
Abstract
The injured growth plate cartilage is often repaired by bony tissue, resulting in impaired bone growth in children. Bone morphogenic proteins (BMPs) are important for bone fracture repair, and as a step to characterize potential involvement of BMPs in bony repair of injured growth plate, expression of BMPs and receptors (BMP-R) was examined by quantitative RT-PCR and immunohistochemistry in rat injured tibial growth plate. During the inflammatory response on day 1, slightly increased expression of BMP-3, BMP-4, BMP-R1a, and BMP-R2 was observed, with immunostaining seen among inflammatory cells at the injury site. During mesenchymal infiltration and osteogenic responses on days 3-14, moderately increased expression of BMP-2, −3, −4, −7, and BMP-R1a was found, with immunostaining observed among infiltrated mesenchymal cells and differentiated osteoblasts lining bony trabeculae. During maturation phase on days 14-25, only BMP-7 was seen upregulated slightly and was localized in osteoblasts and marrow cells at the injury site. The temporospatial expression of BMPs and receptors at the injured growth plate suggests potential involvement of BMP-3 and −4 in regulating the inflammatory response or as its mediators in modulating downstream events, and BMP-2, −3, −4, and −7 in the fibrogenic and osteogenic responses, and BMP-7 in bone remodeling at the injured growth plate.
T
However, because the growth plate is the most fragile structure in a child's developing long bones, trauma injury to the growth plate is a common occurrence, and fractures to the growth plate occur when the mechanical loading placed on the long bone exceeds the mechanical strength of the growth plate (Iannotti 1990). Growth plate injuries, particularly of Salter's types III and type IV that involve the entire vertical destruction of the growth plate, can cause significant problems for a developing long bone. The growth plate cartilage has limited ability to regenerate, and these two types of injuries that are common in young children are often repaired by bony tissue at the injury site, a process often called bone bridge formation (Xian et al. 2004). Bone bridge formation at the injured growth plate results in structural disorganization of the growth plate and disruption of endochondral longitudinal bone growth (Xian et al. 2004), causing significant skeletal problems such as limb length discrepancy and angulation deformity.
Cellular and molecular mechanisms for the bony repair of the injured growth plate remain unclear. Previously, using a growth plate drill-hole injury model in young rats that causes a Salter's type IV-like defect, Xian et al. (2004) observed sequential injury responses leading to the bony bridge formation at the injury site, namely, the inflammatory, fibrogenic, and osteogenic responses and the bony bridge remodeling/maturation. Typically after the injury, inflammation occurs with infiltration of inflammatory cells peaking on day 1 and declining on day 3. From days 3 to 7, infiltration and proliferation of mesenchymal cells occurs at the growth plate injury site, and some of these mesenchymal cells differentiate into osteoblasts (Xian et al. 2004). From day 7 onwards, bony trabeculae form and enlarge at the injury site. During days 14-25, bony bridge is matured with formation of bone marrow between bone trabeculae. Although there was a previous study reporting upregulated expression of some cytokines or growth factors at these various phases of bony repair (Zhou et al. 2004), how these cellular healing responses are regulated remains largely unknown.
Because bone morphogenic proteins (BMPs) are important regulators for bone formation and repair, it can be hypothesized that BMPs might be involved in regulating growth plate injury responses and bony repair. BMPs are a large group of developmental regulatory factors that belong to the TGF-β super family, with BMP-2, −3, −4, −5, −6, and −7 being important members in skeletal development acting on mesenchymal cells to induce cartilage and bone formation (Zoricic et al. 2003). BMPs elicit their responses through binding and activating their membrane-bound cell surface receptors, type-I (BMP-R 1a, BMP-R 1b) and type-II (BMP-R2, ACVR-2a, and ACVR-2b) receptors (Onishi et al. 1998), which activate down-stream signaling transducing Smad molecules that translocate to the nucleus and activate gene transcription (Kloen et al. 2003). BMPs have been shown to stimulate proliferation, migration, and differentiation of mesenchymal cells into osteogenic and chondrogenic lineages (Sakou 1998). BMPs, particularly BMP-2, −4, and −6, control the rate of proliferation and chondrocyte maturation and stimulate collagen production within longitudinal bone growth (De Luca et al. 2001). BMPs are also strong regulators of bone formation as a result of their potential roles in mediating the migration of progenitor cells, proliferation of mesenchymal cells, differentiation of osteogenic cells, and bone remodeling. BMPs such as BMP-2 and −7 induce osteoblast differentiation of mesenchymal cells and stimulate mineralized bone matrix formation (Canalis et al. 2003; Cheng et al. 2003; Shea et al. 2003). Furthermore, BMPs have also been identified as important regulators for bone fracture repair involved in the stages of endochondral formation (cartilage formation) and intramembranous bone formation, followed by bone formation and remodeling (Gerstenfeld et al. 2003).
Although BMPs are important regulators for bone formation and bone fracture healing, their potential roles in the bony repair of injured growth plate cartilage remain unknown. As a step to investigate their potential involvement in regulating the various injury responses leading to bony repair at the injured growth plate, this study examined the expression of BMPs and their receptors through quantitative real-time RT-PCR and immunohistochemical (IHC) analysis in a rat growth plate injury model.
Materials and Methods
Tissues Specimens, RNA Samples, and cDNA Synthesis
RNA samples and paraffin histology sections used in the current study were obtained from previous studies in which a drill-hole injury (with ~35% injury area) to the proximal tibial growth plate was inflicted in 6-week-old male Sprague-Dawley rats (Xian et al. 2004; Zhou et al. 2004). For collection of tissue specimens, rats were sacrificed on days 1, 3, 7, 14, and 25 postoperatively, with the growth plate cartilage collected from the right proximal tibia and stored at −80C for RNA extraction, and the left proximal tibia collected, fixed in 10% formalin, decalcified in 14% EDTA, and processed for paraffin embedding for histological and IHC studies (Xian et al. 2004). Total RNA from each growth plate specimen was isolated by grinding the sample in liquid nitrogen followed by extraction in RNAwiz RNA isolation solution (Ambion; Vic, Australia) and purification using RNeasy mini-column (Qiagen; Vic, Australia) with the on-column DNase digestion step to remove any genomic DNA contaminations (Qiagen) as described (Zhou et al. 2004). Due to small amounts of RNA that can be obtained from the growth plate cartilage and the many numbers of genes (multiple BMPs and receptors) that need to be examined, RNA samples were pooled proportionally from the four animals per each time point group to give a total of 5 μg for cDNA synthesis using random decamers (Geneworks; SA, Australia) and StrataScript reverse transcriptase (Stratagene; NSW, Australia) in a total of 60-μl reaction following manufacturer's instructions.
Real-time Quantitative PCR Gene Expression Analysis
Primer sets for rat BMP-2, −3, −4, −6, −7, BMP-R 1a, BMP-R2, and ACVR-2a were designed using NCBI DNA sequences and Primer Express software (Applied Biosystems; NSW, Australia). Several possible primer sets of each specific gene were used to search the NCBI nucleotide database for sequence homology, specificity, and expected PCR product using BlastN software, and the primer sets selected for the specific target genes are tabulated in Table 1. Cyclophilin-A was selected as the endogenous internal control standard for normalization of target gene expression. It was previously found to be suitable for growth plate injury expression study because its expression levels are ideal for analysis of low abundant genes (Zhou et al. 2004). Primers were ordered from Invitrogen (NSW, Australia).
Primers used in this study for quantitative real-time RT-PCR analysis of bone morphogenic proteins (BMPs) and BMP receptors (BMP-R1a, BMP-R2, ACV-R2a)
SYBR green quantitation for each specific gene was performed in parallel with cyclophilin A in triplicate. Each 20-μl reaction of PCR gene amplification consisted of 1 μl (50 ng) of cDNA, 1.5 μl of forward primer, 1.5 μl of reverse primer, 6 μl of sterile water, and 10 μl of SYBR green quantitation mix. Primer concentrations were set at 225 and 75 nM, respectively, for cyclophilin-A and target genes after prior primer optimization assays to avoid primer dimerization. PCR reactions were carried out using a RotorGene quantitative real-time PCR machine (Corbett Research; NSW, Australia) following reaction parameters as described (Zhou et al. 2004). An amplification curve of the triplicate reaction was generated through SYBR quantitation in Rotor Gene 5 software, and relative gene expression was calculated using the 2-ΔΔ CT method, where threshold cycle (CT) values from triplicate runs of each gene were averaged and calibrated in relation to cyclophilin CT values of the individual samples. Levels of gene expression in injured samples are presented as fold change in relation to normal uninjured samples. Gel electrophoresis in 2% agarose followed by staining in ethidium bromide was performed to visualize and identify the RT-PCR product of appropriate size from the reactions together with a 100-bp ladder (Geneworks). Because the RT-PCR data were generated from pooled RNA samples, data are presented as means of individual pooled samples (without SEM); thus, no statistical analysis was carried out. Sample pooling from the same group is generally an acceptable practice as reported in many studies in the literature, particularly for specimens of small sizes such as growth plate cartilage from rodents.
IHC Analysis of BMPs and Receptors
IHC was conducted to examine and localize specific BMPs (BMP-2, −3, −4, and −7) and their receptor (BMPR-1a) at the injured growth plate. Five-μm tissue sections were obtained from the paraffin specimens collected from the rat growth plate injury trial (Xian et al. 2004). Tissue sections were first deparaffinized and then quenched in 3% H2O2 in PBS, pH 7, for 30 min to remove endogenous peroxidase activity. Sections were then heated in 0.01 M citrate buffer at 70C for 90 min as an antigen retrieval step. After blocking in 10% normal serum and 1% BSA in PBS, sections were incubated at 4C overnight with primary antibodies against BMP-2 (sc-6895, 1:100), BMP-3 (sc-9031, 1:100), BMP-4 (sc-6896, 1:100) (all 0.2 mg/ml IgG PBS solutions; Santa Cruz Biotechnology, Santa Cruz, CA), BMP-7 (1:400; provided by Dr. David Ruger; Stryker Biotech, Hopkinton, MA), and BMP-R1a (1:2000; kindly provided by Dr. D. Baird; Edinburgh, UK) (Souza et al. 2002). After washing in PBS, sections were incubated with secondary biotin-conjugated secondary antibodies (1:600) and ABC complex reagents (1:400), followed by color development in Liquid DAB Plus substrate as instructed (Dako Australia; Botany, NSW, Australia). Sections were thoroughly washed and placed in Mayer's hematoxylin counterstaining solution prior to deydration and coverslipping. As negative controls, addition of normal IgG (of the same dilution) or 1% BSA/PBS instead of a primary antibody on tissue sections in the above immunostaining procedure was performed, which gave negative staining.
Results
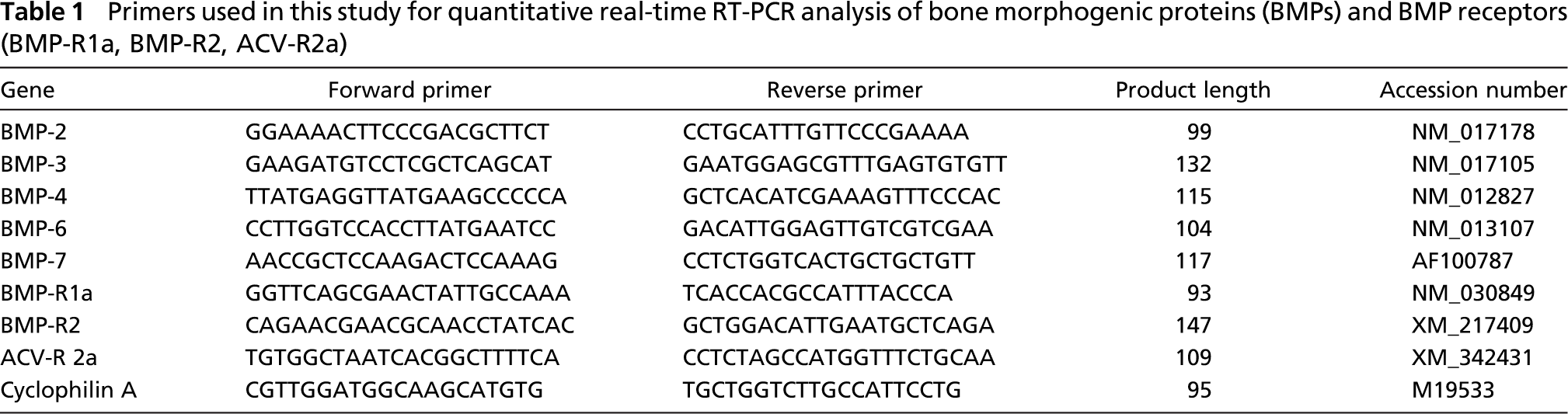
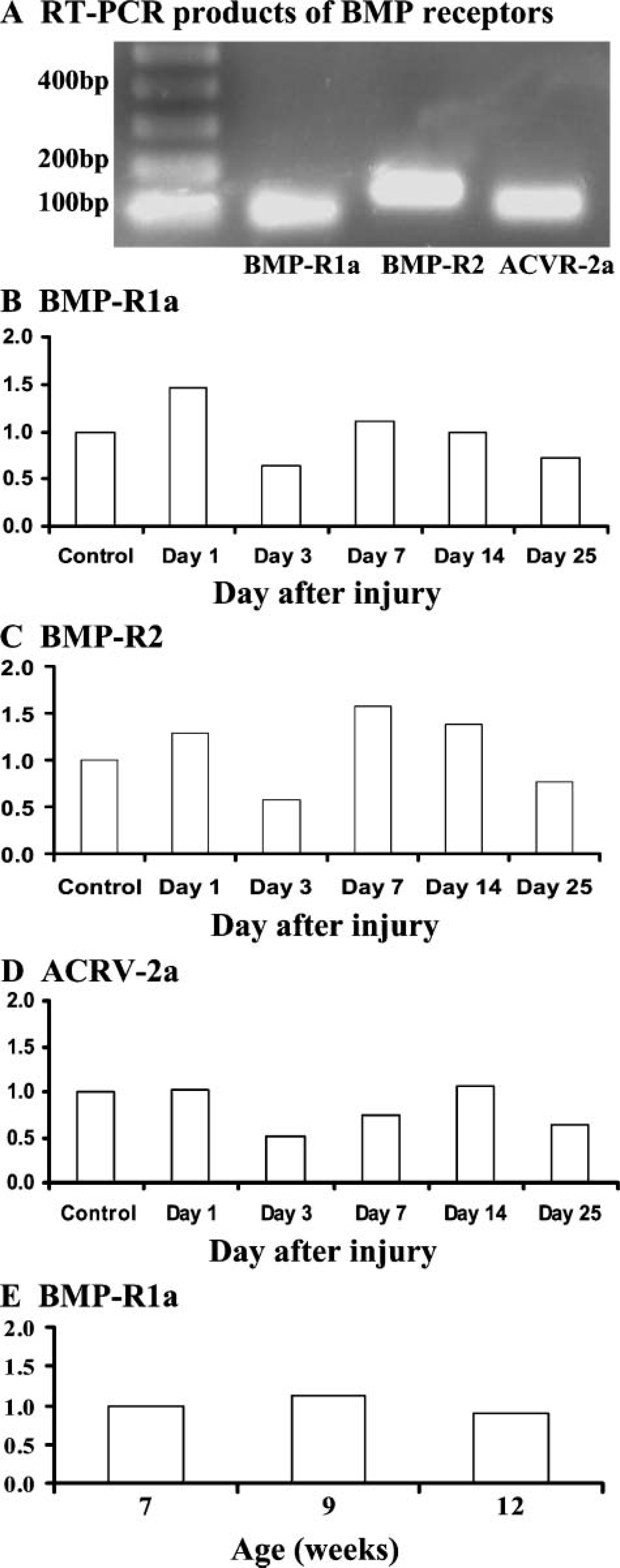
Quantitative real-time RT-PCR analysis of mRNA expression of BMPs and their receptors was conducted, and expression levels of BMPs (Figure 1) and receptors (Figure 2) at different injury time points are presented as fold change compared with non-injury normal controls after being normalized to expression levels of cyclophilin-A of individual samples. Gel electrophoresis of RT-PCR reaction products reveal single-band products of ~100-bp expected size (as predicted in Table 1), suggesting specificity of the RT-PCR reactions (Figure 1A and Figure 2A). In addition, IHC analysis was performed to localize expression at the injury site of molecules identified by RT-PCR having upregulated mRNA expression. Positive immunostaining of BMP-2, −3, −4, −7, and BMP-R1a was found present in the proliferative and hypertrophic chondrocytes at the normal growth plate (Figure 3A), in the osteoblasts lining the bone trabeculae at the normal metaphysis (Figure 3B), and in the fibrous mesenchymal cells in the inner cambium layer of the periosteum (data not shown). As negative controls for immunostaining, a staining procedure with the addition of the normal IgG or 1% BSA/PBS instead of primary antibodies was used, which gave negative staining (Figure 3N).
BMP mRNA expression levels at the injured growth plate as determined by quantitative real-time RT-PCR analysis.
Expression of BMPs
Although BMP-2 mRNA expression remained unchanged during the initial inflammatory response at day 1, it was downregulated at day 3 at the end of the inflammatory phase and the beginning of the fibrogenic response and was increased back to normal levels by day 7 at the beginning of osteogenic response (Figure 1B), suggesting its potential role in initiating bone formation. However, BMP-2 expression declined on later time points during bone bridge maturation and remodeling on days 14 and 25. Consistently, at the inflammatory response at day 1, no positive BMP-2 staining was observed among the inflammatory cells (data not shown). At day 3 at the beginning of the fibrogenic response, limited BMP-2-positive staining was observed in mesenchymal cells at the growth plate injury site (Figure 3C). At day 7, many mesenchymal cells and osteoblasts were abundantly stained positive for BMP-2, including osteoblasts lining the newly formed bone trabeculae (Figure 3D). By day 14 when the newly formed bone trabeculae matured, some staining, but to a much lesser extent, of BMP-2 was observed in the bone cells and in mesenchymal cells in the bone marrow within the injury site (data not shown). Therefore, abundant immunostaining of BMP-2 at day 7 at the beginning of the osteogenic response suggests that BMP-2 may play a role in stimulating osteoblast differentiation and bone formation at the injured growth plate.
Expression of BMP-3 mRNA (Figure 1C) was slightly upregulated during the inflammatory response on day 1 but declined on day 3 at the beginning of the fibrogenic response. However, BMP-3 levels increased on day 14 during the osteogenic and remodeling phase (Figure 1C). Interestingly, BMP-3 immunostaining was observed in the inflammatory cells (most likely neutrophils based on nuclear morphology) on day 1 (Figure 3E). Positive staining of BMP-3 was also observed in some bone marrow-derived infiltrating mesenchymal cells at the beginning of the fibrogenic response at day 3 (Figure 3F), in mesenchymal cells and differentiated osteoblasts surrounding the newly formed bone during the osteogenic response at day 7 (Figure 3G), and during bone bridge maturation at day 14 (Figure 3H). These findings suggest that BMP-3 may regulate early inflammation phase and bone formation/maturation at the growth plate injury site.
As shown in Figure 1D, BMP-4 mRNA expression was slightly upregulated on days 1 and 3 at the inflammatory and fibrogenic responses, followed by a small decrease by day 7. However, its levels at day 14 during the bone bridge formation/maturation were again slightly upregulated compared with non-injury control and returned toward normal levels during the bone bridge remodeling stage on day 25. Positive BMP-4 staining on inflammatory cells was also observed on day 1 at the injury site (Figure 3I). Similar to BMP-3, BMP-4 immunostaining was present in some infiltrating mesenchymal cells at day 3 (Figure 3J) and in some mesenchymal cells and lining osteoblasts on day 7 (data not shown). By day 14 when bony bridge is being remodeled, BMP-4 staining, to a much lesser extent, was seen in only some bone-lining cells and some mesenchymal cells (data not shown). These findings indicate that BMP-4 may potentially be involved in regulating the initial inflammatory and fibrogenic responses as well as bone cell differentiation, bone formation, and bone bridge remodeling in the injured growth plate.
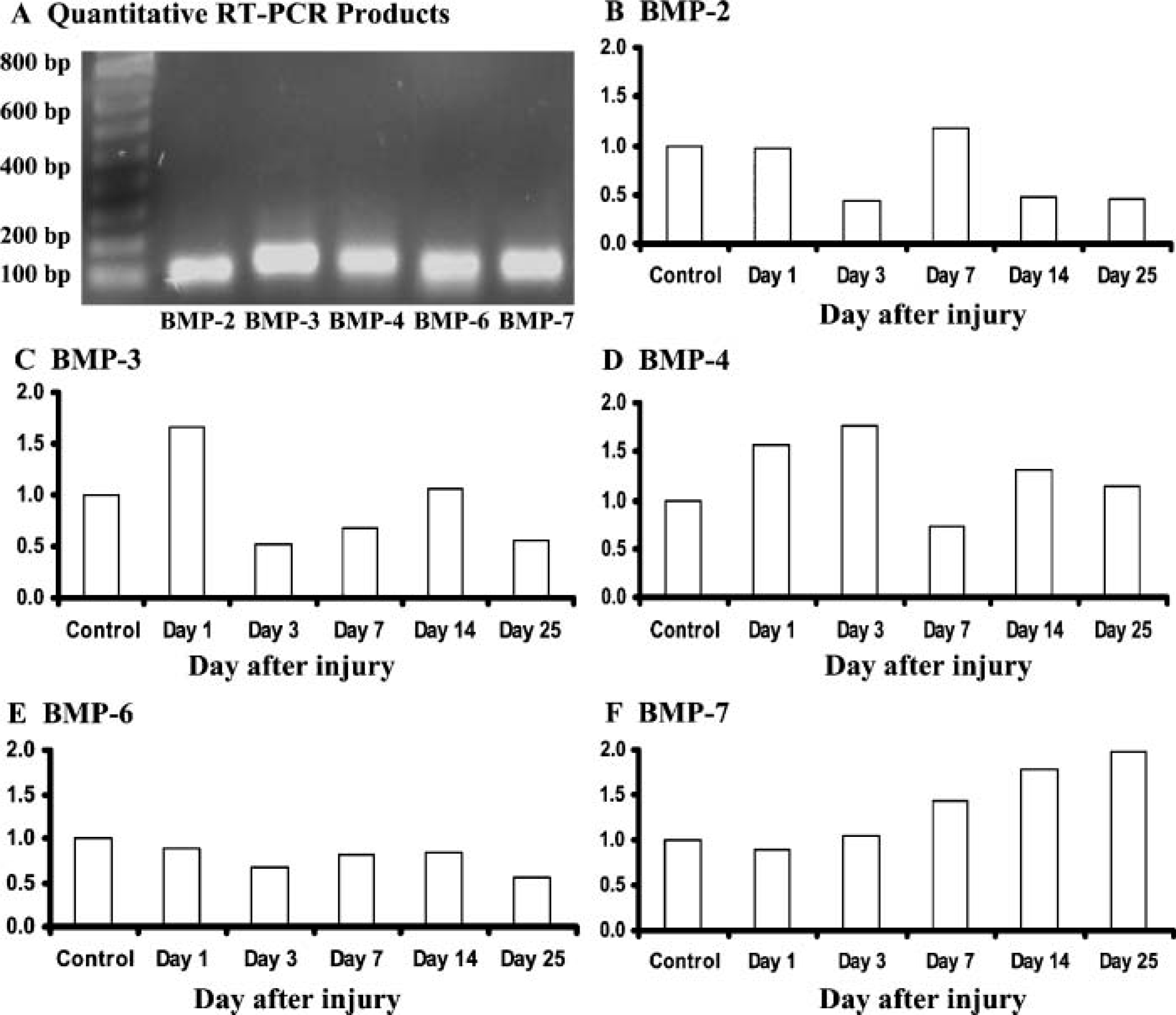
BMP receptor mRNA expression levels at the normal or injured growth plate as determined by quantitative real-time RT-PCR analysis.
BMP-7 mRNA levels remained unchanged during the inflammatory response on day 1 and at the beginning of fibrogenic response on day 3 (Figure 1F). Consistently, limited positive BMP-7 immunostaining was observed among inflammatory cells (data not shown), and by day 3 BMP-7-positive staining was seen in only some infiltrated bone marrow-derived mesenchymal cells at the growth plate injury site (data not shown). Interestingly, mRNA levels of BMP-7 were upregulated gradually from day 7 to day 25 during the entire bone bridge formation and maturation phases. Prominent BMP-7 immunostaining was seen among osteoblasts, bone-lining cells, and some mesenchymal cells on day 7 (Figure 3K). By day 25 when the bony bridge is matured with extensive bone marrow formed between the bone trabeculae, positive BMP-7 immunostaining was present in bone cells lining the bone trabeculae and in many bone marrow cells (predominantly of hematopoietic cells lineage as judged by morphology) (Figure 3L). Interestingly, expression of other BMPs examined (BMP-2, −3, and −4) display much less immunostaining in bone marrow (data not shown). Increased expression levels and positive expression pattern of BMP-7 in primitive mesenchymal cells, bone-lining cells, osteoblasts, and bone marrow cells suggest that BMP-7, a strong promoter for bone formation and remodeling during bone fracture healing, may potentially be important in regulating bony bridge formation and remodeling at the injured growth plate.
Unlike other BMPs examined, mRNA expression levels of BMP-6 remained unchanged after growth plate injury, indicating that BMP-6 may have limited activities in regulating cellular events at the growth plate injury site.
Expression of BMP-Rs
Slightly increased mRNA levels of BMP-R 1a (Figure 2B) and BMP-R2 (Figure 2C) were observed during the inflammatory response on day 1, and some inflammatory cells were seen displaying positive BMP-R 1a immunostaining (data not shown). Although mRNA levels of both BMP-R 1a and BMP-R2 declined by day 3, they returned to normal or slightly higher levels during the osteogenic response on days 7 and 14 and returned to normal levels by day 25. On day 3, BMP-R 1a immunostaining was present in some infiltrated mesenchymal cells at the injury site (Figure 3M), and by day 7, some mesenchymal cells, differentiated osteoblasts, and bone-lining cells were immunopositive for BMP-R 1a (data not shown). Expression levels of BMP receptor-2 ACRV-2a remained unchanged during the inflammatory response on day 1, declined slightly on day 3, and returned to normal on day 14 during the osteogenic phase (Figure 2D). Increased levels of expression and positive localization of BMP-R1 and -R2 at the injury site again suggest the potential roles of BMP signaling pathways in regulating the inflammatory response, bone cell differentiation, and bone formation at the growth plate injury site.
Age Effects on Expression of BMPs and BMP-Rs
To determine whether the changes in mRNA expression levels of BMPs and their receptors were due to the injury responses at the injured growth plate but not due to the age differences of the rats between different time points after surgery, gene expression analysis was conducted similarly for BMP-2 and BMP-R 1a in RNA samples from normal rat tibial growth plate of 7, 9, and 12 weeks of age, which corresponded to day 0 (normal non-injured controls), day 14, and day 35 after injury. This age effect expression analysis showed no obvious changes in levels of BMP-2 and BMP-R 1a mRNA expression (Figure 2E), showing data for BMP-R 1a only, suggesting that during this 5-week study period, BMP-2 and BMP-R 1a expression remained relatively constant in the normal growth plate, and that the changes in mRNA expression levels of BMPs and receptors observed after the growth plate injury most probably resulted from the injury responses at the growth plate.
Discussion
Bony bridge formation at the growth plate injury site has been previously shown to involve the inflammatory, fibrogenic, and osteogenic responses and the bony bridge remodeling maturation, mainly through the mechanism of intramembranous ossification (Xian et al. 2004). BMPs and their receptors are well known to have roles in regulating osteogenesis during bone fracture repair. In this study, to examine the potential involvement and possible roles of BMPs and their receptors in the injury-induced inflammatory, fibrogenic, osteogenic, and remodeling responses in the bony repair of the growth plate injury site, expression of some important members of BMPs and receptors were examined by quantitative real-time RT-PCR analysis and IHC at time points shown to be appropriate to study the injury-induced various responses in a rat model (Xian et al. 2004; Zhou et al. 2004).
The inflammatory response at the injured growth plate has been previously shown to be rapid and transient, with infiltration of inflammatory cells peaking at day 1 and subsiding on day 3 after injury (Xian et al. 2004). The current study has revealed upregulated expression of BMP-3, −4, and their receptor BMP-R 1a in the injured growth plate and detected positive immunostaining of these three molecules among the inflammatory cells at the injury site. Previously, the injured growth plate has been shown to have upregulated expression of TNF-α, IL-1β, IGF-I, and TGF-β (Zhou et al. 2004) during the inflammatory response. Therefore, together with these inflammation-associated molecules, BMP-3 and BMP-4 could be involved in modulating the inflammatory response and/or the downstream healing responses at the injured growth plate. Similarly, early after bone fracture, BMP-4 mRNA was shown induced at the fracture site prior to cartilage/bone callas formation (Nakase et al. 1994), and infiltrated inflammatory cells have been shown to deposit BMP-4 at the bone fracture injury site (Schierbel et al. 2001). Interestingly, BMPs have been suggested to induce chemotaxis of monocytes to the site of fracture and stimulate expression of osteogenesis-promoting growth factors to enhance bone formation (Sykaras and Opperman 2003). For example, BMP-4 increased production of the vascular endothelial growth factor that stimulates migration of endothelial cells for vascular formation during acute inflammation (Ten Dijke et al. 2003). In addition, BMP-4 produced by inflammatory cells at the bone fracture injury site may play a role in regulating mesenchymal cell migration and differentiation (Schierbel et al. 2001). Therefore, the initial inflammatory response after bone or growth plate fracture may be an important multipotential modulator and initiator of downstream healing responses, and BMPs could be a mediator of the inflammatory response in regulating the downstream healing events.
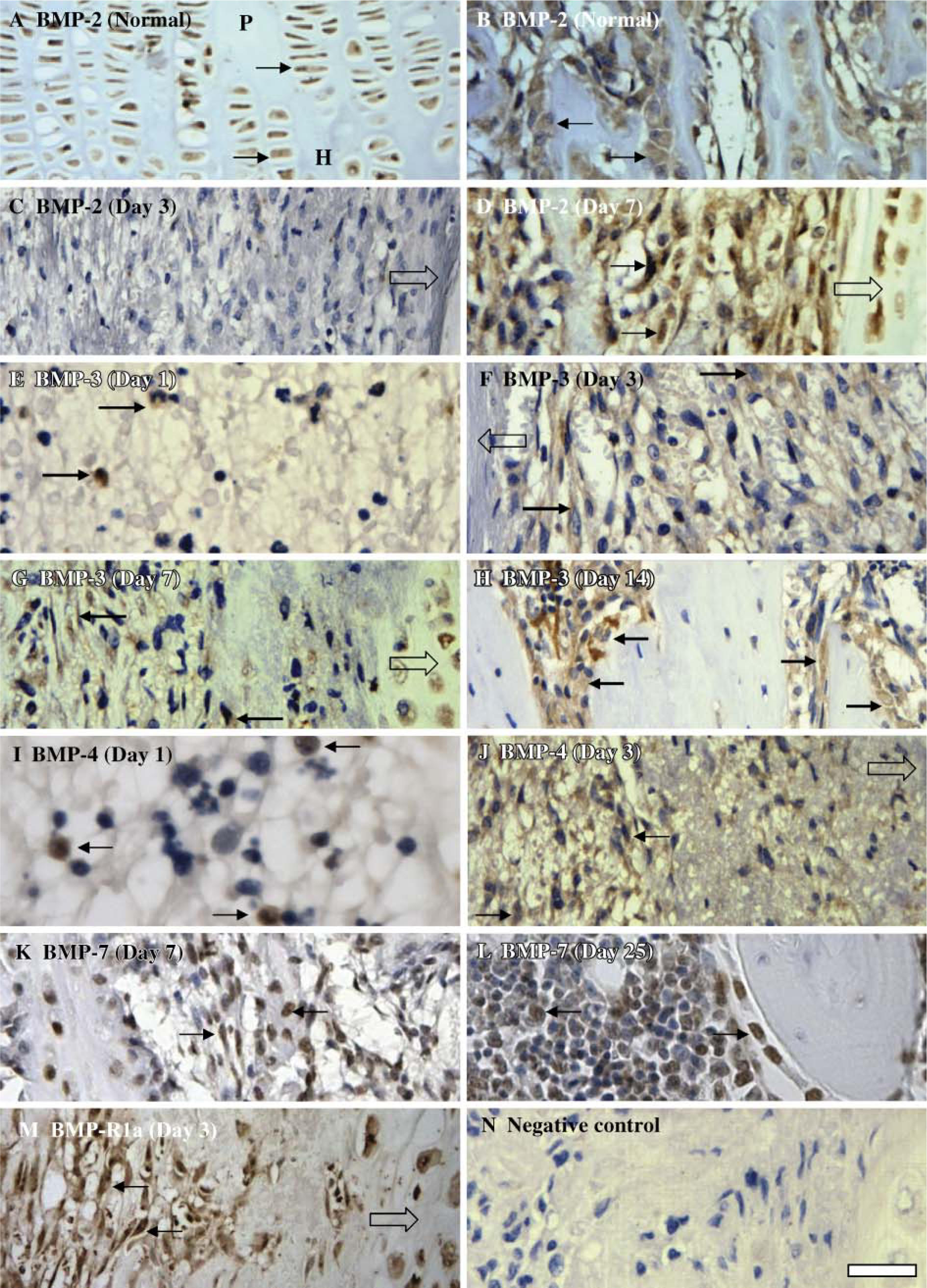
Immunostaining of BMPs and receptor BMP-R 1a in the normal or injured growth plate at various time points.
After the inflammatory response, fibrogenic response is the next dominant healing response that is characterized by the migration/condensation of marrow-derived mesenchymal cells into the growth plate injury site commencing at day 3 and peaking by day 7 (Xian et al. 2004). Subsequently, starting at about day 7, bone cell differentiation from some of these mesenchymal cells and bone formation occur at the injured growth plate (Xian et al. 2004). Previous bone fracture healing studies have suggested that migration and proliferation of primitive mesenchymal cells to the site of injury, their osteogenic differentiation, and bone formation are regulated by the signaling pathways of BMPs and their receptors (Ten Dijke et al. 2003). In the current study, mRNA levels of BMP-2, −3, −4, and −7 were found increased at the injured growth plate on days 3, 7, and/or 14 during the fibrogenic and osteogenic responses, and many infiltrated mesenchymal cells, differentiated osteoblasts, and cells lining the bony trabeculae at the growth plate injury site displayed positive immunoreactivity, suggesting that these BMPs may be involved in regulating mesenchymal cell migration, bone cell differentiation, and/or bone formation at the injured growth plate.
Bone bridge maturation and remodeling stage (from day 14 to day 25 after injury) at this rat growth plate injury model is characterized by new bone formation by osteoblasts and bone resorption by osteoclasts as well as formation of bone marrow (Xian et al. 2004). During this bone bridge maturation phase, mRNA expression of all BMPs analyzed in the current study was shown to have returned to normal levels except for BMP-7. BMP-7 showed upregulated mRNA expression on days 14 and 25, and BMP-7 immunostaining was seen predominantly in differentiated osteoblasts and some bone marrow cells at the growth plate injury site. These findings suggest that BMP-7 may potentially be involved in bone remodeling in this growth plate injury bony repair model. Interestingly, a previous study has suggested that BMP-7 may play a more important role in stimulating osteogenesis producing osteocalcin in mature osteoblasts than in inducing osteoblast differentiation of mesenchymal stem cells (Cheng et al. 2003). In addition, BMP-7 is known to promote and induce bone formation and bone remodeling at ectopic fracture sites (Canalis et al. 2003) and to induce osteoclast differentiation, maturation, and bone remodeling activity in newly formed bone trabeculae during fracture repair (Onishi et al. 1998). Evidence from previous studies and our current study suggests that BMP-7 may regulate bone resorption as well as bone formation during bony repair of the injured growth plate. Although BMPs, particularly BMP-7, have been suggested to play a role in regulating hematopoietic stem cell differentiation (Martinovic et al. 2004), functional roles of BMP-7 expression in bone marrow hematopoietic cells remain unknown.
Upon BMPs stimulation, BMP-R2 and BMP-R 1a form a heterodimer complex to elicit biological responses (Wan and Cao 2005). Actions of BMPs through BMP-R signaling pathway have been shown to activate transcriptional factor cbfa-1 expression, which is critical to induce osteoblast differentiation (Canalis et al. 2003). During rat bone fracture healing, expression of BMP-R 1a and BMP-R2 was detected during mesenchymal cell migration, proliferation, and differentiation as well as in bone formation and remodeling stages (Onishi et al. 1998). In the current study, BMP-R 1a and BMP-R2 mRNA levels showed similar expression trends throughout the growth plate injury time course (i.e., slightly increased expression during the inflammatory response and bone formation), suggesting that both receptors may act in concert to elicit a BMP response in this injury and repair model. In addition, this study showed localization of BMP-R 1a in mesenchymal cell condensation, osteoblasts and bone-lining cells, which further suggests possible involvement of BMP signaling pathway in osteogenesis during bony repair of the injured growth plate. Limited expression of ACVR-2a but with a higher expression of BMP-R2 in this study indicate that BMP-R2 may be the primary type-II receptor that interacts with type-I receptor to elicit a BMP response at the bony repair and injured growth plate.
Numerous studies have indicated that BMPs play important roles in bone fracture healing. For example, BMP-2 was suggested to be an early response gene after fracture (Cho et al. 2002) and was a critical mediator of bone marrow-derived mesenchymal cell migration and osteoblast differentiation potently promoting bone formation and was shown upregulated during callus formation in fracture repair (Bostrom et al. 1995; Canalis et al. 2003; Cheng et al. 2003). In contrast, BMP-3, BMP-4, BMP-7, and BMP-8 showed a restricted period of expression when the resorption of calcified cartilage and osteoblastic recruitment was most active (Cho et al. 2002). During bone fracture healing, infiltrated mesenchymal cells and, later on, differentiated osteoblasts were shown to be immunopositive for BMP-4 at the fracture site (Bostrom et al. 1995; Onishi et al. 1998), and BMP-4 has been found to stimulate differentiation of osteoprogenitor cells toward the osteoblastic lineage (Cheng et al. 2003) and to promote osteogenesis within bone fracture sites (Sakou 1998). BMP-7 was found upregulated at the bone fracture sites (Sakou 1998) and stimulated osteoblast differentiation, osteogenesis, and remodeling (Yeh et al. 2002). Findings from the current study suggest that BMPs might have similar roles in regulating bony repair at the injured growth plate as in regulating bone fracture healing.
In the current study, expression data on the mRNA level by RT-PCR and on the tissue protein level by IHC appear consistent for some BMPs at certain time points (e.g., BMP-2 at day 7) but appear having disparities for others (e.g., BMP-3). Several factors could potentially contribute to these apparent disparities. First, because the RT-PCR data were generated with RNA samples isolated from the whole growth plate, they represent the sum of changes within the injury site (~35% in area) and those in the surrounding uninjured growth plate area. On the other hand, our non-quantitative IHC analysis localized changes in BMP immunoreactivity only within the injury site. Second, due to reasons such as protein secretion and time differences in mRNA transcription and protein translation, it is not surprising that levels of protein vs mRNA expressions in tissues may not necessarily be consistent in some physiological and pathophysiological conditions. For example, we have previously observed that whereas the intestinal trefoil (TFF3) was deficient on the protein level, its mRNA level was upregulated in the small intestine with mucositis damage as a side effect from chemotherapy treatment (Xian et al. 1999).
In summary, the current study showed induction and localization of BMP-3, BMP-4, BMP-R 1a, and BMP-R2 at the growth plate injury-induced inflammatory response, suggesting potential involvement of BMP-3 and −4 in regulating the inflammatory response itself or being a mediator of inflammatory response in regulating downstream healing events. This study has also identified the localization and expression of BMP-2, BMP-3, BMP-4, BMP-7, and BMP-R 1a in infiltrated mesenchymal cells and osteoblasts during subsequent fibrogenic and osteogenic responses, suggesting their potential roles in modulating these two healing responses. At the bone bridge maturation stage, only BMP-7 expression was seen slightly upregulated and expressed in differentiated osteoblasts and bone marrow cells at the injury site. Therefore, this study reveals differential temporospatial expression of individual BMPs and their receptors in various stages of bony repair of the injured growth plate, suggesting their potential roles in regulating the cellular events leading to bony bridge formation and remodeling at the injured growth plate.
Further studies are required for confirming these potential roles and elucidating the action mechanisms of BMPs during the growth plate injury-induced responses, particularly BMP-3 and BMP-4 at the inflammatory and osteogenic responses and BMP-7 in bone bridge maturation. In addition, RT-PCR data were from RNA samples of the pulverized entire growth plate and thus represent the sum of changes within the injury site and from the surrounding uninjured growth plate. In this study, although we have used IHC to localize changes in BMP and BMP-R immunoreactivity within the injury site, one future study would be to use laser-capture dissecting technique in combination with sensitive gene expression analysis tools to analyze gene expressions separately in the injured site and in the surrounding uninjured growth plate cartilage. Perhaps such discriminating analysis would reveal more obvious changes in levels of expression of these genes within the injury site as opposed to the small changes seen in the pulverized whole growth plate as observed in the current study.
Footnotes
Acknowledgements
The project was funded by Bone Growth Foundation (BGF) and in part by grants from Channel 7 Children's Research Foundation of South Australia and from the Australian National Health and Medical Research Council.
The authors thank Rosa McCarty, Jo Cool, and Carmen Macsai for technical advice.