
Abstract
Our aim was to investigate and to compare the intracellular distribution of ubiquitin, 20S proteasome, and all six proteasomal regulatory ATPases in non-diseased human and rat brains. Ubiquitin and ATPases S4 and S7 show dominant nuclear immunostaining, whereas subunits S6a, S6b, and S10b show mainly cytoplasmic immunostaining in both species. However, S8 localization is inconsistent, prevailing nuclear in rat and cytoplasmic in human. In rat brain, small clastosome-like nuclear bodies demonstrate strong ubiquitin, 20S, and S6a immunoreactivity both in neurons and glial cells. Prominent nuclear immunolocalization of members of the ubiquitin-proteasome system provides morphological evidence for function of these proteins in transcription regulation and/or DNA repair.
T
Immunolocalization of UPS, primarily in cell cultures, is well established and summarized in reviews (Gordon 2002). However, there is a paucity of systematic comparative studies on the cellular localization of regulatory ATPases in mammalian brain samples. Here we demonstrate the subcellular distribution of ubiquitin, 20S PS, and the six regulatory ATPases in non-diseased human and rat brains.
Ten human brains were selected (six males and four females, mean age: 65.1 years, mean postmortem delay: 16 hr, cause of death: four due to heart failure, four due to respiratory failure, and two due to pneumonia). All brains underwent neuropathological evaluation to exclude alterations. Light microscopic immunohistochemistry was performed on sections of formalin-fixed, paraffin-embedded tissue blocks from the hippocampus, temporal isocortex, neocerebellum, and medulla oblongata. In parallel, eight 7-week-old male Dark-Agouti (DA) rats were perfused transcardially with Zamboni fixative [4% paraformaldehyde (w/v%), 15% saturated picric acid (v/v%) in 0.1 M phosphate buffer], and brains were embedded into paraffin. We analyzed cingulate, frontal, and pyriform cortex, lateral septal nucleus, caudate putamen, globus pallidus, hippocampal CA3 and granular layer of the dentate gyrus, medial thalamus, anterior hypothalamus, lateral hypothalamic area, dorsal-medial raphe, and cerebellar Purkinje cells. Animals were handled in accordance with the guidelines of the National Research Council. In case of human brains, procedures were in accordance with the Helsinki Declaration of 1975.
Antibodies and pretreatment methods are listed in Table 1. Applying Western blotting analysis and appropriate technical controls of immunostaining on human brain samples, we have recently demonstrated that antibodies against the different proteasome subunits were specific and suitable for immunohistochemical studies (Ádori et al. 2005). For light microscopic immunohistochemistry, immunostaining was developed with a peroxidase/DAB kit (En Vision; DAKO, Glostrup, Denmark). No first antibody and preimmune serum controls were performed.
Antibodies and epitope retrieval methods
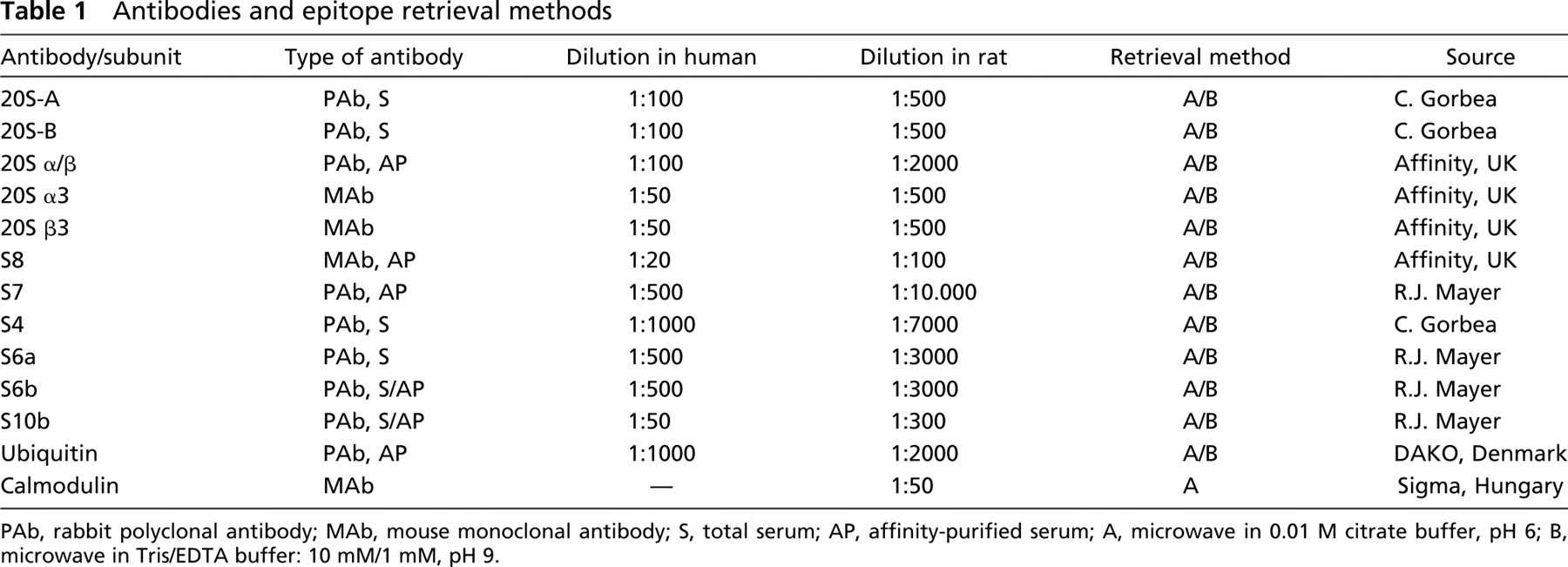
PAb, rabbit polyclonal antibody; MAb, mouse monoclonal antibody; S, total serum; AP, affinity-purified serum; A, microwave in 0.01 M citrate buffer, pH 6; B, microwave in Tris/EDTA buffer: 10 mM/1 mM, pH 9.
For double immunolabeling, the fluorescent-labeled secondary antibody for anti-ubiquitin was Alexa Fluor 488 goat anti-rabbit IgG (1:200; Molecular Probes, Eugene, OR) and for anti-20Sα3 it was 546 goat antimouse IgG (1:200; Molecular Probes). We used argon 488-nm and helium/neon 543-nm lasers to elicit immunofluorescent staining with a Zeiss LSM 510 laser scanning confocal microscope (Zeiss; Jena, Germany).
For immunogold electron microscopy, human temporal cortex and rat hippocampus samples were fixed, embedded, and S4 immunogold labeling was performed with a three-step biotin- antibiotin-gold conjugate method (anti-S4: 1:50, 1:100; in human and rat samples, respectively). Surface density of gold particles was estimated with morphometry (Griffiths and Hoppeler 1986).
The epitope retrieval methods applied did not alter the distribution pattern of 20S either in human or in rat brain sections. Epitope retrieval with Tris-EDTA enhanced the immunostaining intensity of ubiquitin and regulatory ATPases without altering their subcellular localization, both in human and rat brain samples (Figures 1A–1L).
In human brains, ubiquitin and regulatory ATPases S4 and S7 showed mainly nuclear localization, but faint cytoplasmic immunostaining also occurred (Figures 1A and 1B, 1E and 1F, and 1I and 1J). Nucleoli were negative. S6a, S6b, S8, and S10b regulatory subunits showed mainly cytoplasmic and neuritic localization in neurons (not shown). These subcellular patterns were rather uniform in all examined brain areas in all kinds of neuron populations. In rat brains, the prominent nuclear immunolabeling of ubiquitin and S4 and S7 ATPase subunits, and mainly cytoplasmic immunostaining of S6a, S6b, and S10b subunits, confirmed the results on the human brain material (Figures 1A and 1B vs Figures 1C and 1D, Figures 1E and 1F vs Figures 1G and 1H, Figures 1I and 1J vs Figures 1K and 1L, ubiquitin, S4, and S7, respectively). However, in rat brains, S8 also showed prominent nuclear immunostaining in contrast to its predominantly cytoplasmic localization in human brains (not shown). Ultrastructural morphometry of S4 distribution confirmed its significant nuclear localization in human temporal cortical and rat hippocampal samples (Figures 2A and 2B).
In rat brain, a subset of nuclear bodies (2–5/nucleus) was strongly immunoreactive for all five 20S PS antibodies (Figures 1M and 1N, anti-20Sα/β, anti-20Sα3, respectively), for anti-19S RC S6a (Figure 1O), and for anti-ubiquitin (Figures 1P and 1Q). However, no immunoreaction was detected with anti-calmodulin (not shown), an antibody against an antigen not directly related to the UPS. In addition, confocal double labeling revealed the colocalization of 20S PS and ubiquitin in these structures (Figures 1R–1T). These bodies occurred both in neurons and glial cells in all examined brain areas and were specifically enriched in the granular cells of the dentate gyrus. In human brains, UPS-immunopositive nuclear bodies could not be detected.
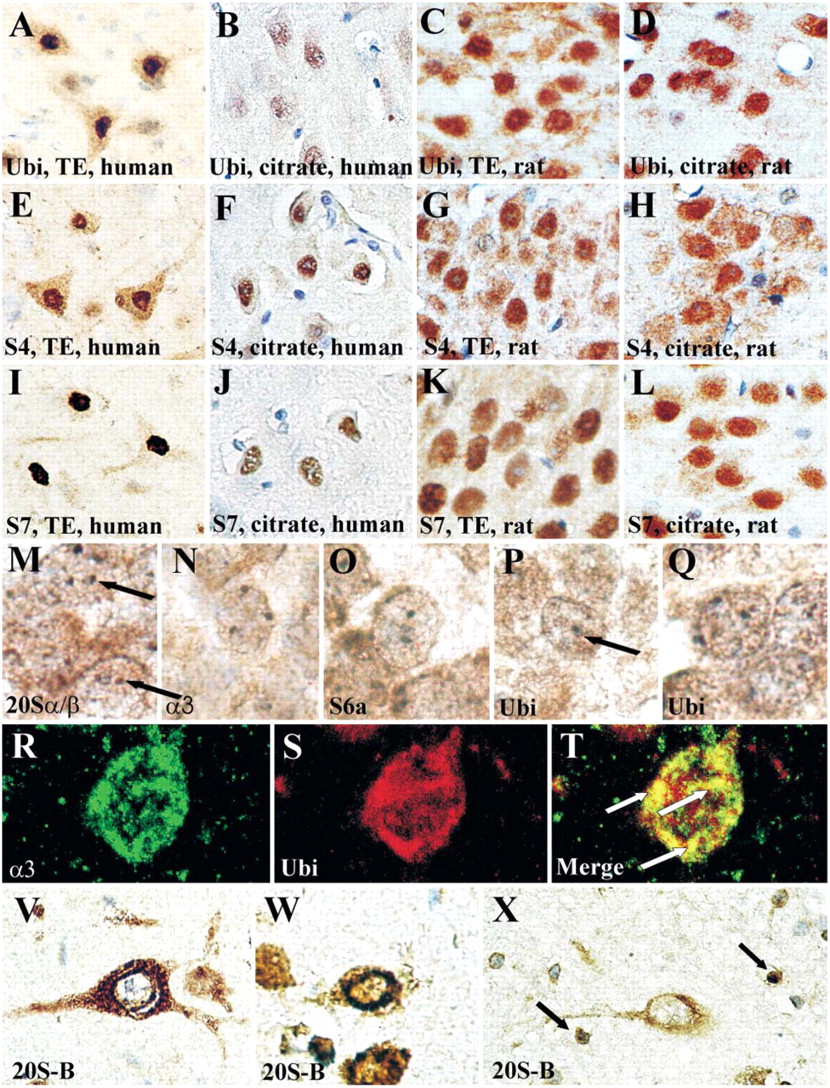
Immunohistochemical localization of the components of ubiquitin-proteasome system (UPS). Immunostaining for ubiquitin (
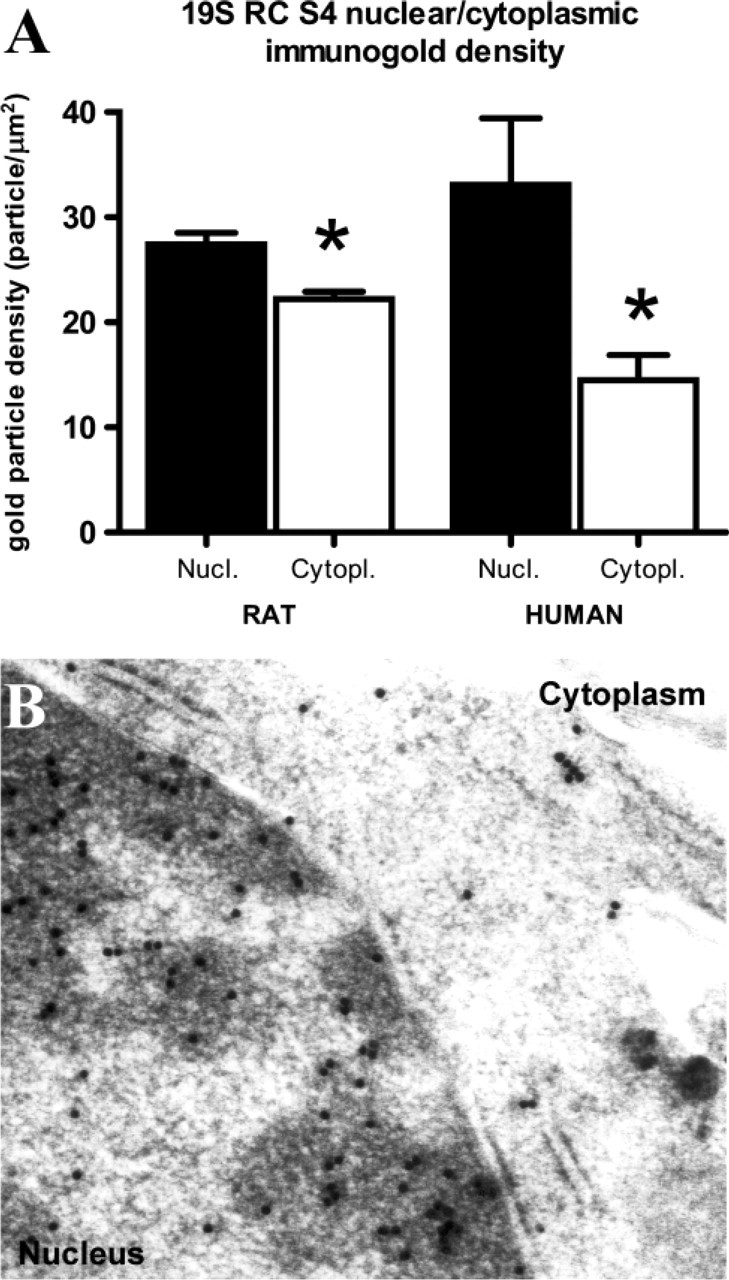
Quantitative evaluation of gold particles corresponding to S4 RC ATPase. Four tissue blocks from ten human cortical samples and four tissue blocks from eight rat hippocampal samples were randomly selected for sectioning and immunogold staining. Surface density of gold particles was estimated with morphometry on electron micrographs representing at least 500 μm2 neuronal area of human and rat brain, respectively. (
In case of 20S PS, both the pericarya and neurites were immunopositive and, frequently, intense perinuclear rings were seen (Figures 1V and 1W). In addition, many glial nuclei were also immunoreactive (Figure 1X, arrows). This was similar in human and rat brains and in cases of all 20S antibodies used (not shown).
Our observation that 20S PS immunoreactivity is both nuclear and cytoplasmic is consistent with former studies on 20S PS cellular localization (Mengual et al. 1996). The intense perinuclear staining may indicate the increased number of 20S at the nuclear envelope, which has been described before in yeast cells (Gordon 2002). The frequent nuclear immunolocalization of 20S PS in glial cells may reflect the mitotic capacity of these cells in contrast to the postmitotic neurons and may indicate the critical role of UPS in the regulation of cell cycle progression (Palmer et al. 1994).
In a recent paper, Lafarga et al. (2002) described a subtype of nuclear bodies, termed clastosomes, enriched in ubiquitin, 20S PS, 19S RC, and some proteasomal substrates in several cell types in vitro and in hypothalamic neurosecretory cells of rat brain in vivo. Clastosomes have been found to be dynamic structures as hypertonic NaCl injection strongly increased their numbers in the rat brain. The UPS-immunopositive nuclear bodies described here are strongly reminiscent of clastosomes. However, our results suggest that these structures are widespread in glial cells and in many types of neurons in rat brain, even in animals without any pretreatment. The lack of UPS-immunopositive nuclear bodies in human brain may reflect the sensitivity of these structures to postmortem effects and/or to the different fixation protocols applied.
Genetic studies in yeast have demonstrated that the six regulatory ATPases are functionally not redundant (Glickman and Ciechanover 2002). However, in a study on fractionated rat liver, significant differences were observed in the amount of different ATPase and non-ATPase 19S subunits between the nuclear and cytosolic compartment (Brooks et al. 2000). Until now, the only known comparative study of the distribution of 19S RC ATPases was performed on the degenerating larval intersegmental muscle of a moth, Manduca sexta (Low et al. 2000). The authors suggest that 26S PS can vary its precise subunit composition. One possible explanation of the different immunolocalization patterns of regulatory ATPases described in our work is that the 19S RC can exist in distinct but relatively uniform ATPase composition forms in the nuclear and the cytoplasmic compartment. On the other hand, general nuclear localization of S4, S7, and S8 is consistent with former findings that some regulatory ATPases can also function (in association with 20S PS or independently) as regulators of transcription and DNA repair (Russell et al. 1999; Gonzalez et al. 2002; Sun et al. 2002; Muratani and Tansey 2003).
Available studies on ubiquitin localization in clinical material focus mainly on pathological inclusions (Lowe et al. 1993), whereas cellular distribution has been studied less frequently. Primary nuclear immunoreactivity of ubiquitin described here may underline the prominent role of ubiquitin in stabilization of histones and regulation of numerous transcription processes (Muratani and Tansey 2003).
In conclusion, prominent nuclear location of ubiquitin, 19S RC ATPases S4, S7, and S8 provides morphological data supporting the additional function of these proteins in transcription regulation or nucleotide excision repair. A subset of nuclear bodies immunopositive for UPS components is widespread in rat brain glial cells and in many types of neurons, even in animals without any pretreatment. Furthermore, the pretreatment methods defined in our work give rationale for comparative studies on pathological human and rat samples to evaluate the exact role of the UPS during disease processes.
Footnotes
Acknowledgements
This work was supported by the Fifth and Sixth Framework Programmes of the European Community (QLG3-CT-2002–00809 and LSHM-CT-2004–503474). G.G.K received a Bolyai Fellowship.
We acknowledge Professor R. John Mayer for providing S6a, S6b, S7, and S10b antibodies and Dr. Carlos Gorbea for providing S4, 20S-A, and 20S-B antibodies. We are grateful for the helpful contribution of Rómeó Andó and for the excellent technical assistance of Ágnes Druskó, Ágnes Keserű, Gerda Ricken, Mariann Saródy, and Katalin Komjáti. Laser scanning microscope used in the Institute of Neurology, Medical University of Vienna is through the courtesy of Professor Herbert Budka.