
Abstract
Wild-type and mutated urate oxidase (UO) proteins were overexpressed in Cos-1 and HEK293 cells and were analyzed by Western blotting and several morphological methods. By immunoelectron microscopy, wild-type UO formed large aggregates in the cytoplasm and nucleoplasm and exhibited a crystalloid structure. Mutated UO (UOdC), from which 28 amino acids, including peroxisomal targeting signal at the C-terminus, were deleted, formed dispersed aggregates in the cytoplasm and nucleus. Chimeric UO (MUOdC), which was made by addition of the mitochondrial targeting signal of serine:pyruvate/glyoxylate aminotransferase to the N-terminus of UOdC, attached to ER to form a complicated MUOdC-ER complex. These three structures were immunostained for ubiquitin- and p32-subunits of proteasomes. Western blotting showed strong signal for UO and UOdC but very weak signal for MUOdC. The results suggest that overexpressed UO and UOdC accumulate in the cells because their synthesis rate is higher than the degradation rate, whereas MUOdC forming a complex with ER is degraded very rapidly. The ubiquitin-proteasome pathway may be involved in the degradation of these proteins.
I
We have studied the removal of excess rat liver peroxisomes caused to proliferate by treatment with di-2(ethylhexyl)phthalate and showed that they are digested mostly by autophagy (Yokota et al. 1993, 1995). By enzyme cytochemistry and immunocytochemistry, we have demonstrated that the isolation membrane that specifically surrounds the proliferated peroxisomes is derived from the ER. During the recovery from peroxisome proliferation to the normal state, peroxisomal cores are often observed to be present free in the cytosol. This phenomenon has been noted by Reddy and collaborators (1974). Although autophagy was presumed to play a role in the degradation of these extruded cores, it is still unclear how and where this degradation takes place. It is well known that the core consists of uricase, one of the peroxisomal enzymes (de Duve and Baudhuin 1966). Uricase is not soluble at neutral pH and forms crystalloid (Hruban and Swift 1964), which can easily be observed by transmission electron microscopy without any specific procedures. These characteristics make it convenient to analyze the destiny of uricase after overexpression by introduction of wild-type and mutant uricase genes. Introduction of the uricase gene into Spodoptera frugiperda cells has been done by Reddy and co-workers (1974), who demonstrated that the expressed uricase has normal enzymatic activity (Alvares et al. 1992). To clarify the cellular degradation of crystallized proteins, we have chosen uricase.
We prepared the following DNA probes as described previously (Niwa et al. 1991): (a) DNA encoding wild-type rat uricase (pCAUO); (b) DNA encoding mutant uricase, which lacked 28 amino acids containing the peroxisome targeting signal at the C-terminus (pCAUOdC); and (c) DNA encoding chimeric uricase, which was created by addition of the mitochondrial targeting signal of serine:pyruvate/glyoxylate aminotransferase to the N-terminus of UOdC (pCAMUOdC) (Oda et al. 1987).
Each DNA was introduced into Cos1 or HEK 293 cells by the calcium phosphate method. These cells originally had no uricase in their peroxisomes. Two days after the introduction, cells were processed for immunofluorescence microscopy, conventional electron microscopy, and immunoelectron microscopy.
Methods, Results, and Discussion
Western Blot Analysis of Products of Wild-type and Mutant Uricase Genes in Cos1 and HEK 293 Cells
Cells were washed with Hanks′ solution, collected by centrifugation, and suspended in a small amount of 0.1 M HEPES buffer, pH 7.2, containing 0.5 mM phenylmethylsulfonyl fluoride, 2 μM pepstatin, 2 μM anti-pain, and 2 μM leupeptin, and were frozen at −80C. Cell suspensions were mixed with the same volume of SDS-PAGE sample buffer and homogenized. About 40 μg of protein from each sample was loaded on to a 10% polyacrylamide gel. After electrophoresis, proteins were electrophoretically transferred to nitrocellulose membrane. Then the membrane was incubated overnight with a rabbit anti-rat uricase antibody, followed by a HRP-labeled goat anti-rabbit IgG antibody. HRP activity was visualized by the DAB reaction. The results are shown in Figure 1. In samples from Cos1 and HEK 293 cells, signals for UO and UOdC were very strong and were observed at the expected migration positions, whereas signal for MUOdC was very weak. No signals were observed in cells transfected with vector only. In samples from HEK 293 cells, in addition to the primary signals several weak signals with lower and higher molecular weights were present. The results strongly suggest that UO and UOdC are relatively slowly degraded, whereas MUOdC is degraded very rapidly. Signals with higher molecular weights might be associated with ubiquitinated wild-type and mutated uricase molecules and those with lower molecular weight might be associated with partially degraded ones.
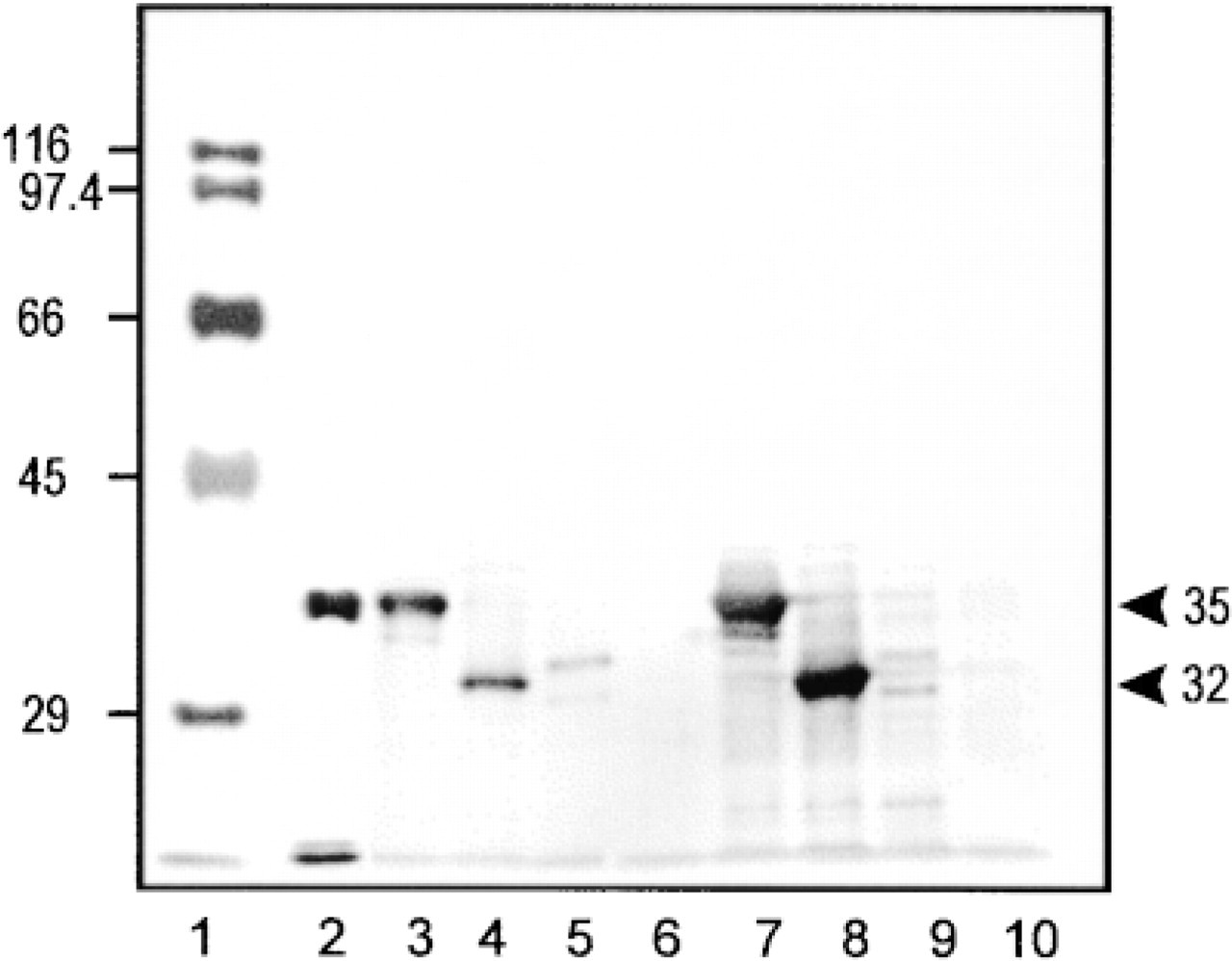
Western blot analysis of homogenates from Cos1 and HEK 293 cells overexpressing wild-type and mutant uricase. Lane 1, molecular markers whose molecular weight is shown at left. Lane 2, purified rat liver uricase. Lane 3, Cos1 cells transfected with pCAUO (encoding wild-type uricase). Lane 4, Cos1 cells transfected with pCAUOdC (encoding uricase without peroxisomal targeting signal). Lane 5, Cos1 cells expressing pCAMUOdC (encoding chimeric uricase with mitochondrial targeting signal). Lane 6, Cos1 cells transfected with vector only. Lane 7, HEK 293 cells transfected with pCAUO. Lane 8, HEK 293 cells transfected with pCAUOdC. Lane 9, HEK 293 cells transfected with pCAMUOdC. Lane 10, HEK 293 cells transfected with vector only. Numbers at right show molecular weights of wild-type uricase (35 kD) and mutant uricase (32 kD).
Immunofluorescent Localization of Uricase Antigen in Cos1 and HEK 293 Cells Expressing Wild-type and Mutated Genes
Cos1 and HEK 293 cells were negative for uricase when they were not transfected with the uricase gene, although these cells do possess peroxisomes. By immunofluorescence microscopy using the anti-rat uricase antibody, two types of staining were observed, one in subcellular compartments in which gene products were expected to be transported or localized as described in a previous study (Miura et al. 1994) and another that was present in large aggregates which were not related to the targeting signal of the products. By double labeling, large aggregates of UO were not stained for catalase, a marker for peroxisomes, suggesting that they were not related to the peroxisomes. The results were quite similar to those of Alvares et al. (1992). UOdC also precipitated to form large aggregates in the cytoplasm when it was overexpressed. When UOdC was not overexpressed, weak and diffuse staining was observed only in the cytoplasm. MUOdC was localized in mitochondria as confirmed by double labeling with anti-mitochondrial outer membrane protein. When overexpressed, it also appeared in large aggregates which had no signal for mitochondria. These staining patterns were observed in Cos1 and HEK 293 cells in common. In the case of uricase, therefore, overexpressed products form the large aggregates, being independent of its targeting signals.
Immunoelectron Microscopic Localization of Uricase Antigen in Cos1 and HEK 293 Cells Expressing Wild-type and Mutant Uricase
The same staining pattern and morphological characteristics of structures formed by UO and UOdC were observed in both Cos1 and HEK 293 cells. In cells overexpressing wild-type UO, gold particles revealing uricase antigen were present in large electron-dense aggregates and small granules (Figure 2a). The large aggregates frequently appeared in the nucleus. A few gold particles were observed in the cytoplasmic matrix (Figure 2a). UOdC (uricase without translocation signal to the peroxisome) was localized in dispersed electron-dense aggregates in the cytoplasm and sometimes in the nucleus, and these appeared quite different from UO (Figure 2b). Gold label was bound to these aggregates and few gold particles were present in cytoplasmic matrix. The results suggest that UOdC also precipitates to form cytoplasmic aggregates immediately after synthesis. In cells expressing MUOdC (chimeric uricase to which was added a mitochondrial instead of peroxisomal targeting signal) uricase antigen was found in the electron-dense matrix of mitochondria and in cyto-membrane complexes expanding in the cytoplasm (Figure 2 c). In this case, there were also no gold particles free in the cytoplasm. All gold particles were bound to the cyto-membrane complexes. The results strongly suggest that most MUOdC is attached to membrane-bound structures and a small amount is imported into the mitochondria. The structures with which wildtype and mutant uricase molecules were bound are analyzed by conventional electron microcopy.
Conventional Electron Microscopic Observation
By conventional electron microscopy, electron-dense crystalloid of UO was seen in the peroxisome matrix and in large aggregates (Figure 3a). The latter had no limiting membrane and was localized to the cytoplasm and nucleus. The results were quite similar to those from other laboratories (Alvares et al. 1992; Yeldandi et al. 1995). Aggregates of UOdC showed an electrondense meshwork of thick threads, which had no limiting membrane (Figure 3b). Therefore, when wild-type uricase and uricase with a deleted peroxisomal targeting signal are overexpressed in the cells, they precipitate immediately in the cytoplasm and the resulting precipitates without any limiting membrane are present free in the cytoplasm. Frequently, we could observe mitotic cells in which the nuclear envelope had broken down and chromosomes were exposed to the cytosol. In such mitotic cells, some aggregates of UO and UOdC attached to chromosomes (Figure 3b). These chromosome-bound aggregates might remain associated with decondensing chromosomes in the nucleus after reassembly of a new nuclear envelope. These observations could explain why uricase without any nuclear targeting signal is present in the nuclei of the cells used in our expression system. The morphology of MUOdC was quite different from that of UO and UOdC (Figure 3c). Mitochondria exhibited normal structures when MUOdC was expressed at a low level. However, when it was overexpressed, the mitochondria contained electron-dense material in their matrix, greatly increased in size, and had totally irregular shapes, suggesting that a large amount of uricase imported into mitochondria might markedly change their shape and also disturb their normal functions.
Another remarkable morphological feature in the pCAMUOdC-transfected cells was formation of cyto-membrane complexes, which consisted of ER, including the nuclear envelope, in which tubules and rings (most of them appearing to be cross-sectioned tubules) were enclosed (Figure 3c). These tubules and rings existed free neither in the cytoplasmic matrix nor in the Golgi apparatus. They were present only in the lumen of the ER or just entering into the ER with forming concavities. These results suggest that MUOdC is isolated by the ER membranes immediately after synthesis. This isolation functions to prevent strange protein from being transported into mitochondria in large quantities.
Structure, Distribution, and Degradation of UO and UOdC
By immunoblot analysis, signals for both UO and UOdC were strong, but that for MUOdC was very weak (Figure 1). This suggests that UO and UOdC are degraded slowly but that MUOdC is digested very rapidly. UO and UOdC, at least, are still present after one cell division cycle because they are imported into the nucleus, together with chromosomes, as described above. To determine the involvement of the ubiquitin system in degradation of wild-type and mutated UO, we studied the immunocytochemical localization of ubiquitin in the transfected cells. By immunofluorescent double staining, strong fluorescence for ubiquitin was observed in the aggregates of UO and UOdC that were also stained for uricase. By immunoelectron microscopy, the aggregates of UO and UOdC were heavily labeled (Figures 4a and 4b). The MUOdC-ER complex was also positive for ubiquitin (Figure 4c). These results show that some of these products were ubiquitinated to send them to the degradation system of proteasomes. The aggregates produced by these three gene products were stained for 32-kD-subunits of the 26 S proteasome (Figures 5a-5c), suggesting that the 26 S proteasome is involved in the degradation of these three products. Few autophagic profiles containing aggregates of UO, UOdC, and MUOdC were observed in both Cos1 cells and HEK 293 cells. Therefore, the ubiquitin-proteasome degradation system could be a candidate pathway for degradation of these overexpressed proteins. The slow degradation rate of UO and UOdC appears to be due to the fact that uricase molecules precipitate at neutral pH to form a crystalloid structure for UO and aggregates for UOdC. Even if proteasomes can function, they probably cannot efficiently break down these precipitated forms of uricase.
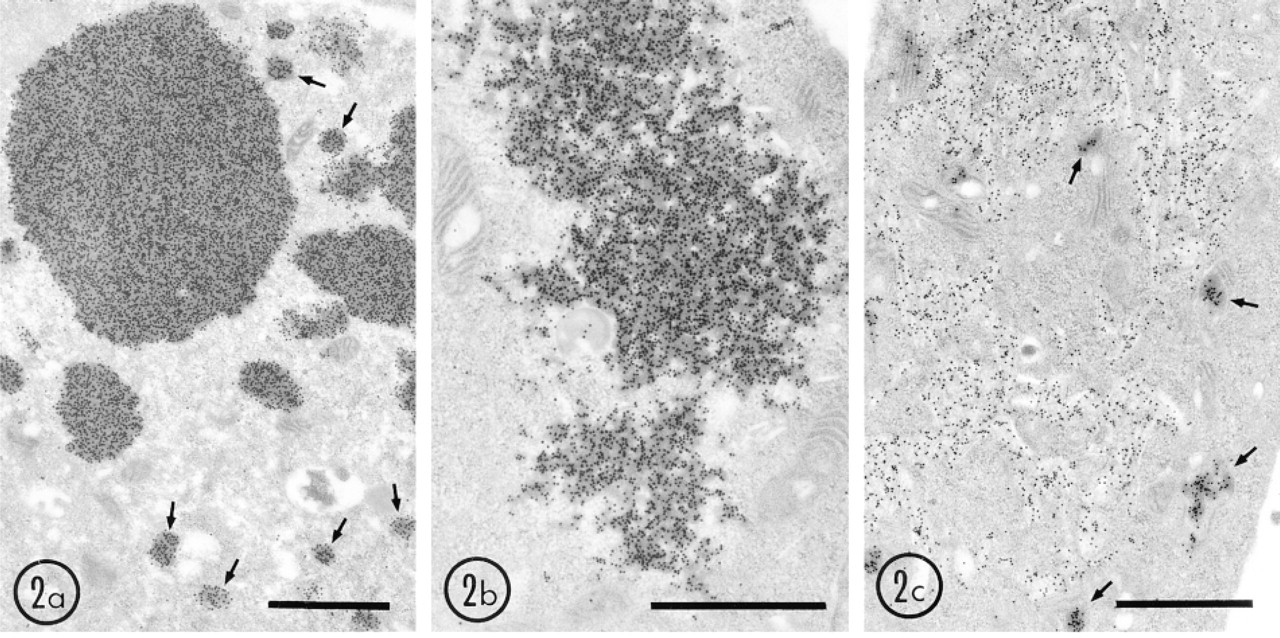
Immunogold localization of uricase in HEK 293 cells overexpressing wild-type and mutant uricase genes. Cells were fixed with 4% paraformaldehyde and 0.25% glutaraldehyde in 0.15 M cacodylate buffer, pH 7.4, for 30 min and embedded in LR White at −20C. Thin sections were immunostained with rabbit anti-rat uricase antibody and protein A-gold probe (15 nm). (
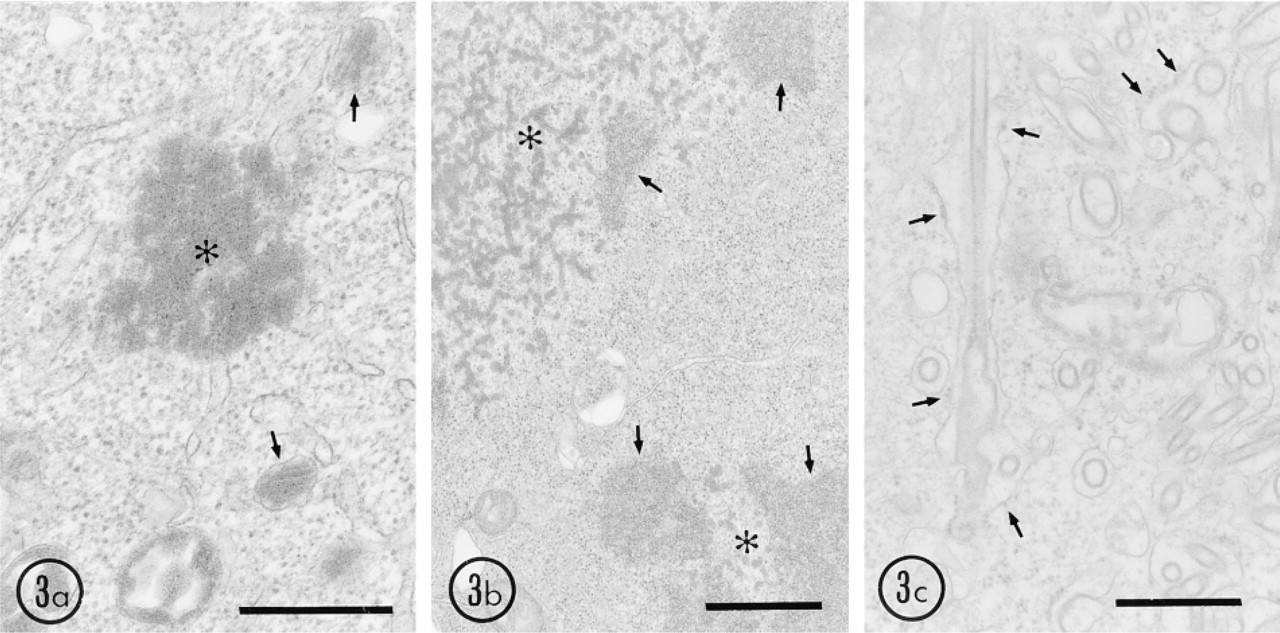
Conventional electron microscopy of HEK 293 cells overexpressing wild-type and mutant uricase genes. Cells were fixed in 2% glutaraldehyde in 0.1 M cacodylate buffer, pH 7.4, for 30 min and embedded in Epon 812. (
On the other hand, it cannot be ruled out that autophagy also functions in this system, although few profiles suggesting autophagy were observed in this study. In a previous study (Yokota 1992), we have shown that only a few autophagic vacuoles are present in hepatocytes of rats recovering from peroxisomal proliferation to the normal state. However, when such animals are treated with a proteinase inhibitor, leupeptin, marked accumulation of autophagosomes containing peroxisomes is observed. Therefore, it is necessary to test whether leupeptin treatment induces the accumulation of autophagosomes containing uricase aggregates in the present cultured cells. If autophagy acts to remove the products of UO and UOdC, we might see accumulation of autophagosomes containing the aggregates of these proteins.
In this study, aggregates of UO and UOdC were frequently present in the nucleus. It is unlikely that the autophagic system is working to remove them because there appear to be no sources for the isolation membrane enclosing the targets. An alternative degradation system might operate to remove these aggregates. After immunocytochemical staining for ubiquitin, these nuclear aggregates were stained more strongly than the cytoplasmic aggregates (Figure 4a). This indicates that UO and UOdC confined in the nucleus are much more ubiquitinated. Furthermore, they were also stained for 32-kD-subunits of the 26S proteasomes. These results suggest that the ubiquitin-proteasome system might act in degradation of UO and UOdC in the nucleus.
However, UO and UOdC appear to be degraded very slowly compared with MUOdC. This slow degradation might be due to the fact that the products form aggregates immediately after synthesis. It is likely that although some of the products are ubiquitinated, proteasomes cannot take them up into the degradation site.
Structure, Distribution, and Degradation of MUODC
By immunoblot analysis, signal for MUOdC was very weak, although strong signals for UO and UOdC were preserved (Figure 1). This suggests that MUOdC is degraded rapidly in a different manner from that of UO and UOdC. Furthermore, unlike UO and UOdC products, when the SDS-PAGE samples were freeze-thawed several times, the MUOdC signal gradually disappeared, suggesting that chimeric MUOdC, having a false signal to mitochondria, is degraded rapidly after synthesis and that the degradation occurs in the SDS-PAGE sample buffer. This result appears to be inconsistent with that of immunocytochemical staining for MUOdC, the staining intensity of which is almost identical to that of UO and UOdC. MUOdC is probably degraded during homogenization and during sampling for SDS-PAGE. Although we used proteasome inhibitors, PSI and MG-115, as well as several other protease inhibitors by homogenization of cells, degradation could not be inhibited. These results suggest that MUOdC is not broken down by proteasomes but by some other unknown degradation system. However, as shown in Figures 4c and 5c, membrane structures containing MUOdC are stained by the anti-ubiquitin antibody and by the antibody against the 32-kD-subunit of proteasomes. These results do not rule out the possibility that the ubiquitin-proteasome system might participate in degradation of MUOdC.
In cells expressing MUOdC, unique structures were observed that were never present in cells expressing UO or UOdC. MUOdC was always associated with complicated, interdigitated cyto-membrane structures or with tubules and rings locating in the dilated lumen of the ER (Figure 2c). Some of these structures were also present in the nuclear envelope but were never present in Golgi saccules and vesicles. Conventional electron microscopy confirmed that the tubules and rings are always present in the ER lumen but never directly in the cytosol (Figure 3c). Furthermore, the wall of these tubules and rings has a thick trilaminar structure that appears to be formed by close adhesion of two ER membranes with MUOdC molecules between them. These results suggest that a small amount of MUOdC is transported into mitochondria but that the ER membrane isolates the majority of it immediately after synthesis. When MUOdC is overexpressed in the cells, isolation of the nascent protein by the ER prevents massive transport of MUOdC into mitochondria. Therefore, addition of a mitochondrial targeting signal to uricase causes markedly distinct behavior in transport and degradation, which is completely different from that seen in overexpressed wild-type and mutated uricase whose peroxisomal targeting signal is deleted.
It is unclear where this chimeric uricase with a mitochondrial targeting signal is degraded, although the ER membrane isolates it as described above. Unlike UO and UOdC, this chimeric uricase appears to be rapidly degraded. It is unlikely that this rapid degradation occurs in the ER because the chimeric uricase appears to be sandwiched between two ER membranes. Consequently MUOdC is located on the outside of the ER lumen. In yeast, secretory polypetides misfolded in the ER lumen are exported for degradation from the ER to the cytosol via channels formed by Sec61p (Pilon et al. 1997). In the cytosol, these proteins are ubiquitinated and degraded by proteasomes (Kopito 1997). We cannot rule out the possibility that MUOdC is also degraded by proteasomes, as discussed above.
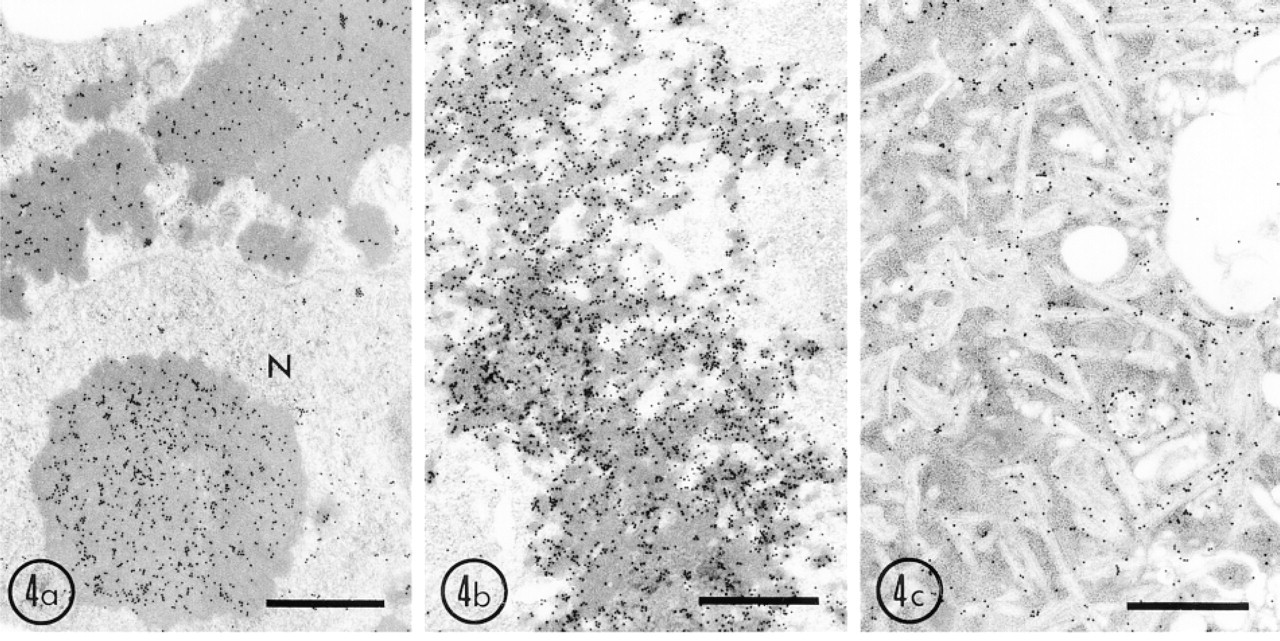
Immunogold localization of ubiquitin in HEK 293 cells overexpressing wild-type and mutant uricase genes. Cells were fixed with 4% paraformaldehyde and 0.25% glutaraldehyde in 0.15 M cacodylate buffer, pH 7.4, for 30 min and embedded in LR White at −20C. Thin sections were immunostained with rabbit anti-bovine erythrocyte ubiquitin and a protein A-gold probe (15 nm). (
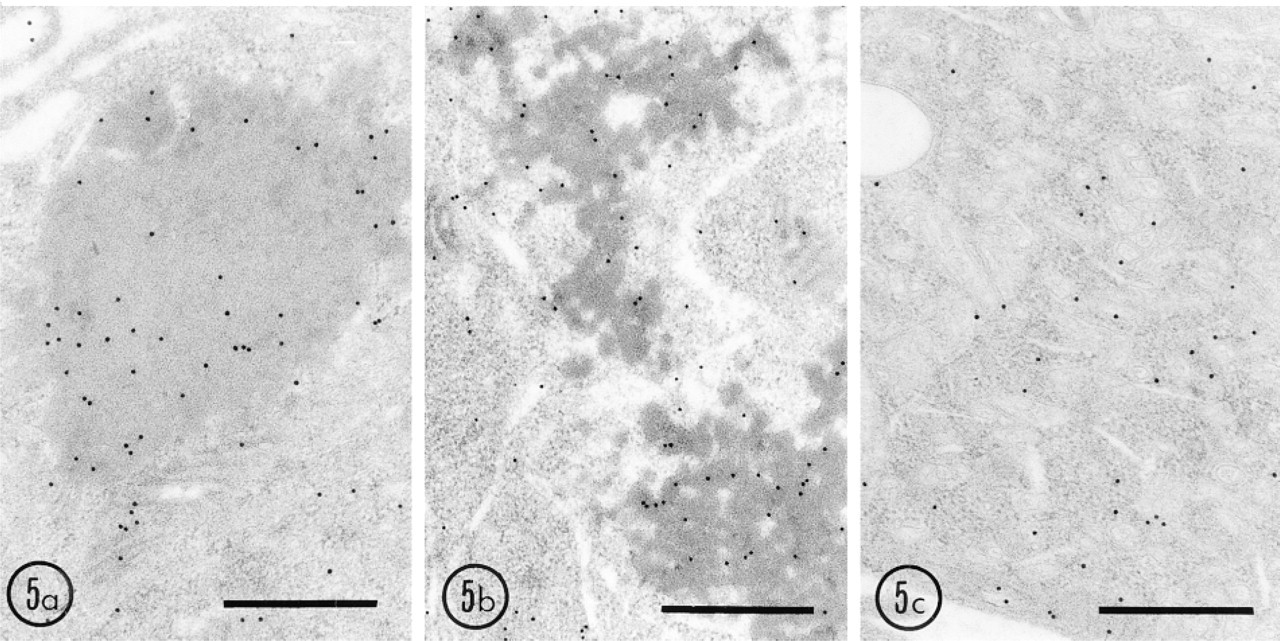
Labeling of HEK 293 cells expressing wild-type and mutant uricase genes for the 32-kD-subunit of the 26S proteasome. Thin sections of cells fixed and embedded in LR White as described above were immunostained with mouse monoclonal antibody against the 32-kD-subunit of the 26S proteasome (PROGEN; Heidelberg, Germany) and protein A-gold probe (15 nm). (
Our overexpression system presented here should be further improved because we did not use a system in which the expression machinery can be turned on and off. To study the precise degradation of the expressed proteins, the expression should be controlled in some way, e.g., by temperature, hormones, or drugs. Although our uricase expression system is not complete, it is clearly shown that products of wildtype and mutated uricase proteins might be degraded via different pathways, which can be morphologically distinguished from the lysosomal system. Furthermore, we have shown that when organelle protein is manipulated via its targeting signal, it is broken down in different manners and in different places. The present overexpression system of uricase appears to provide a good method for study of the intracellular degradation pathway of proteins.
Footnotes
Acknowledgements
Supported in part by grants-in-aid for Scientific Research No. 03833013 to SY and No. 10480165 to TO from the Ministry of Education, Science, Sport and Culture of Japan, and by a grant to TO from the Naito Foundation, Japan.
We are grateful to Dr J. Miyazaki (Osaka University Medical School) for pCAGGS vector, to Drs M. Mori (Kumamoto University School of Medicine) and A. Ichiyama (Hamamatsu University School of Medicine) for useful advice and help during this study, to Dr S. Asano (Toyama Medical & Pharmaceutical University) for HEK293 cells, and to Dr J.K. Reddy (Northwestern University Medical School) for reading this paper and correcting the English usage.