
Abstract
Mucin glycoproteins and trefoil peptides play an important role in protection and repair of the gastrointestinal epithelium. This study investigates alterations in mucin and trefoil peptide gene expression and product localization in ulcerative colitis (UC). Product localization and message expression of mucin MUC1 to 6 and trefoil peptide TFF1 to 3 genes was analyzed in rectosigmoid tissue from a cohort of patients with active UC and compared with that of normal colorectal mucosa. MUC1 expression was upregulated in severe UC at the site of rupture of crypt abscesses. Reduction in MUC2 expression occurred in UC adjacent to ulceration. No alteration in MUC3 or MUC4 gene expression was detectable in UC compared with normal colorectal mucosa. No ectopic expression of MUC5AC, MUC5B, or MUC6 was identified in UC. Ectopic TFF1 expression was identified in tissues eliciting histological features of severe disease. Decreased TFF3 localization was demonstrated in UC tissues, but no TFF2 expression was detected in any colorectal specimens. Subtle alterations in composition of the supramucosal defense barrier exist in UC and vary in relation to clinical severity of disease. There is upregulation in mucin MUC1 at crypt abscesses and neo-expression of TFF1 trefoil peptide in severe disease.
Keywords
U
Mucin glycoproteins MUC1–4 are expressed by the colorectal epithelium, and the physical properties of the mucus gel layer in UC are certainly altered compared with normal characteristics (Pullan et al. 1994). Alterations, however, in the expression of the mucin genes MUC1-4 have not been conclusively identified (Myers-cough et al. 1995; Weiss et al. 1996). Sulfation of mucins appears to be reduced in UC (Corfield et al. 1992,1996). This may have important bearing on the stability of the mucus gel layer and its ability to protect the colorectal mucosa from luminal microorganisms. The role of the transmembranal mucins (MUC1, MUC3, and MUC4) in the immunoregulatory control of large-bowel mucosal homeostasis is, at present, uncertain.
Trefoil peptides play an important role in the repair of the injured gastrointestinal mucosa (Mashimo et al. 1996). TFF3 (trefoil factor family) expression may be reduced in UC (Podolsky 1997), but there is, as yet, no evidence for ectopic expression of TFF1 or TFF2 in adult large bowel inflammation. Any role for trefoil peptides in the immune responses seen in the gastrointestinal tract has yet to be identified. Likewise, any relationship between the severity of disease in ulcerative colitis and alterations in the mucin and trefoil components of the colorectal supramucosal defense barrier/mucus gel layer has not previously been examined.
This study investigates alterations in mucin gene and trefoil peptide gene expression and product localization between normal large bowel mucosa and UC mucosa. Not only are the constitutively expressed colorectal mucins (MUC1, MUC2, MUC3, and MUC4) and trefoil peptides (TFF3) examined but so are other gastrointestinal tract mucins (MUC5AC, MUC5B, and MUC6) and trefoils (TFF1 and TFF2). Furthermore, this study examines alterations in these components of the supramucosal defense barrier in relation to clinical severity of disease. With the knowledge that mucin sulfation may play an important physical and biological role in mucin function, this study also investigates potential alterations in localization of an important sulfated mucin epitope in relation to UC severity.
Materials and Methods
A prospective study recruiting a cohort of patients with active UC was undertaken. Patients admitted to the hospital and those attending as outpatients for colonoscopy with a known or probable diagnosis of UC were considered for inclusion in the study. All patients admitted to the hospital were managed under a standardized treatment protocol. Those patients receiving parenteral, rectal, or oral steroids were excluded from the study, as were patients <16 years of age. All patients were fully informed and provided consent to participate in the study following Local Research and Ethics Committee approval. All patients meeting the inclusion criteria provided a rectal biopsy taken at endoscopy. Biopsies were taken from the rectosigmoid region at an area of maximal endoscopic disease but where intact mucosa appeared to exist. The patients recruited for the study were classified into clinical disease severity according to the Truelove and Witts (1955) criteria with hematological, biochemical, and physiological markers available from the routine investigation and management of those in-patients with UC.
A cohort of patients undergoing elective surgery for colorectal resections were recruited with full informed consent to obtain normal rectal tissue to be used as controls. No patients had known inflammatory bowel disease or were undergoing large bowel resection for either neoplastic or functional bowel disease. Biopsies of macroscopically normal rectosigmoid tissue were taken from each patient's resection specimen at a distance >5 cm from any identified mucosal pathology to exclude the possibility of a field change effect (Jass 1993; Longman et al. 2000a).
Paraffin-embedded tissues from normal bronchus (n=2), cervix (containing endo- and exo-cervix, n = 1), jejunum (n=2), ileum (n=3), esophagus (squamo-columnar junction, n = 1), stomach (fundus, n=2; body, n = 3; antrum, n=3), submandibular salivary gland (n=2), and from lobular breast cancer (n=1) were used as standard positive and negative controls for all immunohistochemical (IHC) and in situ hybridization (iSH) experiments.
Tissue Histochemistry
All tissues collected were immediately formalin fixed and subsequently paraffin embedded within 24 hr. Tissue sections were stained with hematoxylin and eosin to confirm diagnosis and to differentiate severity of inflammation according to the Bristol Index (Warren et al. 1988) (Table 1). Two independent histopathologists (NAW and BFW) performed this confirmation. Sections were stained with a number of histochemical techniques to identify patterns of mucin production.
Diastase periodic acid Schiff's (PAS) reagent and Alcian blue (AB) stain were used to identify patterns of acidic and neutral mucins. PAS identifies sialylated and sulfated carbohydrates and neutral mucins, whereas AB identifies the most sialylated mucins. PAS/AB stain provides a histochemical representation of the acidity or neutrality of mucins and a representation of sialylated and sulfated mucins within the tissue sections. Tissue sections were then stained with 1% AB in 3% acetic acid (pH 2.5) for 5 min. After washing, they were immersed in 1% periodic acid for 8 min and stained with Schiff's reagent for an additional 8 min. Sections were then counterstained with Carazzi's hematoxylin.
Mild PAS (mPAS) stain was used to identify sialylated mucins; mPAS can only oxidize sialic acids and not sulfated mucins or other carbohydrates. Sections were rehydrated and then washed in 0.1 mol/liter acetate buffer (pH 5.5) at 4C for 5 min prior to oxidation with 1 mM sodium periodate in 0.1 M acetate buffer (pH 5.5) at 4C for an additional 5 min. Sections were then incubated in 1% aqueous glycerol for 5 min before staining with Schiff's reagent at room temperature for 15 min before incubation with 0.5% potassium metabisulfate in 0.05 M HCl for 10 min. Sections were then counterstained with Carazzi's hematoxylin. mPAS stain was also used with and without saponification. The saponification process allows comparison of O-acetylated sialic acids, as mPAS or PAS will not demonstrate sialic acid groups if they contain O-acetyl groups. Sections were mildly saponified by incubation with 0.1 mol/liter potassium hydroxide for 30 min at room temperature and then stained with Schiff's reagent and hematoxylin as above.
High iron diamine (HID) and AB stains were used to identify sulfated and carboxylated mucins in addition to sialylated mucin. Tissue sections were rehydrated and then stained for 24 hr with a solution containing 0.116% ferric chloride, 0.1 M N,N-dimethyl-meta-phenylenediamine-dihydrochloride and 2 mM N,N-dimethyl-para-phenylene-diamine-dihydrochloride. Sections were then counterstained with 1% AB in 3% acetic acid (pH 2.5) for 5 min (Filipe and Lake 1983).
Mucin Glycoprotein and Trefoil Peptide IHC
IHC detection and localization of mucins MUC1, 2, 4, 5AC, 5B, and 6 and trefoil peptides TFF1, TFF2, and TFF3 were performed (Table 2 for primary and secondary antibody characteristics and their dilutions). No suitable anti-human MUC3 antibody was available at the time of this study.
Bristol Index scoring systema for histological severity of ulcerative colitis (UC) (Warren et al. 1988)
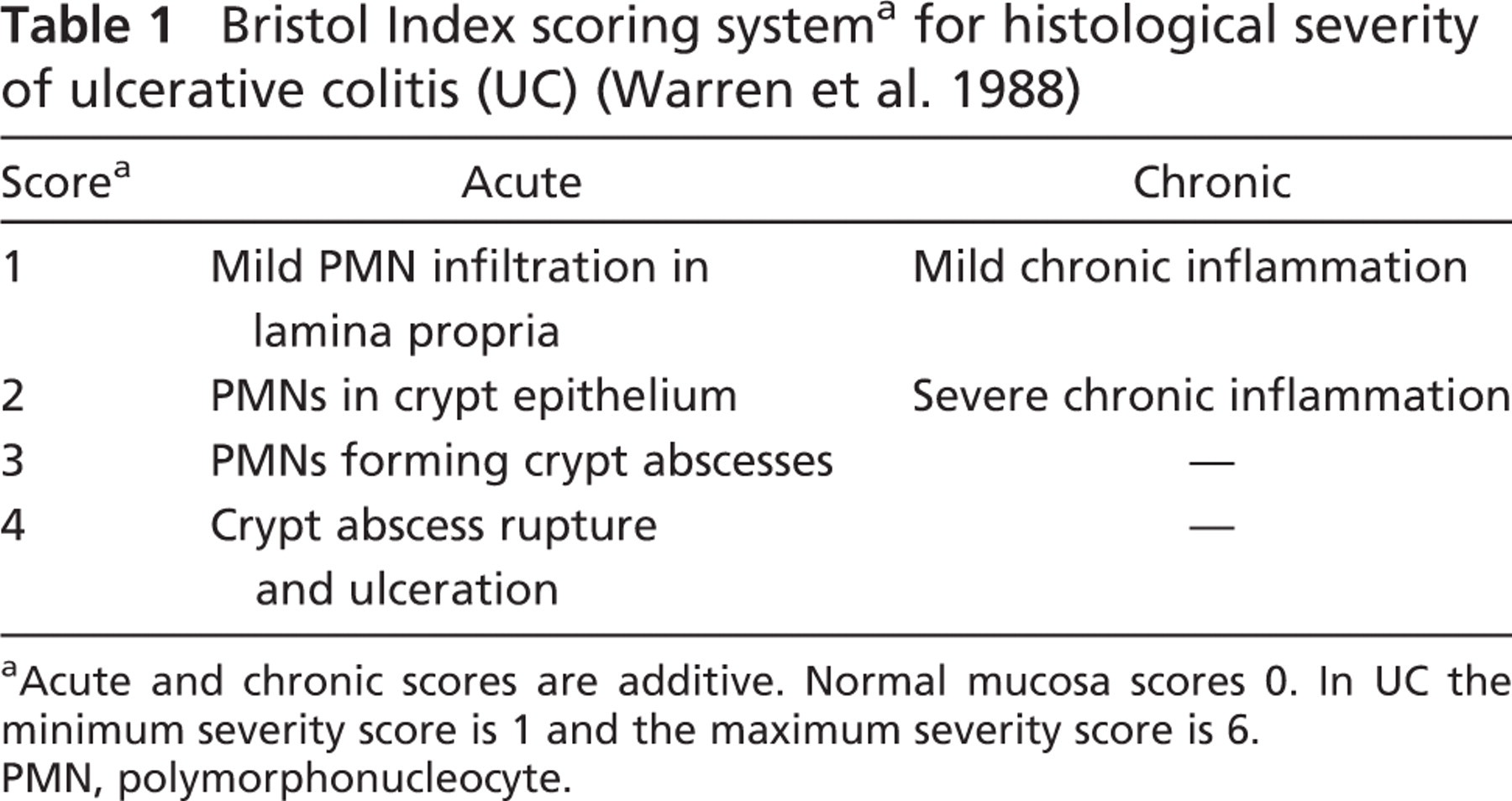
Acute and chronic scores are additive. Normal mucosa scores 0. In UC the minimum severity score is 1 and the maximum severity score is 6.
PMN, polymorphonucleocyte.
All immunohistological staining was performed on 5-μm serial sections cut onto poly-l-lysine-coated slides. Tissue sections were rehydrated before immunostaining. MUC2, MUC4, and TFF3 antigen presentation was undertaken by heating tissue sections in a domestic pressure cooker at 121C for 2 min while immersed in 10 mM citrate buffer, pH 6.0. Endogenous peroxidase activity was then blocked with 1% hydrogen peroxide in water. Tissue reduction for MUC2 immunostaining was performed with 10 mM dithiothreitol in 10 mM Tris (tris [hydroxymethyl] aminomethane) HCl, pH 8.0, for 30 min at 37C. Tissue sections for MUC1 and MUC4 staining were incubated in 4% commercial non-fat skim milk powder in PBS, 0.15 M sodium chloride, 2.5 mM sodium dihydrogen orthophosphate dihydrate, 8 mM disodium orthophosphate, pH 7.4, for 15 min. All sections were subsequently blocked with 10% normal goat serum (Dako; High Wycombe, Buckinghamshire, UK) in PBS for 20 min. Primary anti-human and secondary antibody characteristics and their working dilutions are shown in Table 2. Tissue sections were incubated overnight at 4C with the appropriate primary antibody diluted in PBS. Secondary antibodies diluted in PBS were applied for 1 hr at room temperature. Streptavidin-horse-radish peroxidase conjugate (Dako), diluted to 1:500 in PBS, was applied to sections for MUC1 and MUC4 staining.
Immunostaining for MUC5AC, MUC5B, and MUC6 was performed as for MUC2 but without a reduction stage. IHC staining for TFF1 used a mouse monoclonal IgG1 antibody directed against the C-terminal domain of human TFF1. Tissue sections were rehydrated before blocking for endogenous peroxidase with 1% hydrogen peroxide in methanol for 5 min. Sections were washed in PBS before blocking with 1:25 rabbit serum (Dako) in PBS for 15 min at room temperature. After draining off the rabbit serum, sections were incubated with the primary antibody in 1:25 fetal calf serum (Gibco BRL; Paisely, Renfrewshire, UK) in PBS for 35 min at room temperature. The primary antibody was washed off with PBS and then replaced with the secondary antibody made up in PBS for 35 min at room temperature. After further washings in PBS, sections were incubated in 1:500 streptavidin-peroxidase (Dako) in PBS for 30 min at room temperature.
Staining for TFF2 was carried out similarly. After rehydration and blocking for endogenous peroxidase, sections were incubated with the TFF2 antibody, anti-hSP, and diluted in PBS for 35 min at room temperature. Following washings in PBS, sections were incubated with the hydrogen peroxidase-conjugated secondary antibody for 35 min at room temperature.
Slides were developed with 0.6 mg/ml 3,3′-diaminobenzidine and 0.037% (v/v) H2O2 to demonstrate peroxidase activity and finally counterstained with Ehrlich's hematoxylin.
TFF3 IHC of all tissue sections was repeated but counterstained with AB to aid discrimination of goblet cell vesicles with and without TFF3 staining.
Substitution of primary antibodies with PBS was used as negative controls for all tissue sections.
Scoring of tissues for immunostaining was performed by conventional light microscopy. Maximal intensity of staining was scored on a scale of 0 (0, +, ++, and + + +) to 3. Proportion of staining for cell types and cell localization was performed for goblet cells, absorptive colonocytes, cells at the luminal surface, and deep cells in crypts or crypt remnants (0 = no cells with staining; 1(1) = 0% to 25% of cells with staining; 2(++) = 25% to 50%; 3(+++) = 50% to 75% of cells with staining; 4(++++) = 75% to 100% of cells with staining). Two independent observers (NAW and BFW) performed all IHC scoring.
Sulfomucin IHC
IHC staining of the tissue sections with the F2 antibody was undertaken (see Table 2 for primary and secondary antibody characteristics and their dilutions). This antibody is directed against part of the sulfated Lewisa (SO3-Galβ1-3GlcNAc) epitope common to colonic mucins (Veerman et al. 1997). Expression of this epitope in salivary mucins is strongly correlated with mucin sulfate content. Although this is probably only one of many sulfated mucin carbohydrate epitopes present in colonic mucins, its histochemical detection may provide insight into altered mucin sulfation in UC.
Human mucin and trefoil peptide antibodies
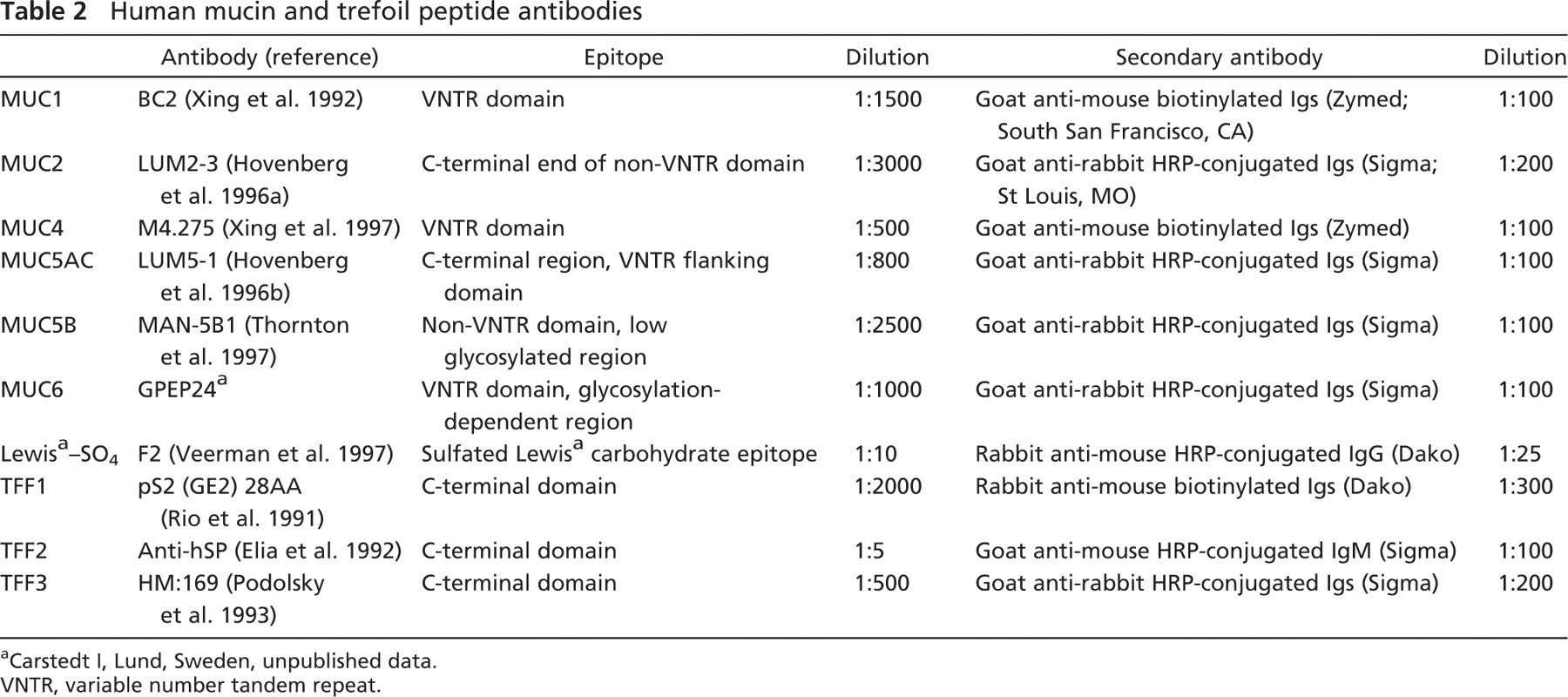
Carstedt I, Lund, Sweden, unpublished data.
VNTR, variable number tandem repeat.
Human mucin and β-actin 48-mer probe oligonucleotide sequences

Tissue sections were rehydrated prior to antigen presentation undertaken by heating tissue sections in a domestic pressure cooker at 121C for 2 min while immersed in 10 mM citrate buffer, pH 6.0. Endogenous peroxidase activity was then blocked with 1% hydrogen peroxide. Sections were incubated for 1 hr at room temperature with 1:5 dilution of the F2 antibody and 0.1% sodium azide in PBS. After washings with PBS, sections were incubated overnight at 4C with the secondary antibody made up in PBS containing 3% rabbit serum (Dako). After further washings with PBS, all IHC slides were developed and then counterstained with hematoxylin according to the methodology stated previously.
Scoring of tissues for immunostaining was performed by conventional light microscopy, as for mucin and trefoil IHC by two independent observers (NAW and BFW).
Mucin Glycoprotein ISH
Detection of mucin MUC1, MUC2, MUC3, MUC4, MUC5AC, MUC5B, and MUC6 mRNA in tissue sections by ISH was performed. Specific 48-mer oligonucleotide anti-sense probes directed against variable number tandem repeat (VNTR) domains of the mucin gene mRNAs were end-labeled with 35S-deoxy-adenosine triphosphate (see Table 3). Hybridizations with 2 × 105 cpm/slide of labeled oligonucleotide probe to the specific mRNA were performed and to β-actin mRNA as a control.
Probe labeling, ISH, and tissue section developing was performed as previously described (Longman et al. 2000b). Controls consisted of a prehybridization digestion of duplicate tissue sections with RNase A (Boehringer Mannheim; Lewes, East Sussex, UK) to remove cleaved RNA fragments. After developing, tissue sections were counter-stained with toluidine blue. Localization of mucin mRNA was performed by examining the counterstained slides under light- and darkfield microscopy, and semiquantitative assessment of mRNA was scored on a scale of 0 (0, +, ++, and +++) to 3.
Trefoil Peptide ISH
Detection of TFF1, TFF2, and TFF3 tissue mRNA was performed by ISH with 1 × 106 cpm/slide of 35S-UTP labeled anti-sense riboprobes. The probe for TFF1 was derived from a 408-base pair (bp) PVUII/BamHI fragment of the TFF1 cDNA subcloned into pSP73. The TFF2 λ200 clone 5 probe was a 300-bp BamHI/BamHI fragment of the TFF2 cDNA subcloned into PGEM1. TFF3 hybridization used a 120-bp XhoI/BamHI fragment of the TFF3 cDNA subcloned into pBluescript-II/SK-EcoRI. Transcribed and labeled sense riboprobes were used as controls. Plasmid vectors containing DNA templates were supplied by Dr. R. Poulsom (In Situ Service; Histopathology Unit, Cancer Research UK, London) (see Table 4).
Probe labeling, ISH, and tissue section developing were performed as previously described (Poulsom et al. 1997).
Semiquantitative assessment of mRNA tissue content was performed according to the mucin oligoprobe ISH methodology, with independent assessment by two histopathologists (NAW and BFW).
Data Analysis
Concordance between independent observers was >85%, and when disagreement between observers occurred the average score of histological assessment was taken and rounded up to the nearest whole number.
Trefoil peptide riboprobes, plasmid vectors, linearizing restriction enzymes, and RNA polymerases
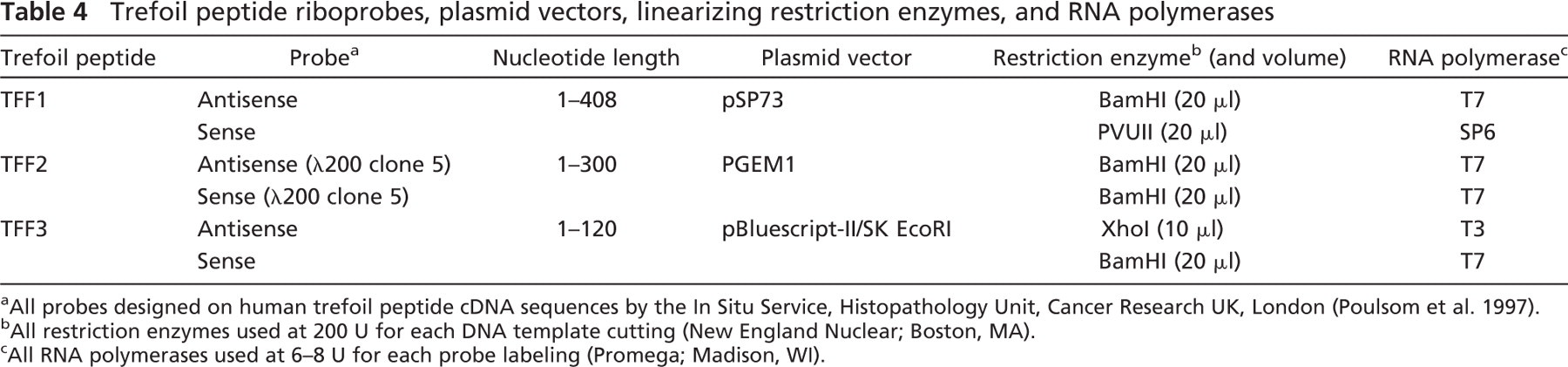
All probes designed on human trefoil peptide cDNA sequences by the In Situ Service, Histopathology Unit, Cancer Research UK, London (Poulsom et al. 1997).
All restriction enzymes used at 200 U for each DNA template cutting (New England Nuclear; Boston, MA).
All RNA polymerases used at 6–8 U for each probe labeling (Promega; Madison, WI).
Summary of details of patients with UC
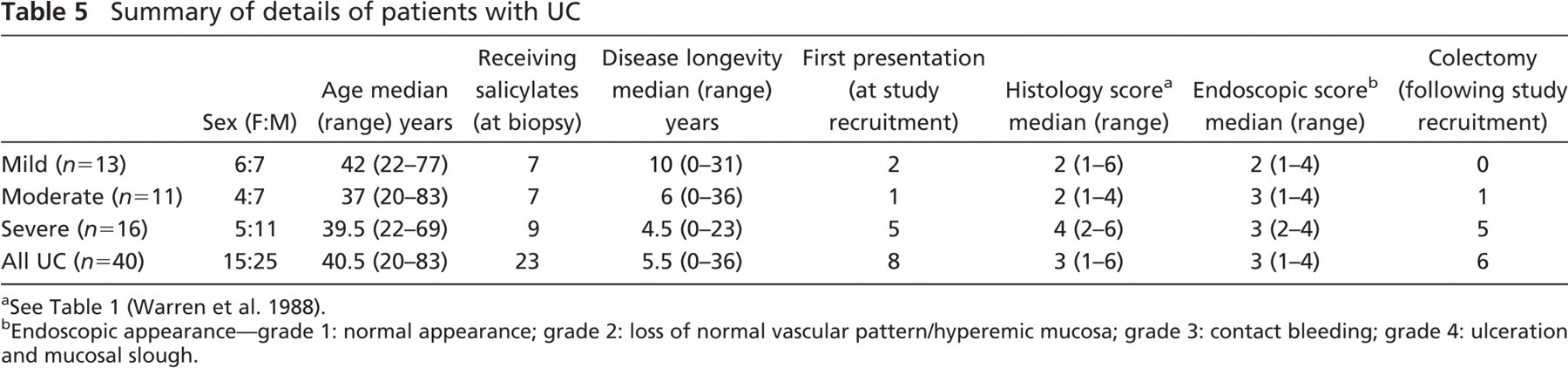
See Table 1 (Warren et al. 1988).
Endoscopic appearance-grade 1: normal appearance; grade 2: loss of normal vascular pattern/hyperemic mucosa; grade 3: contact bleeding; grade 4: ulceration and mucosal slough.
Comparison between clinical severity and histological severity of UC was determined by Kendall's rank correlation. This test was continuity corrected and corrected for ties (Stata version 6.0, 1999; Stata Corporation, College Station, TX) for Apple Macintosh.
Results
Tissues and Histochemistry
Tissues studied from 40 Caucasian patients were confirmed to exhibit histological features consistent with a diagnosis of acute active UC. Of the 40 patients with active UC, 13 were classified as clinically mild UC, 11 with moderate disease, and 16 with severe colitis (Truelove and Witts 1955). The cohort consisted of 15 female and 25 male patients with a median age of 40.5 years (range: 20–83 years) (Table 5).
Strong correlation was statistically detected between clinical severity and histological severity of UC for the patients (τ-a = 0.423; τ-b = 0.573; p<0.001).
Surgical resection biopsies from 17 patients without inflammatory bowel disease and exhibiting macroscopically normal rectosigmoid mucosa were obtained as control tissues. All control biopsies from these 17 patients were confirmed as histologically normal.
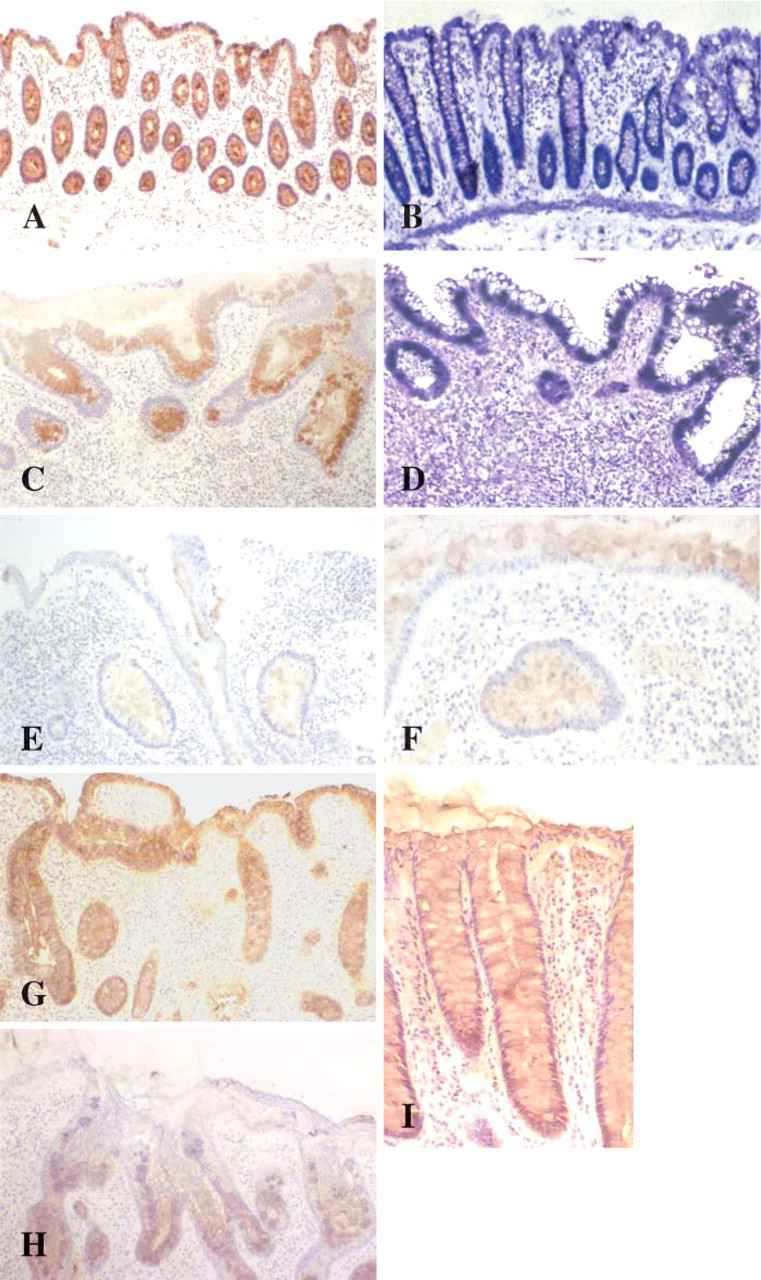
Detection of mucin charge using PAS/AB stain demonstrated a mixed pattern of neutral and more acidic sialylated mucins throughout the mucosa in all colorectal sections studied. More blue staining acidic sialylated mucins were detected toward the luminal surface in all sections, whereas more magenta-staining neutral mucins were detected in the crypts. This pattern remained in colitic tissues (Figures 1A and 1B).
With mild PAS stain, goblet cell vesicles in all colitic tissues either demonstrated no staining or stained lightly pink, indicating the presence of sialylated mucins. Following saponification, the majority of goblet cells stained more strongly pink indicated the presence of O-acetylated sialic acids. No colonic tissue failed to exhibit this histochemical staining pattern.
HID/AB stain demonstrated a dark brown stain in goblet cell vesicles deep in crypts indicating the presence of sulfomucin populations, whereas a greater presence of blue coloration was seen in vesicles toward the surface of the lumen indicating a greater presence of carboxylated sialomucins. All colitic tissues studied exhibited this pattern of staining where sufficient crypts and surface epithelium remained (Figures 1C and 1D).
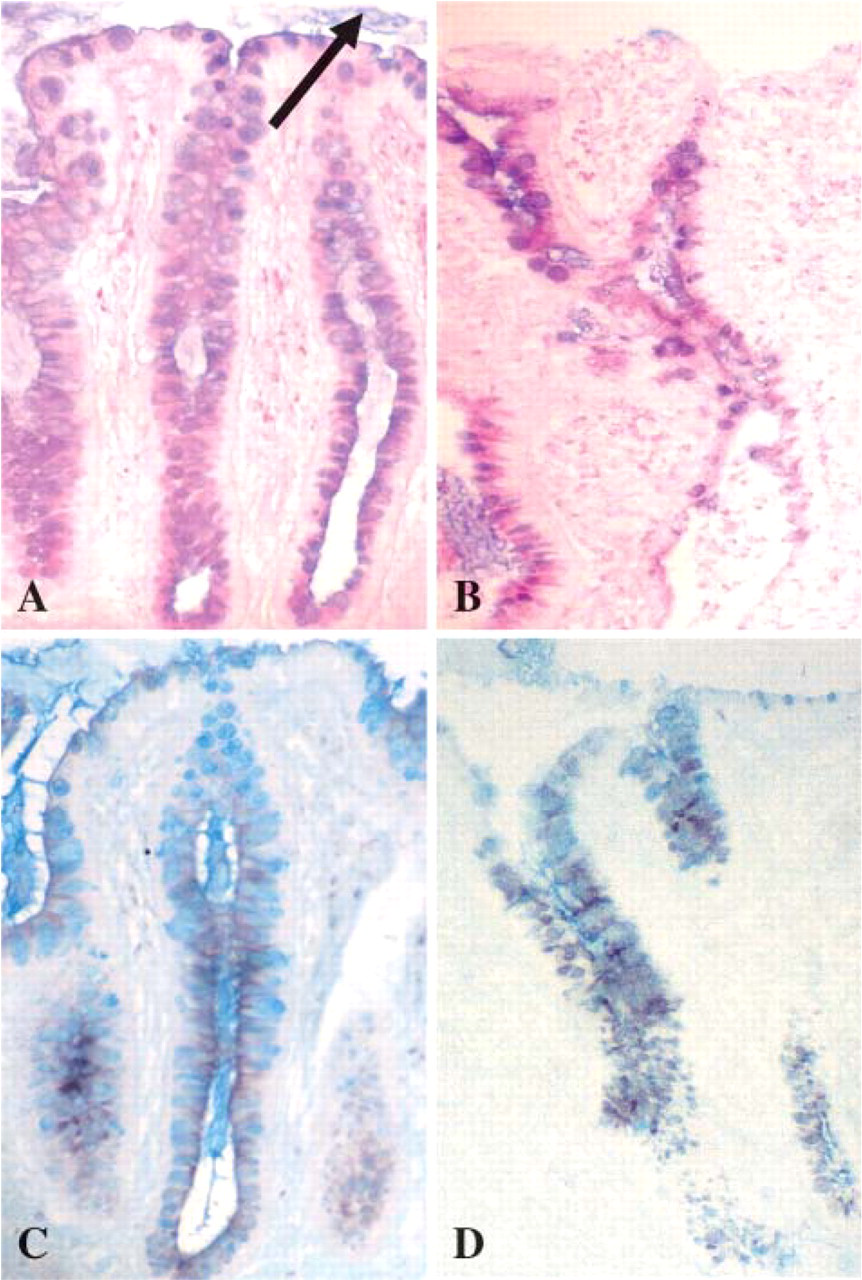
Histochemical staining of colorectal tissue: normal mucosa (
No striking differences were detected between the proctosigmoiditis samples and that of non-inflamed normal colorectal tissue. No clear differences in histochemistry were identified in relation to differences in clinical severity of UC.
Membrane-associated Mucin Glycoprotein Gene and Product Expression
In normal rectal tissues, MUC1 mRNA and product was localized to both absorptive and goblet-type colonocytes (Figures 2A and 2B). Localization of glycoprotein was to both absorptive colonocytes and goblet cells with cytoplasmic staining and membranal staining. A preponderance of MUC1 glycoprotein localization, both in number of cells and intensity, to the luminal surface was increased in UC compared with non-inflamed mucosa (Table 6). In UC this increase in immunostaining was predominantly in the cytoplasm. In severe UC the strongest expression of MUC1 mRNA (Figure 2C) was within epithelial cells at the point of potential or actual crypt abscess rupture at the crypt bases, and this was matched by the MUC1 glycoprotein localization (Figures 2E–2G). Crypts in active UC without abscess formation did not demonstrate this expression pattern (Figure 2D).
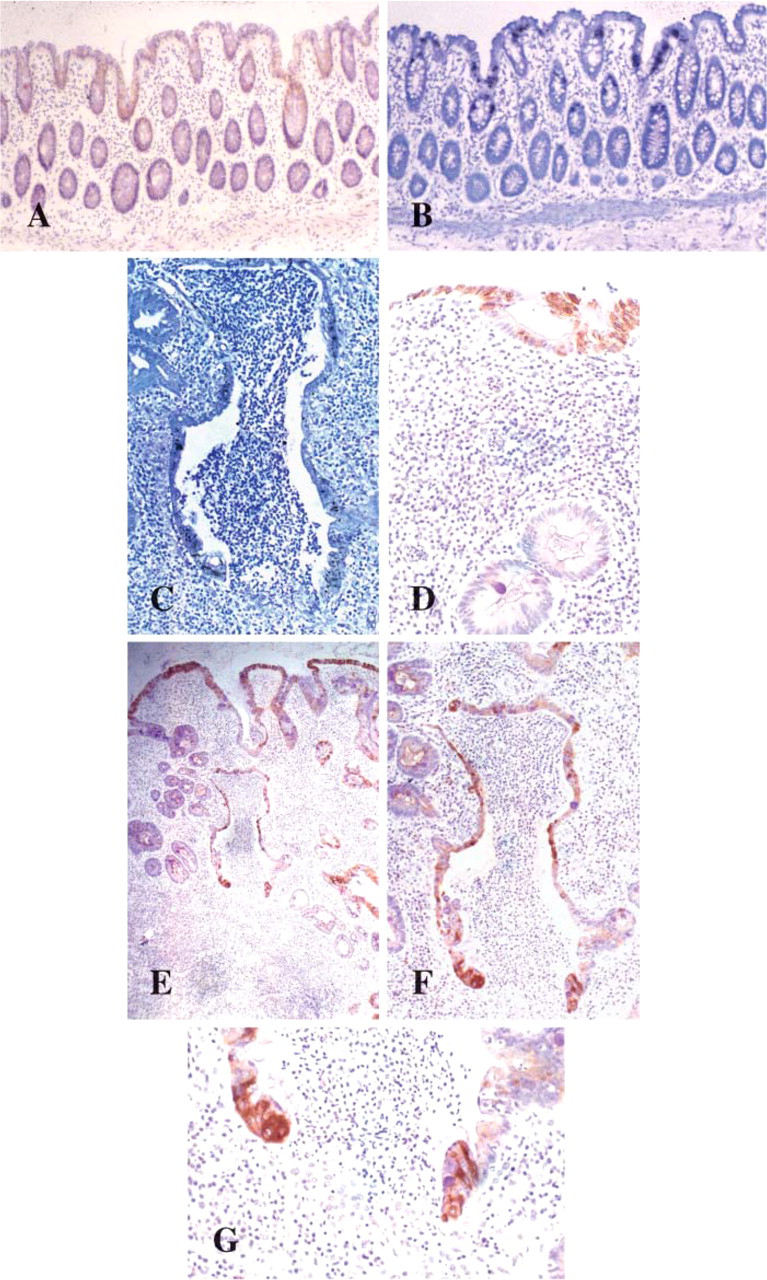
MUC1 glycoprotein (
Weak MUC3 mRNA expression was unchanged between normal and UC biopsies irrespective of disease severity (Table 6). MUC3 gene message was located at the surface epithelium and upper half of the crypts in 13/17 normal colorectal biopsies (Figure 3A) and in 26/40 colitic biopsies (Figure 3B).
The localization pattern for MUC4 glycoprotein identified strong cytoplasmic staining and strong mRNA expression throughout the colorectal epithelium in both UC and normal tissues (Figures 3C–3F, 3I). IHC staining was detected in both absorptive and goblet cell colonocytes. The MUC4 M4.275 antibody used in this study is raised against the VNTR domain of MUC4 and is thus likely to detect the unglycosylated precursor rather than the mature glycoprotein. This is reflected by the IHC localization of mucin MUC4 to the cytoplasm (Figure 3G). Only subtle differences in staining patterns occurred with a degree of heterogeneous expression from region to region along the mucosa in UC (Figure 3H). A slight preponderance for a decreased proportion of cell staining was identified deep in crypts and in the goblet cell population when contrasted with increasing disease severity (Table 6).
Secreted Mucin Glycoprotein Gene and Product Expression
Mucin MUC2 gene expression and product were solely associated with the secretory vesicles of the goblet cell population throughout inflamed and normal colorectal tissue (Figures 4A–4D). A subtle reduction in mRNA expression was notable in colitic sections, and a reduction in the numbers of goblet cells exhibiting immunostaining in the severe colitic group was apparent (Table 7). In some colitic tissues a relative absence of MUC2 staining was apparent at the mucosal surface adjacent to ulcers (Figure 4E) although this was not a global finding, whereas immunostaining was maintained in areas distant to ulceration (Figure 4F).
There was no expression of the other chromosome 11p15-secreted mucins, namely, MUC5AC, MUC5B, and MUC6, in either the colitic or normal colorectal mucosal tissues studied (Table 7).
Patterns of Sulfomucin Localization
Detection of the sulfo-Lewisa mucin carbohydrate epitope with the F2 antibody demonstrated strong cytoplasmic staining in absorptive colonocytes and cytoplasmic and vesicular staining in goblet cells throughout the mucosa in normal colorectal tissue (Figure 4G). In colitic biopsies this strong staining pattern was also evident in mild disease (Figure 4I), but loss of staining intensity and reduction in positive staining in absorptive cell populations occurred with clinically severe disease (Table 8; Figure 4H).
Summary of expression and localization of membrane-associated mucins in ulcerative proctosigmoiditis
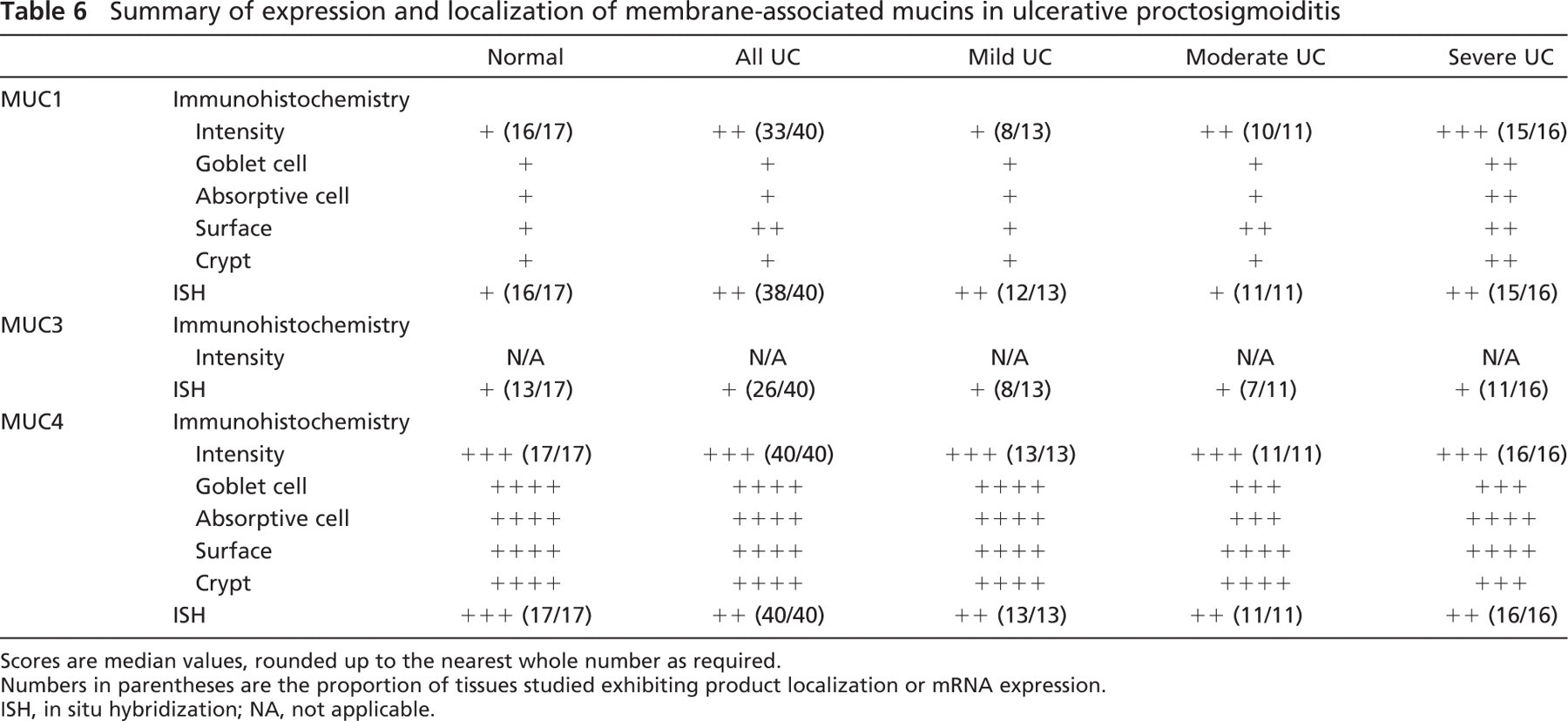
Scores are median values, rounded up to the nearest whole number as required.
Numbers in parentheses are the proportion of tissues studied exhibiting product localization or mRNA expression.
ISH, in situ hybridization; NA, not applicable.
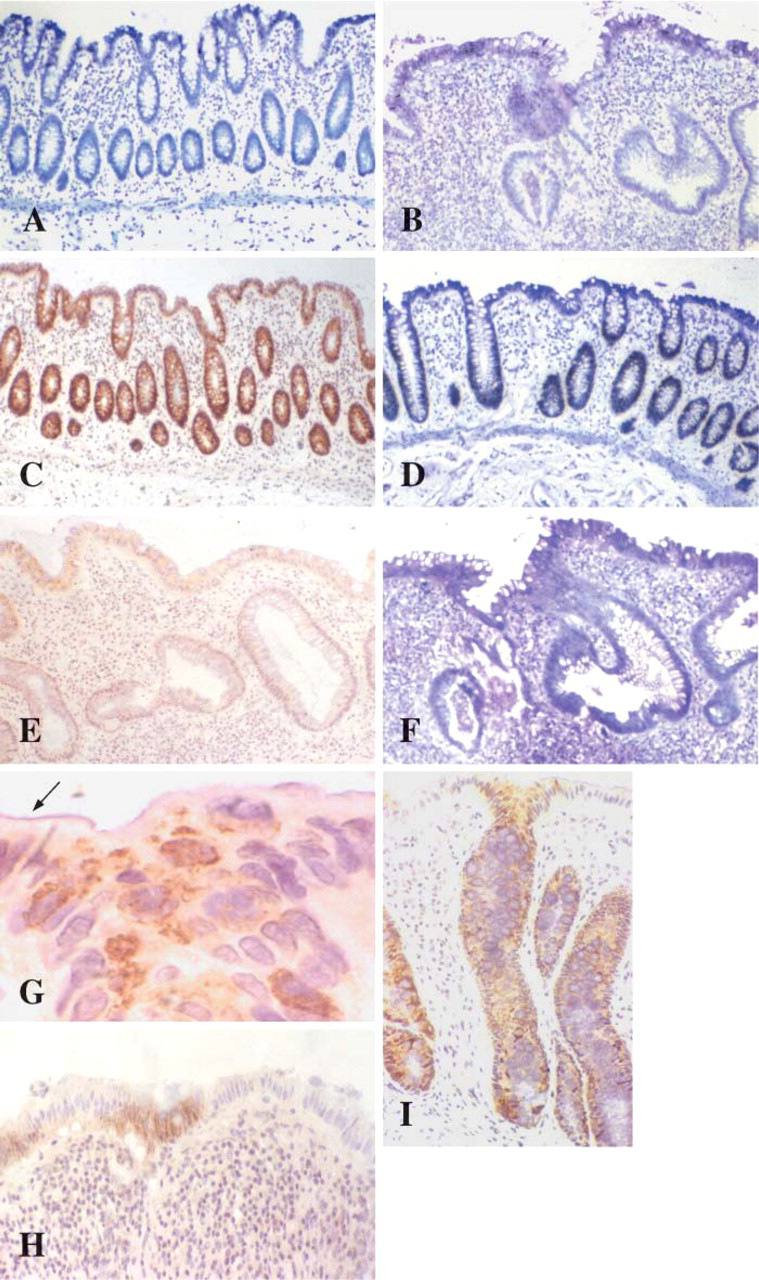
MUC3 mRNA expression in normal (
Trefoil Peptide Gene and Product Expression
TFF1 mRNA and peptide were absent from all normal colorectal biopsy specimens. In colitis, however, TFF1 mRNA was detected in 12/40 specimens and peptide was detected immunohistochemically in 10 of these specimens (Table 9; Figures 5A and 5B). All colitic tissues exhibiting TFF1 gene expression possessed a histological severity score of >3 [Bristol Index (Warren et al. 1988)], and eight of these tissues were from patients with clinically severe disease. In the colitic tissues exhibiting TFF1 staining, localization of the peptide was to goblet cell vesicles predominantly at the upper half of crypts (Figure 5C) and in probable restitutive mucosa at the luminal surface (Figure 5D).
LUM2–3 non-variable number tandem repeat antibody detects MUC2 glycoprotein localized to the goblet cell vesicles in normal (
No TFF2 gene mRNA or peptide product was detected in any of the colorectal tissues studied (Table 9).
Summary of expression and localization of secreted mucins in ulcerative proctosigmoiditis
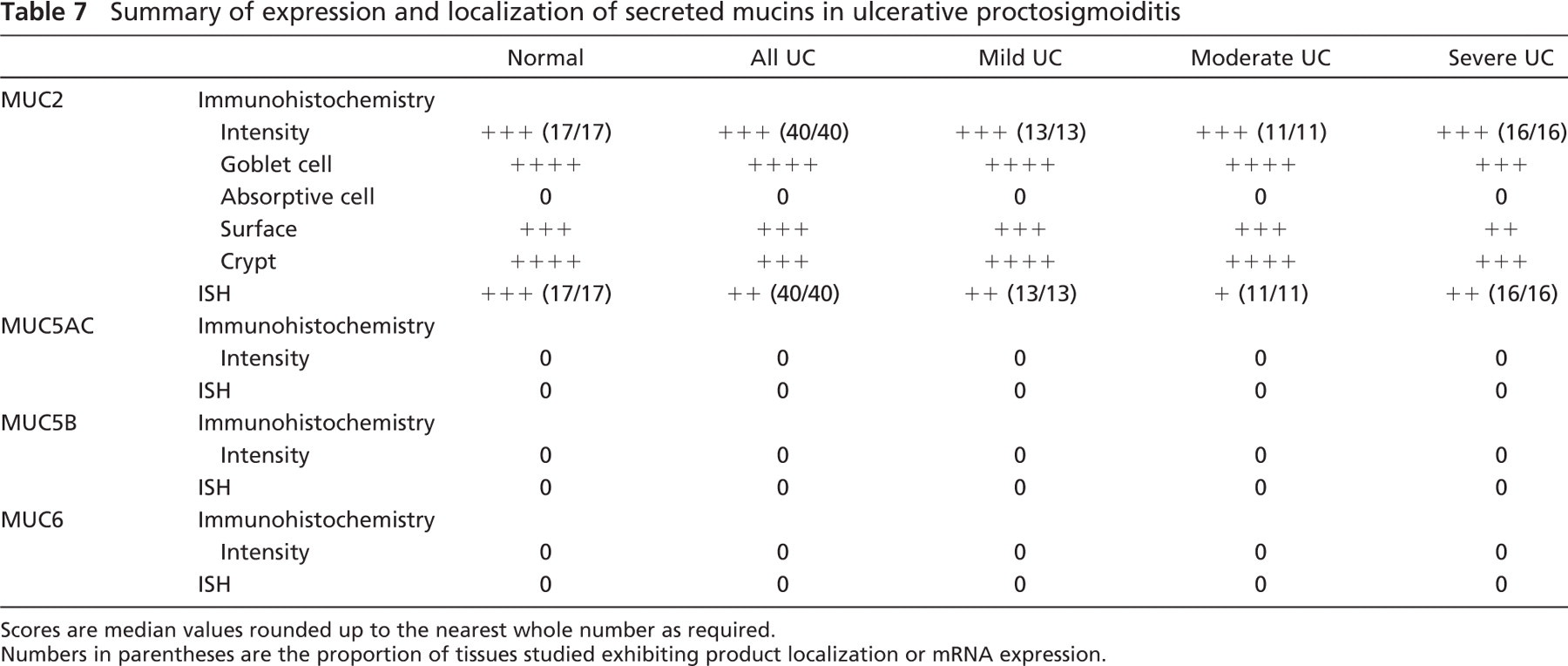
Scores are median values rounded up to the nearest whole number as required.
Numbers in parentheses are the proportion of tissues studied exhibiting product localization or mRNA expression.
The indigenous trefoil peptide to the large bowel mucosa, TFF3, was detected by ISH and IHC in all colorectal biopsies studied. TFF3 peptide was localized to goblet cell vesicles with preponderance for the lower half of crypts in both normal (Figures 5E and 5F) and diseased colorectal mucosa (Table 9). Whereas predominant localization was to goblet cell vesicles, weak goblet cell cytoplasmic staining was also identified in UC tissues. In UC a reduction in staining intensity was apparent, and a relative decrease of staining in the goblet cell population occurred at the luminal surface and upper half of crypts, a finding that was more pronounced with increasing severity of disease (Figures 5G–5I).
Discussion
This study has demonstrated alterations in the composition of the supramucosal defense barrier in UC, and such alterations vary in relation to clinical severity of the disease. Most notably there is an upregulation in mucin MUC1 and neo-expression of the trefoil peptide TFF1 in severe disease.
MUC1 glycoprotein, whose gene has been mapped to chromosome 1q21 (Gum et al. 1990), is a transmembranal mucin expressed by a number of mucosal tissues where it is localized to the apical surface of epithelial cells (Zotter et al. 1988). The physiological role of MUC1 in the colonic epithelium is not certain. MUC1 may play an important role in mucosal protection, cell-cell adhesion, immunoregulation, and cell-signaling processes in the epithelium of the colorectum (Fung and Longenecker 1991; Ligtenberg et al. 1992a; Regimbald et al. 1996; Nakagawa et al. 1999; Ren et al. 2004.)
This study has shown that MUC1 expression is upregulated in severe UC, with localization at the sites of potential or actual crypt abscess rupture. Previous reports using ISH did not detect any increased expression (Myerscough et al. 1995), and the current result may reflect the severity of the disease in the group of patients studied. Although overexpression of MUC1 has been widely described in neoplastic tissues, this is the first report of overexpression in UC.
The BC2 anti-MUC1 antibody used in this study is directed against the highly glycosylated VNTR region of the MUC1 protein core. The substantial colocalization of MUC1 mRNA and immunoreactive MUC1 in ulcerative colitic crypt abscesses would suggest that the detection of MUC1 product is not the result of a marked reduction in glycosylation of the MUC1.
Summary of localization of the sulfo-Lewisa mucin carbohydrate epitope in ulcerative proctosigmoiditis

Scores are median values, rounded up to the nearest whole number as required.
Numbers in parentheses are the proportion of tissues studied exhibiting product localization or mRNA expression.
Summary of expression and localization of trefoil peptides in ulcerative proctosigmoiditis

Scores are median values rounded up to the nearest whole number as required.
Numbers in parentheses are the proportion of tissues studied exhibiting product localization or mRNA expression.
Splice variants of MUC1 have been isolated in breast cancer, which yield MUC1 products with no VNTR domain (MUC1/Y) and with no membrane anchor (MUC1/SEC) (Zrihan-Licht et al. 1994; Smorodinsky et al. 1996). Only the MUC1/SEC form would be detected using the antibody used in this study. It is not possible, however, to discriminate between normal and MUC1/SEC variants in our study. Cellular expression of these forms may be reflected in alterations of subcellular distribution but would require biochemical separation to identify the mature proteins (Ligtenberg et al. 1992a) or mRNA analysis to identify the splice variants. Any MUC1/SEC shed from the apical surface would not be detected in this histological study. The relevance to this study of MUC1 splice variance and gene polymorphism is unknown.
The presence of MUC1 in the cell membrane of mucosal epithelia is significant. The dimensions of extended mucin molecules in the glycocalyx have suggested a range of interactions with secreted mucin gels, secreted proteins and effector molecules, bacteria, viruses, and other microorganisms (Patton et al. 1995; van Klinken et al. 1995). Cell-cell interactions may arise through mutual repulsion due to sialic acid negative charge (Ligtenberg et al. 1992b; Wesseling et al. 1996) or in the case of lectin binding, such as the intercellular adhesion molecule ICAM-1 where enhanced interaction is found (Regimbald et al. 1996).
Understanding of the mechanism of MUC1 upregulation at sites of crypt abscess rupture contributes to the understanding of the disease process in UC. A role for MUC1 in UC can been proposed at several levels including overproduction of modified mucin, alterations in cellular signaling processes, and modified cell-cell interactions. The location of MUC1 in the deep crypt may relate to the relative paucity of secreted mucus gel and contribute to surface protective properties (Winterford et al. 1999). Production of aberrant mucin may contribute to the loss of normal protective function, whereas overexpression of abnormal mucin provides an immunological target for the action of anti-mucin antibodies on colonic cells. Detection of circulating anti-MUC1 antibodies in UC patients has provided some support for the latter proposal (Hinoda et al. 1993; Hayashi et al. 2001).
Unlike Crohn's disease of the small bowel, there appears to be no neo-expression of the chromosome 11 mucins MUC5AC, MUC5B, or MUC6 (Buisine et al. 1999). Small bowel expression of MUC5AC, MUC5B, and MUC6 occurs in areas of ulcer-associated lineage cell lineage, and we advocate that this metaplastic phenomenon does not exist in the inflamed colorectal mucosa (Longman et al. 2000b; Longman and Warren 2000). MUC5AC expression certainly occurs in colorectal neoplastic conditions (Buisine et al. 1996; Longman et al. 2000a). Recently, however, sporadic MUC5AC localization without evidence of MUC5AC mRNA expression has been demonstrated in childhood UC (Shaoul et al. 2004). An explanation for this finding remains uncertain.
A decrease in the immunostaining of the sulfo-Lewisa mucin epitope is consistent with previous evidence of a loss of sulfation of mucins in UC (Corfield et al. 1996). Sulfo-Lewisa, however, is only one of a number of sulfated mucin epitopes found in the colorectum, and this study has not investigated alterations of the non-sulfated Lewisa epitope. Staining pattern of the F2 antibody, however, demonstrated a similar pattern to that found with total sulfomucin localization as exhibited by HID staining.
A reduction in the expression of the trefoil peptide, TFF3, has previously been identified in the mucosa of UC patients (Podolsky 1997). This study has demonstrated a reduction in goblet cell TFF3 localization occurring at the surface of remaining colonic crypts and in regenerative epithelium. Trefoil peptides are potent motogens to the gastrointestinal epithelia (Kindon et al. 1995; Playford et al. 1995) and aid in the protection and healing of gastrointestinal mucosa (Playford et al. 1995; Babyatsky et al. 1996; Konturek et al. 1997; McKenzie et al. 1997). In TFF3 gene knockout animals, repletion of recombinant TFF3 has been shown to aid recovery from chemically induced colitis (Mashimo et al. 1996). Although the precise mechanisms of action of the TFF peptides are unclear, they appear to have direct effects on apoptosis, cell differentiation, and cell-cell adhesion (Efstathiou et al. 1998; Taupin et al. 1999). This may suggest that the colonic epithelium in UC has either an inability to protect itself from injury or possesses a defective restitutional ability. Whereas MUC2 and TFF3 are colocalized in goblet vesicles in normal small and large bowel (Longman et al. 2000b), a decrease in goblet cell numbers expressing TFF3 in surface/regenerative colonic mucosa in UC may not be reflected by the MUC2 expression patterns. Neo-expression of TFF1 may have further important consequences in the pathophysiology of UC. TFF1 expresion in severe UC may be a reflection of the switch-on of protective or healing mechanisms within the colorectal mucosa. This certainly seems apparent in other regions of the gastrointestinal tract such as in the gastric epithelium or in the ulcer-associated cell lineage in the small bowel where TFF1 is aberrantly or newly expressed (Wright et al. 1990; Playford et al. 1995). There is, however, no associated neo-expression of TFF2 in UC.
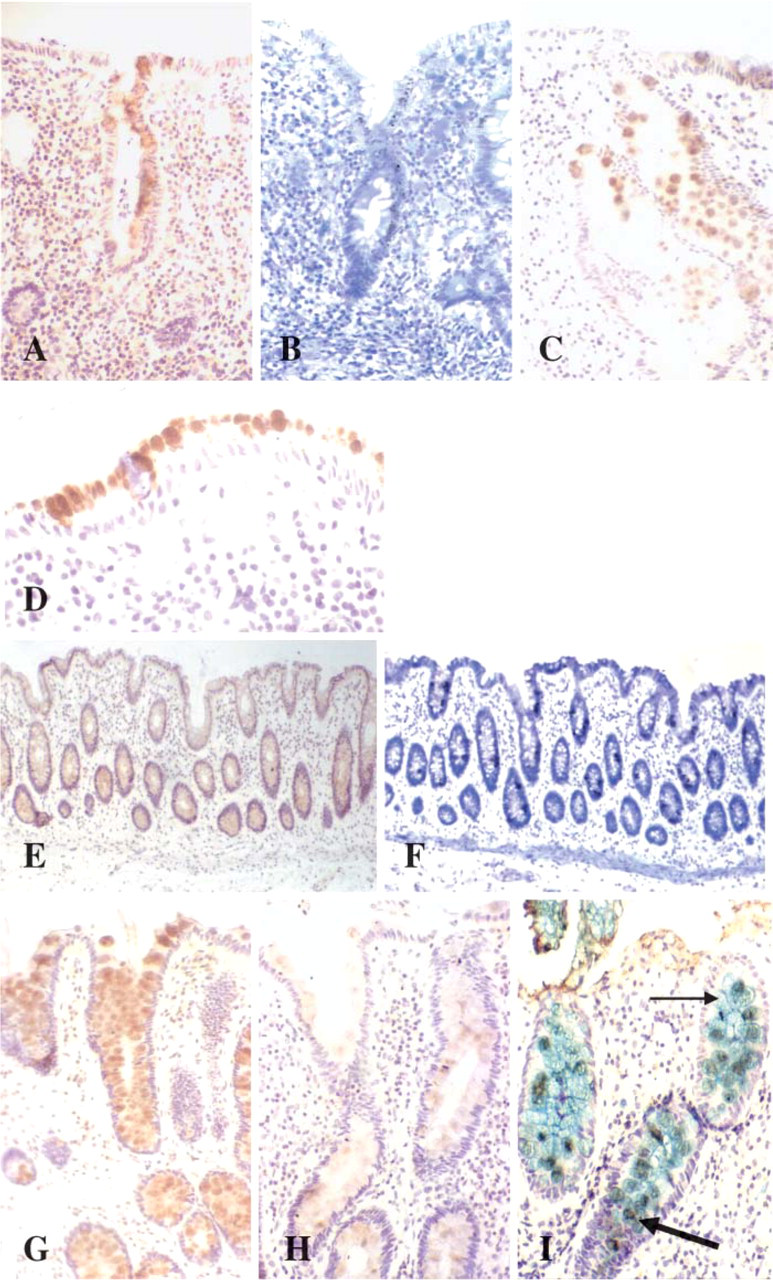
TFF1 peptide (
The importance of changes in mucin glycoprotein and trefoil peptide expression in UC remains uncertain but is likely to be a consequence of the disease process. Further investigation into the mechanisms of function and interaction is required before mucins or trefoil peptides can be excluded from playing a primary role in the inflammatory process. Mucin glycoproteins are a heterogeneous group of macromolecules, and a large number of different mucin genes are expressed by the colonic mucosa. The in vivo functions of all the mucin genes are as yet unclear. The more recent discovery of the chromosome 7q22 mucins, namely, MUC3A, MUC3B, MUC11, and MUC12, and also the chromosome 3q13.3 mucin MUC13, allows the possibility of investigating the role that they may play in UC (Williams et al. 1999; Pratt et al. 2000). Evidence that rare alleles of the previously known MUC3 gene occur in UC requires further elucidation to show how polymorphisms of this gene may be implicated in the disease (Kyo et al. 1999). The diversity and complexity of mucin glycoproteins, however, makes the investigation of their physiological purpose difficult. The importance of trefoil peptides in inflammatory bowel disease will only be fully realized when their epithelial and immunological functions, and their possible interactions with mucins, have been identified.
In summary, we have characterized the expression patterns of the transmembranal mucins MUC1, MUC3, and MUC4; the secreted mucins MUC2, MUC5AC, MUC5B, and MUC6; and the trefoil peptide TFF1, TFF2, and TFF3 in UC with relevance to disease severity. Specific neo-expression of TFF1 in severe disease and localization of MUC1 to sites of actual or potential crypt abscess rupture have been identified.
Footnotes
Acknowledgements
LUM2-3 and LUM5-1 antibody development was supported by the European Union consortium (grant CEEBMH4-CT98-3222).
Mucin glycoprotein antibodies LUM2-3, LUM5-1, and GPEP24 were kindly provided by Dr. I. Carlstedt (Lund University, Lund, Sweden). BC2 and M4.275 antibodies were kindly provided by Dr. M.A. McGuckin and Dr. M. Walsh (University of Queensland, Brisbane, Australia). MAN-5B1 antibody was kindly provided by Dr. J.K. Sheehan and Dr. D.J. Thornton (University of Manchester, Manchester, UK). F2 sulfo-Lewisa antibody was kindly provided by Dr. E.C.I. Veerman and Dr. A.V. Nieuw Amerongen (Vrije University, Amsterdam, The Netherlands). HM:169 antibody was kindly provided by Dr. K. Lynch-Devaney and Professor D.K. Podolsky (Inflammatory Bowel Disease Research Institute, Massachusetts General Hospital, Boston, MA).