratio is physiological and that only 50% of β-cells produce IAPP. Whether IAPP is consistently expressed in a certain β-cell population or randomly in only half of the cells requires further examination. In non-diabetics, the ratio of IAPP/insulin is reportedly ∼1:7 (Hanabusa et al. 1992). In our study this ratio was ∼1:2. Therefore, morphometrical findings do not seem to correlate with the clinical laboratory data.
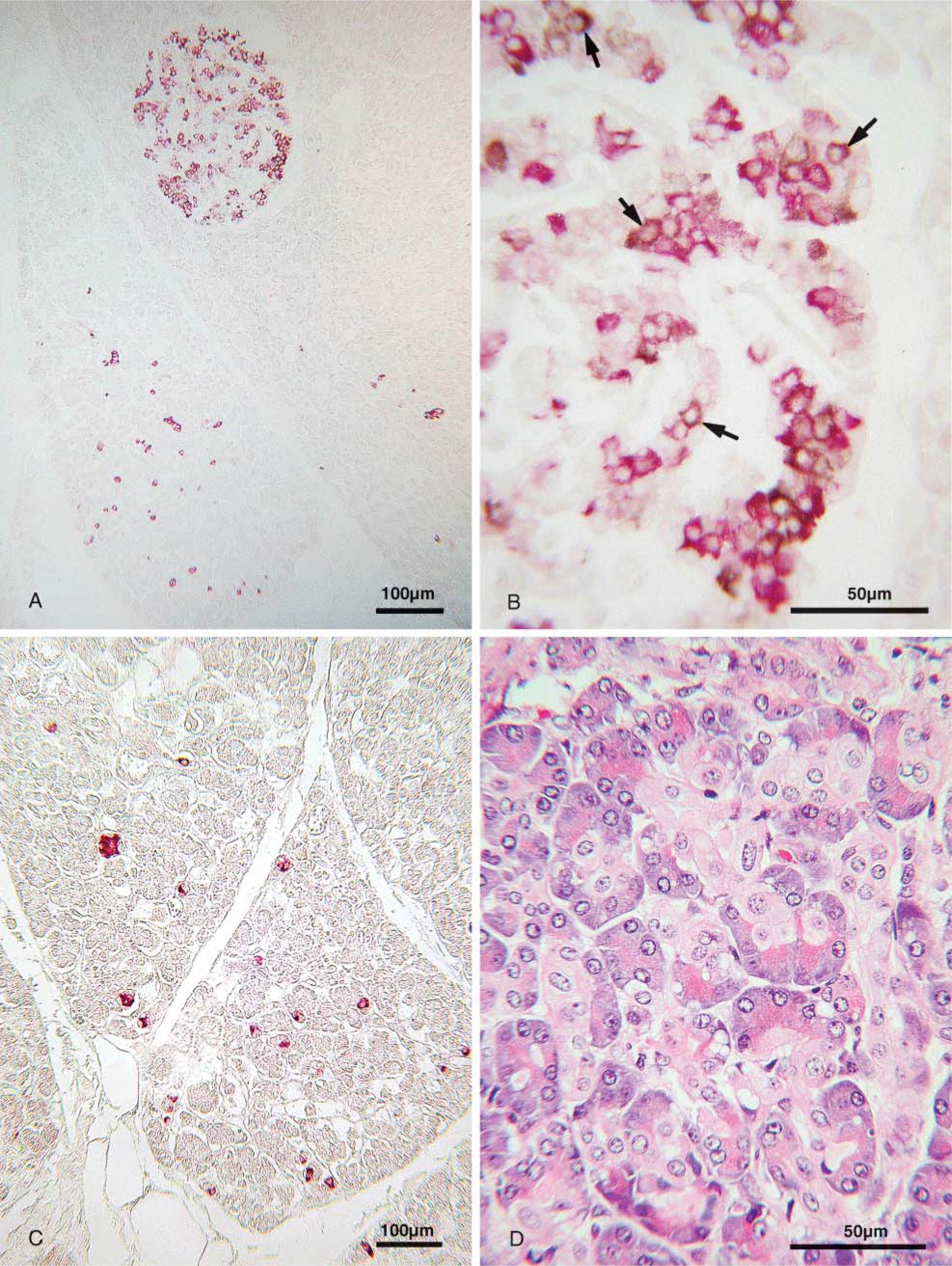
Examination of the pancreases of diabetic cases by IHC
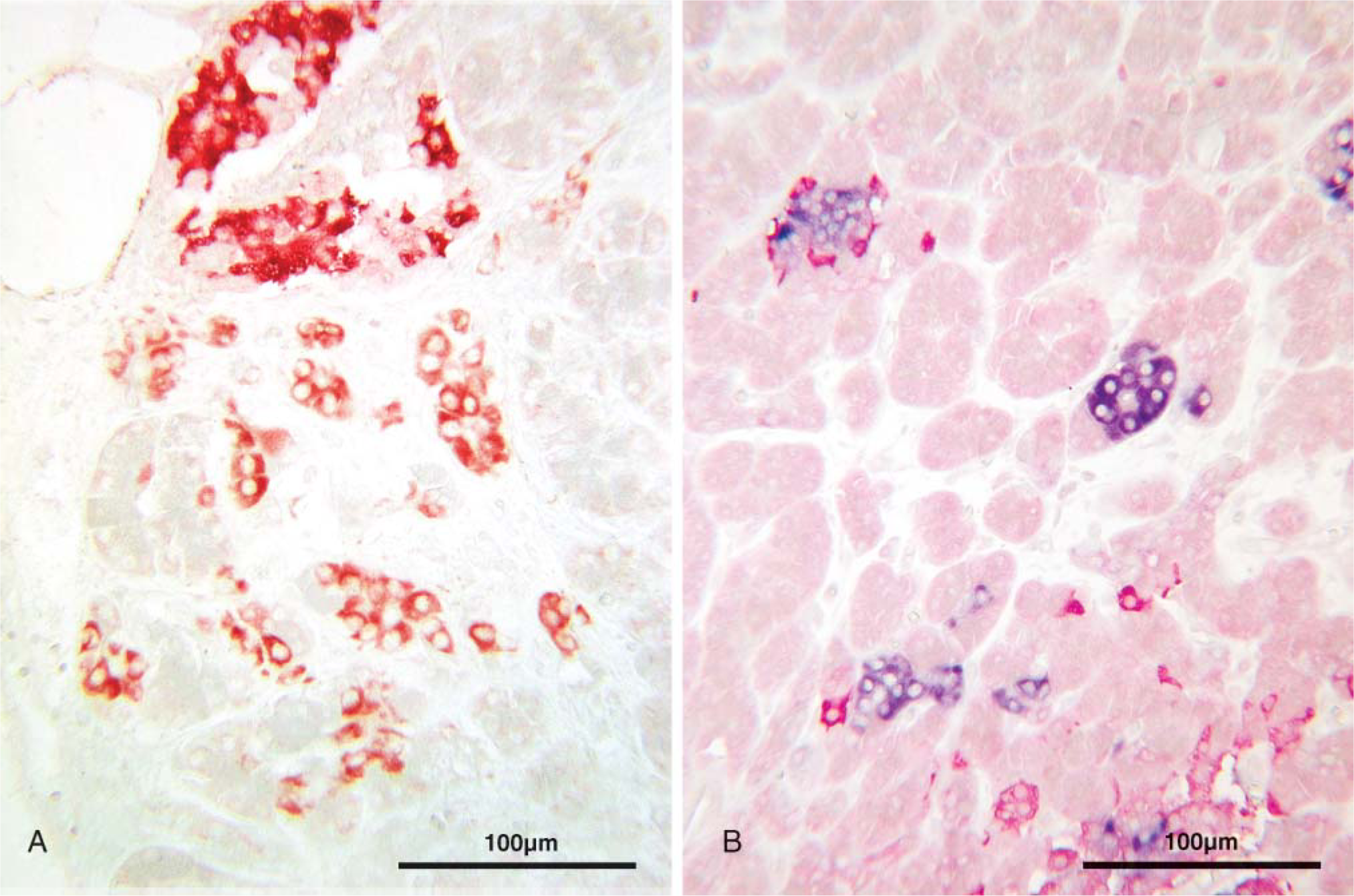
Pancreases of diabetic cases processed by IHC (ABC method).
If diabetes is caused by an increase of IAPP secretion from β-cells as reported, a larger number of IAPP-expressing cells was expected to be found in diabetic cases. The opposite, however, was noted. The number of cells immunoreactive with anti-insulin and anti-IAPP was significantly lower in diabetic than in non-diabetic cases. Also, a lower number of β-cells coexpressed IAPP. Reduction of β-cells by ∼24% is in agreement with a previous study (Clark et al. 1995). Reduction in the number of IAPP-expressing cells in the diabetic cases, however, was 57%, and the ratio of IAPP/insulin was 41% less than in non-diabetic cases. Possibly the amount of IAPP released by each β-cell is more important than the number of β-cells that produce IAPP in diabetics. Unfortunately, we did not have the circulating level of IAPP in our cases to reach a conclusion.
We did not have any information on the therapeutic scheme of the type 2 diabetic cases. Based on the severity of the islet alteration in case 11, we assumed that the patient was treated with insulin. If so, then there was no difference in the pattern of IAPP between this and the other cases. Contrary to the experimental study (Lindstrom et al. 1997), this indicates that exogenous insulin in diabetics has no effect on the pattern of IAPP. If the remaining cases were non-insulin dependent, then our data are in agreement with the clinical study that the secretion of IAPP is decreased in non-insulin-dependent diabetics (Hanabusa et al. 1992).
Nevertheless, the morphometrical finding seems to argue against the notion that more IAPP is produced and released in diabetes. In fact, if one of the many functions of IAPP is to suppress the secretion of insulin possibly by a paracrine route (Tokuyama et al. 1997), a reduction of IAPP in the islets of diabetics where the number of insulin cells are reduced is an expected condition.
In agreement with previous studies (Madsen et al. 1991; De Vroede et al. 1992), IAPP was also found in a few non-β-cells. In diabetics, however, more somatostatin cells coexpressed IAPP than in non-diabetic cases (Figure 1D).
The increase of glucagon and somatostatin cells in diabetic cases was much greater than reported elsewhere (Clark et al. 1995). In our cases, the increase of the α- and δ-cells in diabetic cases was 40% and 58%, respectively. This was higher than in the non-diabetic cases. Comparing the size of the islets and the number of islet cells, the increase appears to be absolute. However, adequate tissue sampling as reported by Stefan et al. (1982) is required for confirmation. An increase of somatostatin and glucagon cells in diabetics has also been shown in previous studies along with the demonstration of the potent inhibitory effect of somatostatin on insulin and glucagon release (Orci et al. 1976; Gerich 1981; Yoon et al. 2003). Hyperplasia and hypertrophy of somatostatin cells has been documented in the islets of chronic juvenile-type diabetes (Orci et al. 1976) and has been regarded as a compensatory response to the excessive glucagon secretion of diabetes (Corbett et al. 1997; Bonner-Weir 2001). The predominant expression of IAPP in the δ cells in experimental diabetes (De Vroede et al. 1992) and in our cases further complicates the understanding of the interaction between individual pancreatic endocrine cells in controlling glucose metabolism.
Marked hyperplasia of centroacinar cells found only in diabetic cases was noteworthy. The strong reactivity of these cells with the anti-IAPP that we used has not been reported with any other monoclonal anti-IAAP antibodies (Ma et al. 1997,2000) or polyclonal antibody (Rocken et al. 1992). Presently, we do not know if the reactivity of the hyperplastic centroacinar cells to the antibody is specific. If so, then the increased IAPP in diabetics is explained. Different monoclonal and polyclonal IAPP antibodies show different reactivity (Ma et al. 1997,2000); however, further studies with other monoclonal and polyclonal antibodies are required for a better understanding of the issue. Nevertheless, the presence of cells reactive with antibodies to islet hormones within the hyperplastic centroacinar cells supports our previous findings that centroacinar cells present islet progenitor cells (Pour 2006). Although it is generally accepted that islet cells derive from ductal epithelium, in diabetes, centroacinar cells seem to be the primary cells for β-cell regeneration.
The present study warrants further morphometrical and clinical studies in diabetic subjects to better understand the role of IAPP in this disease. In such studies, attention should be paid to the expression of IAPP in extrapancreatic tissues because the coexpression of IAPP has been found in the intestinal endocrine cells of animals (Mulder et al. 1994).
Footnotes
Acknowledgements
This study was supported by grants from the National Cancer Institute's Laboratory Cancer Research Center (Support Grant CA-367127) and the American Cancer Society Special Institutional Grant.