
Abstract
This study first investigated the ontogeny of Islet-1 and neuronal nitric oxide synthase (nNOS) expression and their co-localization in the DRG of sheep fetuses during gestation by immunohistochemistry (IHC). The results showed that Islet-1 and nNOS were located in the nuclei and cytoplasm of DRG neurons, respectively. The relative percentages of Islet-1-immunopositive (Islet-1+) neurons accounting for the total DRG neurons were 90%, 79%, 66%, and 53% at days 60, 90, and 120 of gestation and postnatally, respectively. The percentage of nNOS-immunopositive (nNOS+) neurons was 94% at day 60 and declined to ∼30% at day 90, with no obvious further change until the postnatal period. Dual IHC showed that ∼69% Islet-1+ neurons express nNOS at day 60 of gestation. This proportion declined to ∼24% at day 90, after which there was no significant change until birth. We also observed that most Islet-1+ and nNOS+ neurons belonged to small and medium-sized DRG neurons from day 90 of gestation to the postnatal period. These results suggest that both Islet-1 and nNOS are important for the differentiation and maintenance of some specific phenotypes of DRG neurons during late gestation of sheep fetuses, although the related mechanisms need to be further elucidated. (
I
It has been well documented that nNOS, acting as an essential synthase for the regulation of nitric oxide (NO), has a variety of functions in the development of the nervous system. nNOS is a calcium/calmodulin-dependent enzyme stimulated by activation of NMDA-type glutamate receptors (Garthwaite et al. 1988; Bredt and Snyder 1990). NO signaling occurs primarily through activation of soluble isoforms of guanylyl cyclase, and the cellular effects of NO are the results of an NO-induced increase in the level of cGMP (Jaffrey and Snyder 1995). In the peripheral nervous system, NO plays an important role in nociceptive signaling (Moulton 1996; Robbins and Grisham 1997; Wallace and Chin 1997). nNOS is mainly expressed in small and medium-sized nociceptive DRG neurons in rat and monkey (Zhang et al. 1993; Qian et al. 1996). During fetal development, many DRG neurons express nNOS in the early stage, while most of these become nNOS-negative in the late stage in rat (Wetts and Vaughn 1993) and chick (Ward et al. 1994), especially when the axons extend peripherally and start to take up nerve growth factor (Thippeswamy and Morris 2002).
Both Islet-1 and nNOS are expressed and play important roles in developing DRG, but to date there are no data describing the ontogenic patterns of Islet-1 and nNOS expression and their relationships in DRG of sheep fetuses. The aim of this study was to determine the ontogeny of Islet-1 and nNOS expression by IHC in developing DRG of sheep fetuses during gestation.
Materials and Methods
Animals and Tissue Collection
After the animal use and experimental design for this study was approved by Chinese Association for Laboratory Animal Sciences, pregnant Chinese Xiaowei ewes with known single insemination dates were anaesthetized with an overdose of sodium pentobarbital on day 60, 90, or 120 of gestation (term = 147 days). The fetuses were exposed by caesarean surgery. The DRG at the level of lumbar vertebrae 6–7 (L6, L7) were collected from sheep fetuses at days 60 (n = 6), 90 (n = 6), and 120 (n = 6) of gestation and from newborn lambs (n = 6) by surgery, fixed in 4% paraformaldehyde in PBS, pH 7.4, for 12 hr and then embedded in paraffin. Five-μm serial sections were then cut for IHC detection.
Immunohistochemistry
Immunocytochemical localization of Islet-1 was performed using a mouse monoclonal antibody (Mab) against Islet-1 (40.2D6, Developmental Studies Hybridoma Bank), which had been used to detect Islet-1 in chicken (Avivi and Goldstein 1999; Cui and Goldstein 2000), mouse (Kubota et al. 1999), and rat (Yamamoto and Henderson 1999). Comparison of the sequence data of Islet-1 between sheep (Chen and Cui, unpublished data) and mouse showed 98% homology. Immunocytochemistry for Islet-1 detection was performed as described previously (Avivi and Goldstein 1999; Cui and Goldstein 2000). Briefly, the sections were dewaxed and rehydrated, and antigen retrieval was performed by microwaving the sections for 20 min (four times for 5 min) at full power in 0.01 M sodium citrate buffer (pH 6.0). Endogenous peroxidase was blocked by incubating the sections in 3% H2O2 in methanol for 30 min. After nonspecific binding was blocked with 10% normal sheep serum in PBS, the sections were then incubated with the Islet-1 antibody (diluted 1:50) overnight at 4C. After further washing in PBS, the sections were incubated with sheep anti-mouse IgG (Serotec, Poole, UK; 1:50) for 2 hr at room temperature (RT). Mouse peroxidase-anti-peroxidase (Scottish Antibody Product Unit, Scotland, UK; 1:50) was then added for 2 hr, and the peroxidase activity was detected with diaminobenzidine (Sigma; St Louis, MO). Finally, the sections were counter-stained with hematoxylin and mounted using conventional methods.
nNOS was detected with a rabbit anti-nNOS polyclonal antibody (Upstate Biotechnology, Lake Placid, NY; 1:40) which had been used to detect nNOS in sheep (Peterson et al. 2001). nNOS detection procedures were similar to that for Islet-1, with the exception that the primary, secondary, and tertiary antibodies were rabbit anti-nNOS polyclonal antibody (Upstate Biotechnology; 1:40), biotinylated swine anti-rabbit IgG (DAKO, Glostrup, Denmark; 1:300), and avidin–biotin complex (Vector Laboratories; Burlingame, CA).
For dual labeling of Islet-1 and nNOS, the primary antibodies were same as those for the single staining. The sections were incubated with a mixture of anti-Islet-1 MAb (1:50) and anti-nNOS polyclonal antibody (1:40) overnight at 4C. The slides were then rinsed in PBS three times and incubated in a mixture of sheep anti-mouse IgG (Serotec; 1:50) and biotinylated swine anti-rabbit IgG (DAKO; 1:300) for 2 hr at RT. The mixture of mouse alkaline phosphatase–anti-alkaline phosphatase (DAKO; 1:30), and avidin–biotin complex (Vector) was then added for 2 hr. After three further washes in PBS, the alkaline phosphatase was visualized by the addition of NBT/BCIP for 10 min and the color reaction was stopped by washing in water. The peroxidase was visualized by the addition of diaminobenzidine (Sigma) for 2 min.
DRG cell proliferation was detected with the same procedures as that for Islet-1 detection except that the primary antibody was replaced by a mouse anti-proliferating cell nuclear antigen (PCNA) MAb (Zymed, S. San Francisco, CA; 1:200).
The controls included replacement of anti-Islet-1 and anti-PCNA Mabs and of anti-nNOS polyclonal antibody by normal mouse serum and normal rabbit serum, respectively, and by the omission of the primary, secondary, and tertiary antibodies, all of which eliminated staining (data not shown).
Data Analysis
From day 60 of gestation, DRG neuron profiles could be distinguished. In all cases, only the neurons that had distinguishable nuclei were observed and counted under a high-magnification objective with an eyepiece graticule. In each specimen, five fields were selected randomly throughout the DRG and the number of neurons immunopositive for Islet-1 or nNOS were counted and expressed as a percentage of the total number of neurons in that field, while the dual staining neurons were expressed as a percentage accounting for the total number of Islet-1+ or nNOS+ neurons, respectively. Results were presented as mean ± SEM. The differences between means of the repeated measures (n = 6) over time at different stages of sheep fetuses were tested for significance using ANOVA Student's t-test.
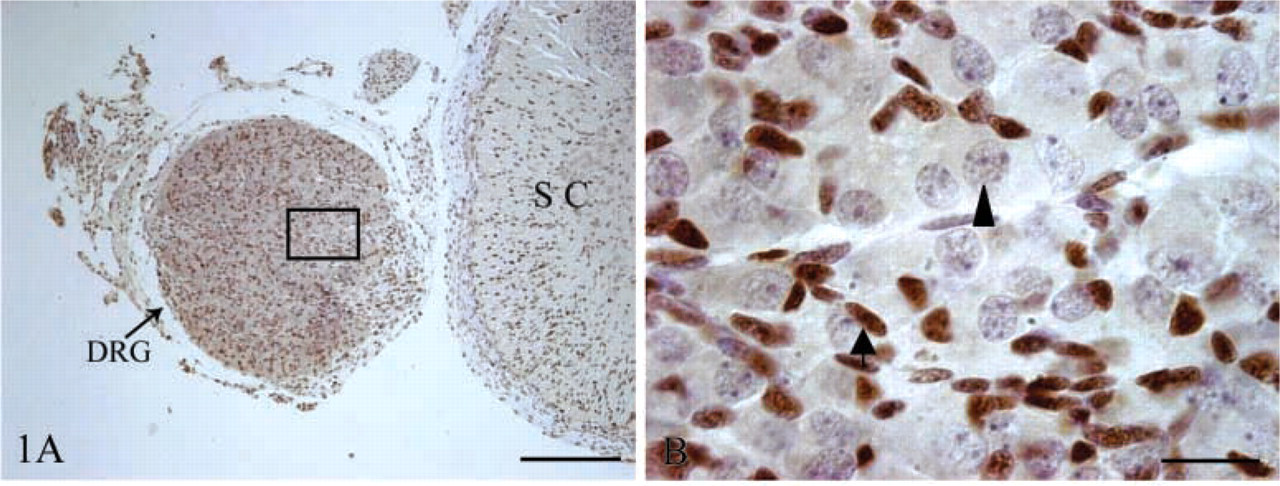
PCNA IHC staining of sheep fetal DRG at day 60 of gestation. Neurons are indicated with an arrowhead and glia with an arrow. (
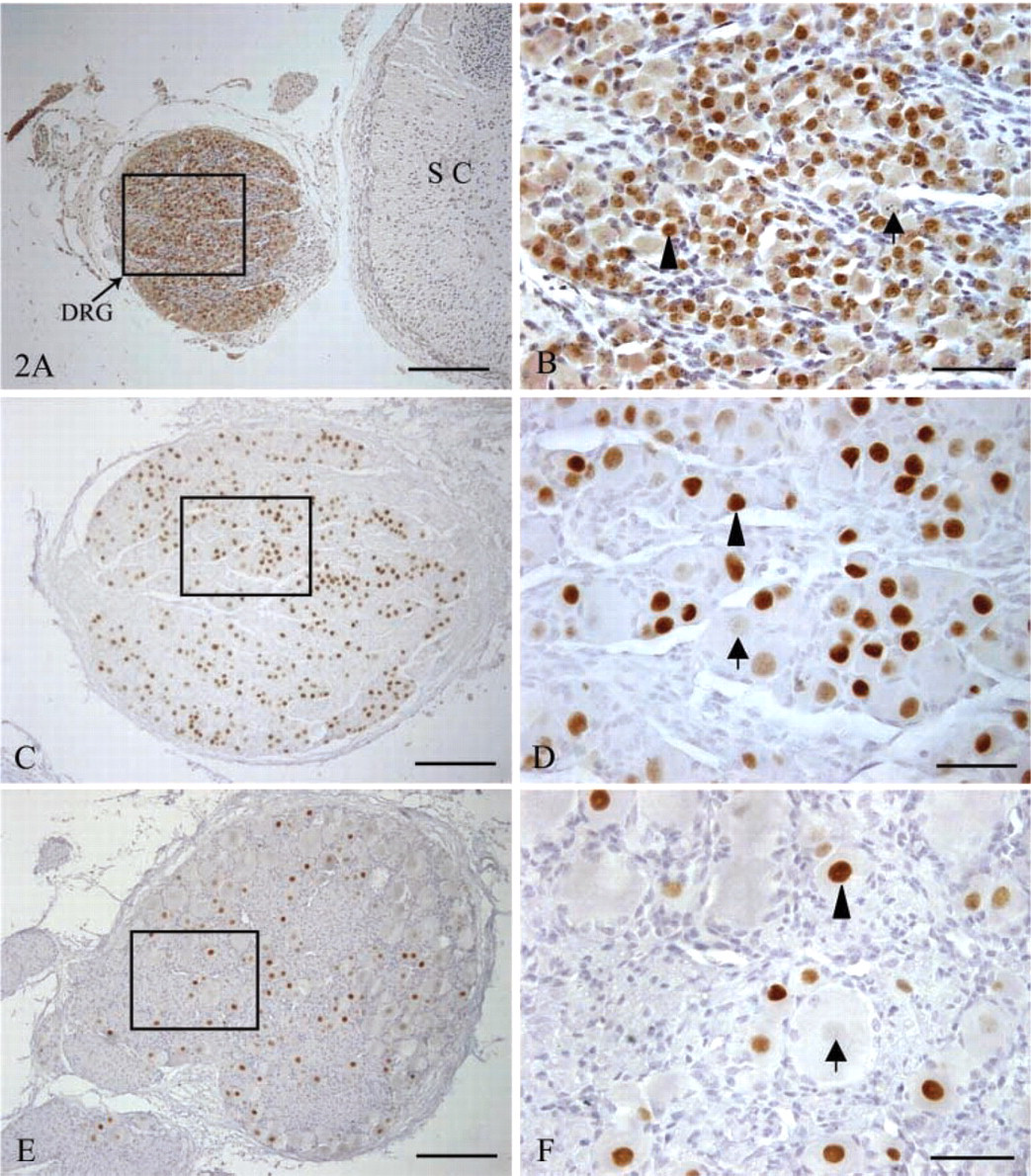
Islet-1 IHC staining of sheep fetal DRG at days 60, 90, and 120 of gestation. Positive cells are indicated with an arrowhead and negative cells are identified by an arrow. (
Results
We did not find any PCNA-immunopositive neurons in the DRG at day 60 of gestation (Figures 1A and 1B), whereas Islet-1 was located in nuclei of DRG neurons and Islet-1+ neurons were evenly distributed within the DRG for the duration of this study. At day 60 of gestation, there was no evident difference in size and morphology of DRG neurons (Figures 2A and 2B). By day 90 of gestation, DRG neurons were of different sizes and the number of Islet-1+ neurons obviously decreased (Figures 2C and 2D). This difference in cell morphology was very clear. Most Islet-1+ neurons were of small and medium size, although a few large neurons were Islet-1-immunopositive at day 120 of gestation (Figures 2E and 2F). Islet-1 staining was homogeneous in DRG, and this pattern persisted through the postnatal period. The cell counting results showed that the percentages of Islet-1+ neurons accounting for the total DRG neurons at days 60, 90, and 120 of gestation as well as postnatally were 90.33 ± 1.09%, 78.94 ± 2.68%, 66.08 ± 4.45%, and 53.14 ± 5.66% respectively (Figure 5A). These results demonstrated that there was a significant decrease (p<0.05) in the number of Islet-1+ neurons from day 60 to day 120 with no significant change at birth.
The immunochemical detection of nNOS expression showed that nNOS was localized in the cytoplasm of DRG neurons in sheep fetuses throughout gestation (Figure 3). The percentages of nNOS+ neurons accounting for total DRG neurons at the various gestational ages were 93.88 ± 2.04%, 29.07 ± 3.09%, 32.7 ± 2.08% and 30.58 ± 2.71%, respectively (Figure 5B). These indicated that the proportions of DRG neurons expressing nNOS did not change markedly after day 90 of gestation, even if it was significantly (p<0.05) higher at day 60 of gestation.
Dual ICC results viewed at high magnification showed that ∼60% of DRG neurons expressed Islet-1 and nNOS at day 60 of gestation (Figure 4A), whereas only a subset of small and medium-sized DRG neurons continued to express Islet-1 and nNOS from day 90 of gestation to the postnatal period (Figures 4B–4D). The proportions of Islet-1+ neurons possessing nNOS at the various gestational ages are show in Figure 6B. This proportion decreased sharply (p<0.01) from day 60 (69.2 ± 3.0%) to day 90 (23.9 ± 3.6%) of gestation, after which there was no significant change until birth, whereas the proportions of nNOS+ neurons possessing Islet-1 at days 60, 90, and 120 of gestation and at birth were 69.3 ± 4.5%, 45.55 ± 8.06%, 39.6 ± 4.8%, and 40.3 ± 4.5%, respectively (Figure 6A).
Discussion
The present study was the first IHC detection of DRG Islet-1 and nNOS expression in sheep fetuses, although Islet-1 or nNOS immunoreactivity had been detected in the DRG of chick (Ward et al. 1994; Avivi and Goldstein 1999; Cui and Goldstein 2000; Avivi et al. 2002), mouse, and rat embryos (Chen and Chiu 1992; Ericson et al. 1992; Wetts and Vaughn 1993). In fact, much less is known about the developmental course of DRG structure and function in sheep fetal life. This study showed that the DRG neuron profile could be distinguished by microscopy at day 60 of gestation and that there was no evident difference in the morphology of DRG neurons. In addition, our results confirmed the reports that all DRG neurons were negative for PCNA immunoreactivity (Waseem and Lane 1990; Cardoso et al. 1993) at day 60 of gestation, and over 90% DRG neurons were positive for Islet-1 IHC staining. These findings demonstrate that the DRG neurons of sheep fetuses were just out of the cell cycle and of differentiation at day 60 of gestation. In addition, these data support the reports that Islet-1 is expressed shortly after the final mitosis of sensory neurons and that its expression is closely related to the differentiation of DRG neurons (Chen and Chiu 1992; Ericson et al. 1992; Cui and Goldstein 2000), especially related to determination of peripheral axon extension and targeting (Tsuchida et al. 1994; Daston and Koester 1996; Thor and Thomas 1997). By day 90 of gestation, the DRG neurons showed differences in morphology and size, and these differences became more evident from day 90 of gestation to birth, although we did not quantify these data. For the duration of gestation, the proportion of Islet-1+ neurons accounting for total DRG neurons continued to decline, and most Islet-1+ neurons were small or medium-sized. In addition, it has been reported that the DRG neurons of sheep fetuses began to function from day 75 of gestation (Rees et al. 1994,1995) and that the small and medium-sized DRG neurons were responsible for the nociceptive properties (Zhang et al. 1993). Therefore, Islet-1 expression is probably involved in the maintenance of the specific phenotypes of DRG sensory neurons during fetal development.
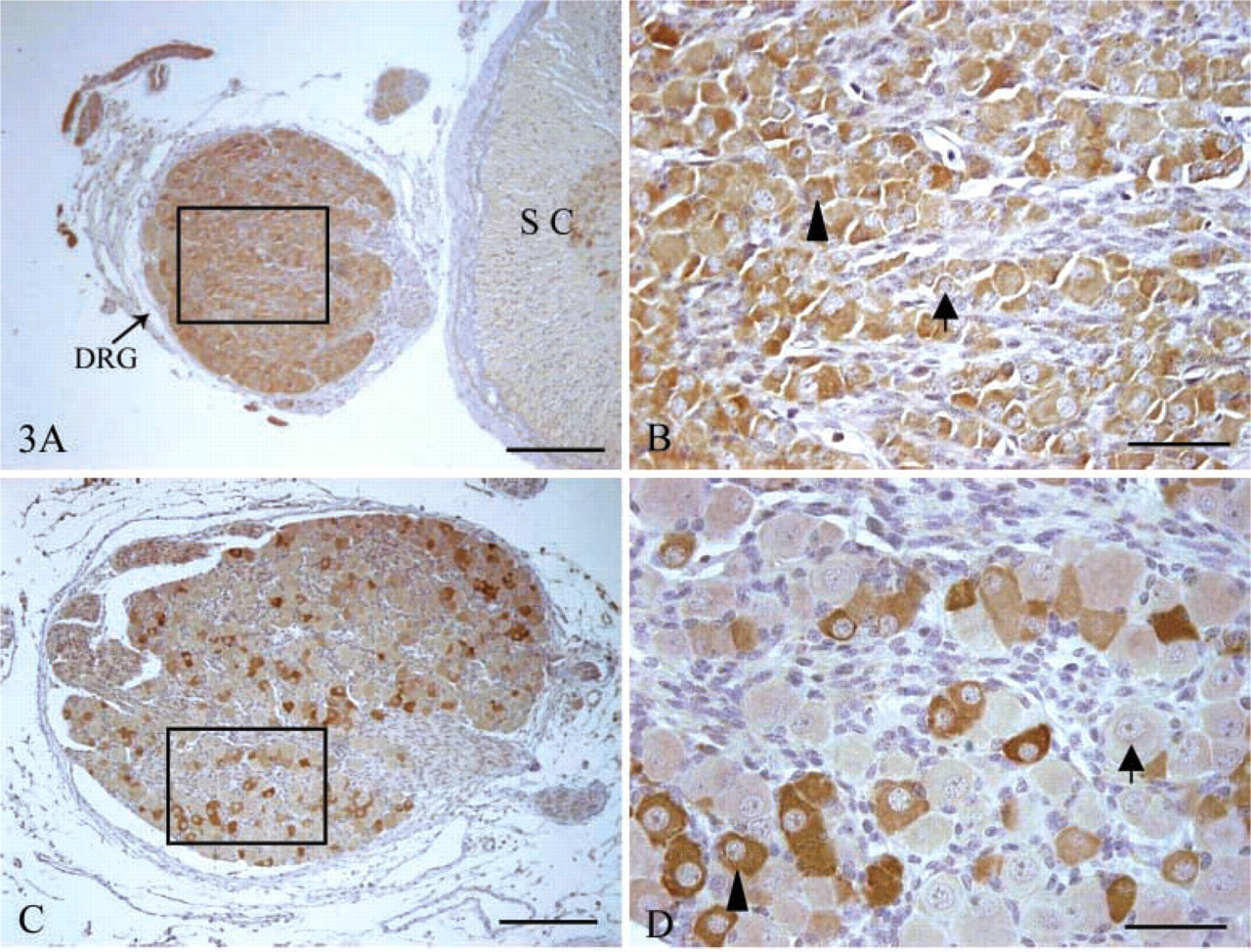
IHC of nNOS in sheep fetal DRG obtained at day 60 and 90 of gestation. Positive cells are indicated with an arrowhead and negative cells are identified by an arrow. (
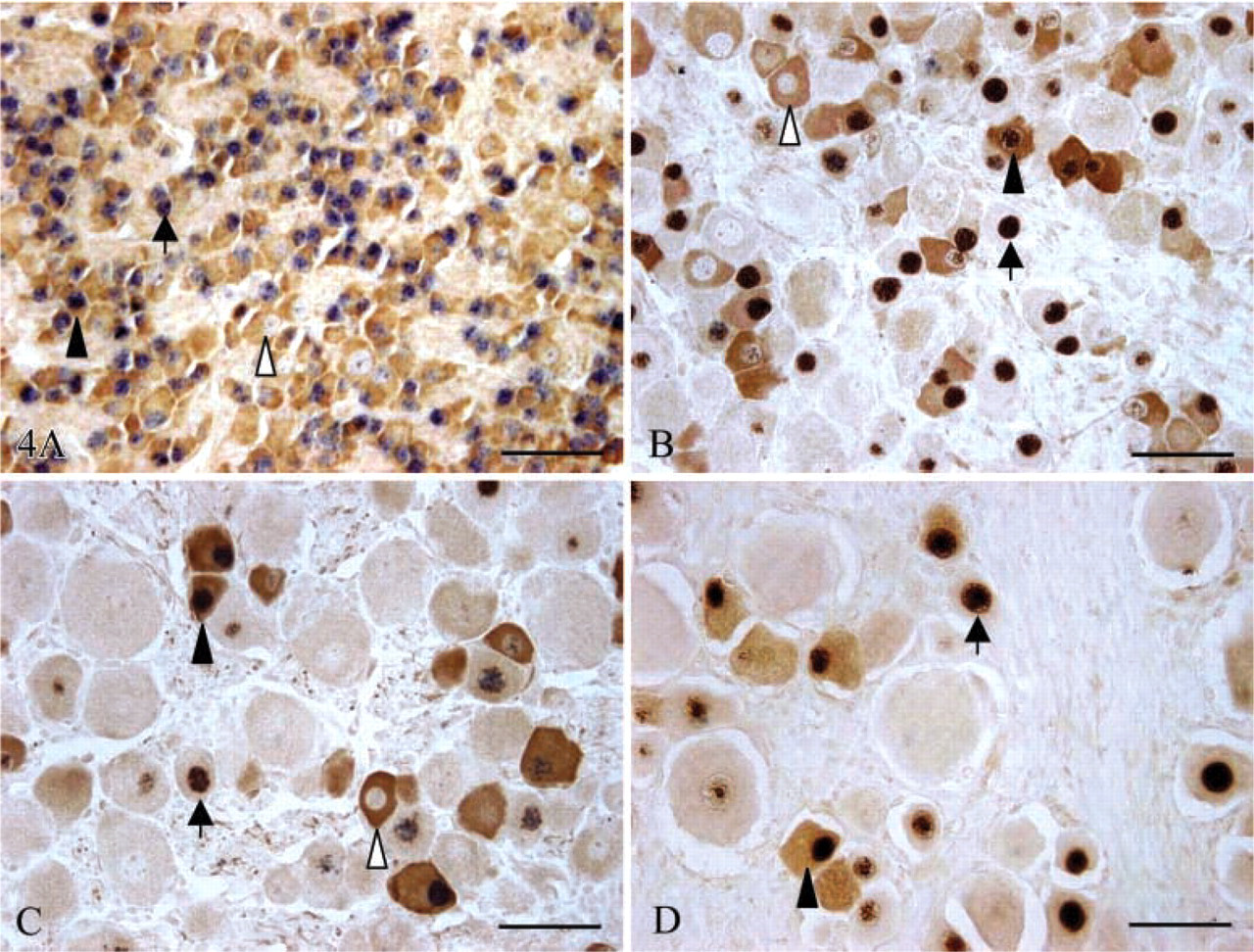
Co-expression of Islet-1 and nNOS in sheep fetal DRG at days 60 (
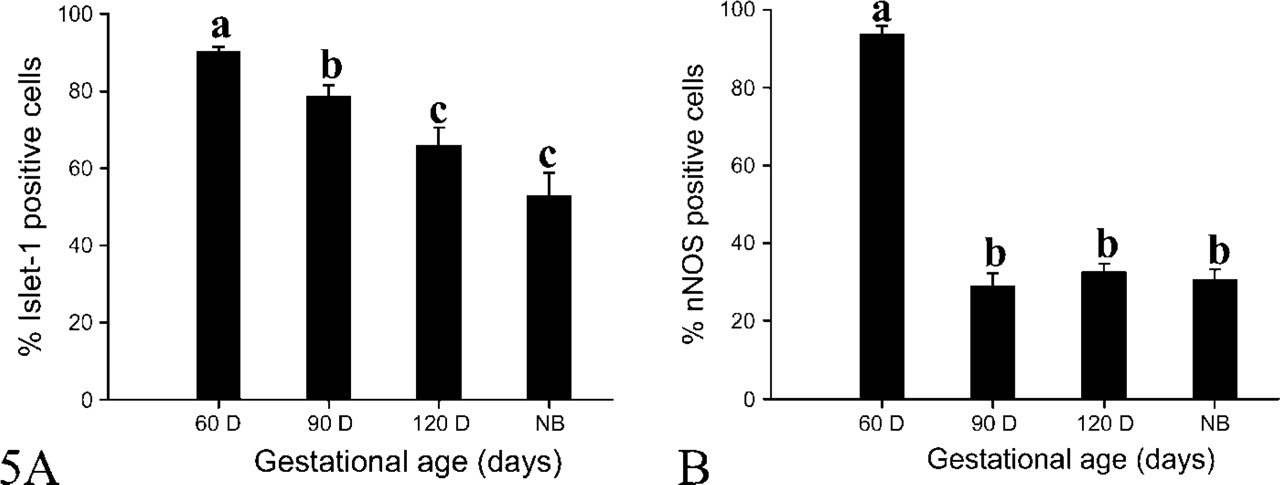
The percentages of Islet-1 (
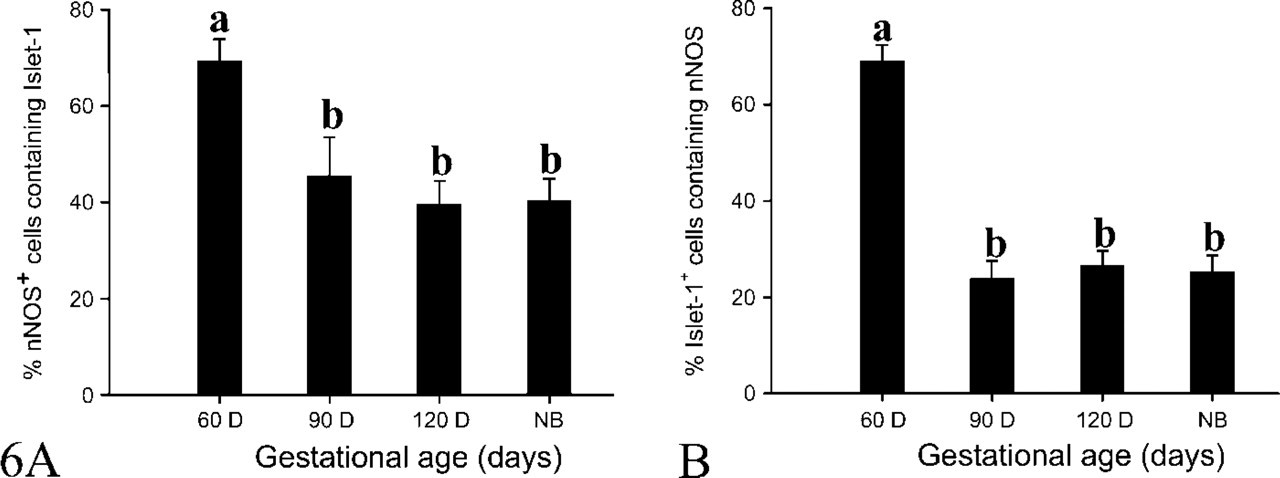
The percentages of dual staining neurons accounting for nNOS+ (
Similar to the Islet-1 expression, nNOS immunoreactivity was detected in over 90% of DRG neurons of the sheep fetuses at day 60 of gestation, but this proportion sharply declined to around 30% at day 90 of gestation and persisted at that level until birth. This changing pattern of nNOS expression is in accordance with that in rat (Wetts and Vaughn 1993) and chick embryo (Ward et al. 1994). Although the relationship of nNOS expression to DRG development and functional regulation remains to be further investigated, the present results support the reports that nNOS expression is important for the differentiation of DRG neurons, especially in determining the developmental direction of DRG neurons by regulating the synthesis of NO in the early stage of gestation (Robbins and Grisham 1997; Thippeswamy and Morris 2002). However, from the middle stage of gestation until birth the relative proportion of nNOS-immunopositive neurons was around 30% accounting for the DRG neurons, and sheep sensory innervation of skin and muscle occurred from day 75 of gestation (Rees et al. 1994,1995). During this period, many DRG neurons extend peripherally (Rees et al. 1994,1995) and begin to take up nerve growth factor (NGF) (Thippeswamy and Morris 2002), accompanied by depression of nNOS expression (Wetts and Vaughn 1993; Ward et al. 1994). Furthermore, most nNOS-immunopositive neurons were small and medium-sized DRG neurons. These observations suggest that nNOS expression may be associated with the functional maintenance of some nociceptive DRG neurons.
It has been documented that Islet-1 functions mainly through interactions with other proteins to regulate the transcription of some genes relative to development (Chen and Chiu 1992; Ericson et al. 1992; Dawid et al. 1995; Hobert and Westphal 2000), whereas nNOS regulates the synthesis of NO, which acts as a molecular signal to induce an increase in intracellular cGMP levels and triggers a series of cellular effects that include enhancement of neurotrophin-induced neurite outgrowth and prevention of neuron apoptosis (Farinelli et al. 1996; Hindley et al. 1997; Thippeswamy and Morris 2002). The dual IHC staining results first demonstrated that Islet-1 and nNOS were co-expressed in more than 60% of DRG neurons at day 60 of gestation. This implies that Islet-1 may have cooperative effects with nNOS or NO in DRG development. In addition, after the middle stage of gestation only a subset of DRG neurons co-expressed Islet-1 and nNOS. These were mainly small and medium-sized neurons that were mainly related to nociceptive properties (Zhang et al. 1993; Qian et al. 1996). This suggests that Islet-1 and NO may interact with each other to maintain some functions of a subset of DRG neurons after the middle stage of gestation, supported by reports that the expression of nNOS was upregulated (Verge et al. 1992; Fiallos-Estrada et al. 1993; Zhang et al. 1993) and Islet-1 was downregulated in the DRG during peripheral nerve regeneration (Pfaff et al. 1996; Hol et al. 1999). However, further study is needed to determine the functions and the related mechanisms of Islet-1 and nNOS in the development and functional regulation of the nervous system.
Acknowledgments
Supported by grants from the Natural Science Foundation for Outstanding Young Scientists of China (30325034) and by the Science and Technology Key Program (01018) of the China Educational Ministry.
The Islet-1 monoclonal antibody 40.2D6 was developed by Thomas Jessel and was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by the University of Iowa, Department of Biological Sciences, Iowa City, IA.