
Abstract
The skeleton of demosponges is built of spicules consisting of biosilica. Using the primmorph system from Suberites domuncula, we demonstrate that silicatein, the biosilica-synthesizing enzyme, and silicase, the catabolic enzyme, are colocalized at the surface of growing spicules as well as in the axial filament located in the axial canal. It is assumed that these two enzymes are responsible for the deposition of biosilica. In search of additional potential structural molecules that might guide the mineralization process during spiculogenesis to species-specific spicules, electron microscopic studies with antibodies against galectin and silicatein were performed. These studies showed that silicatein forms a complex with galectin; the strings/bundles of this complex are intimately associated with the surface of the spicules and arranged concentrically around them. Collagen fibers are near the silactein/galectin complexes. The strings/bundles formed from silicatein/galectin display a lower degree of orientation than the collagen fibers arranged in a highly ordered pattern around the spicules. These data indicate that species-specific formation of spicules involves a network of (diffusible) regulatory factor(s) controlling enzymatic silica deposition; this mineralization process proceeds on a galectin/collagen organic matrix.
I
In Demospongiae, initiation of spicule formation starts intracellularly within specialized cells called sclerocytes. There an axial filament is assembled in organelles around which the first siliceous deposits are layered (Müller et al. 2006), thus forming the nucleus and initial growth form of a spicule. The first siliceous layers surrounding the axial canal measure 1-2 μm in diameter (Uriz et al. 2000). When reaching lengths of ~5-10 μm, spicules are extruded from the cells into the mesohyl where their final sizes and shapes are completed. Thickening of spicules proceeds by apposition of concentric silica layers (Uriz et al. 2000; Müller et al. 2006). In recent years, the molecular basis of spicule formation has been elucidated in some detail. Silicon is actively taken up by demosponges through a membrane-bound transporter, which can very likely be grouped to the family of Na+/HCO3 - cotransporters (Schröder et al. 2004a). Intracellularly, polycondensation of silicic acid is mediated enzymatically through the enzyme silicatein (Shimizu et al. 1998; Weaver and Morse 2003). Several isoforms of silicatein have been identified in Tethya aurantium and termed silicatein-α,-β, and -γ (Shimizu et al. 1998). Subsequently, silicateins have been identified in other demosponges such as Suberites domuncula (Müller et al. 2003,2006) and Lubomirskia baicalensis (Kaluzhnaya et al. 2005).
Forms and sizes of spicules are highly diverse (Boury-Esnault and Rützler 1997). Because the morphology of the spicules differs species-specifically, they are used as a major taxonomic character. However, very little is known about the molecular mechanism(s) that determine the position and arrangement of the spicules within the demosponge skeleton (Uriz et al. 2003). Based on experimental evidence, it was proposed that collagen fibrils, adhesive proteins, and polysaccharides are those elements that transport the spicules to their functional position within the sponge body (Garrone 1978,1984,1998). However, structural macromolecules that regulate and control the growth of spicules remained obscure, very likely because growth of spicules is a rapid process. It was calculated that longitudinal growth of spicules proceeds at a velocity of 5 μm/hr (Weissenfels 1989). Investigation of spicule formation under controlled conditions became possible after the successful establishment of the sponge tissue culture system, a special form of three-dimensional cell aggregates. These aggregates are composed of proliferating and differentiating cells (termed primmorph system) (Müller et al. 1999).
In the present study, using microscopic analyses we show that the two enzymes, silicatein, the major biosilica-forming enzyme (Weaver and Morse 2003), and silicase, the biosilica-catabolic enzyme (Schröder et al. 2003), are colocalized during synthesis of silica depositions. Furthermore, microscopic data are presented indicating that two structural molecules, galectin and collagen, are involved in control of growth and shape formation of spicules. Our data suggest that galectin organizes silicatein molecules to form primary strings/bundles and allows the matrix-guided deposition of the first biosilica deposits. In a second step, these structures are directed by collagen fibers very likely in a concentric pattern around the spicules. The view that spicule formation is under the control of regulatory factors came from the identification of the sponge osteoclast-stimulating factor. The first factor, potentially involved in osteoclast formation, is this osteoclast-stimulating factor whose sequence was reported in the database (accession number AAH07459). Data presented in the manuscript experimentally support the view (Weiner and Dove 2003) that mineralization of spicules is a biologically controlled extracellular process.
Materials and Methods
Materials
Restriction enzymes and reagents for RACE procedure were purchased from Invitrogen (Carlsbad, CA). Cy3-conjugated F(ab')2 goat anti-rabbit IgG were purchased from Jackson ImmunoResearch (Cambridgshire, UK).
Sponges and Primmorphs
Specimens of the marine sponge Suberites domuncula (Porifera, Demospongiae, Hadromerida) were collected in the Northern Adriatic near Rovinj, Croatia and then kept in aquaria in Mainz, Germany at a temperature of 17C for >5 months. In one series of experiments, a tissue sample from a specimen was treated for 30 min in seawater supplemented with 30 mM EDTA at room temperature. Spicules were then obtained by sedimentation and processed for in situ hybridization.
The in vitro primmorph cell system was used (Krasko et al. 2000). Primmorphs were kept at 17C in natural seawater (enriched with 60 μM of silicate) and supplemented with 1% RPMI 1640 medium. Approximately 20 days later the primmorphs were used for analysis.
Cloning of the Osteoclast-stimulating Factor
In a screening program for the osteoclast-stimulating factor, degenerate primers deduced from the conserved first ankyrin domain of the human osteoclast-stimulating factor (accession number AAH07459), spanning the stretch aa76 to aa82, were used. Polymerase chain reaction was performed on the total S. domuncula cDNA library prepared in a Lambda ZAP Express vector (Kruse et al. 1997). Fragments of the expected size were obtained, cloned, and sequenced. The sequence was completed with insert-specific primers in combination with 5′-RACE primer or with 3′-RACE primer using the CapFishing Full-length cDNA Premix Kit (Seegene; Rockville, MD). The final sequence was confirmed by an additional PCR using primers directed against the non-translated region of the cDNA, followed by sequencing. The clone encoding S. domuncula cDNA for the osteoclast-stimulating factor (SDOSTF) is 739-nt long; cDNA for the putative SDOSTF has been deposited (EMBL/GenBank) under AM167903.
Preparation of Antibodies
Polyclonal antibodies (PoAb) were raised against purified, recombinant silicatein (rSILIC_SUBDO), recombinant galectin (rGALEC_SUBDO), and recombinant silicase (rSIA_SUBDO) in female rabbits (White New Zealand) as described (Schröder et al. 2003; Wiens et al. 2003; Müller et al. 2005). Titers of the antibodies PoAb-aSILIC, PoAb-aGALEC, and PoAb-aSIA were >1:3000.
Electron Microscopy
Transmission Electron Microscopic (TEM) Analysis. For TEM, primmorph specimens were cut into pieces and incubated in 0.1 M phosphate buffer (supplemented with 2.5% glutaraldehyde and 0.82% NaCl, pH 7.4) and finally washed in 0.1 M phosphate/1.75% NaCl buffer, pH 7.4, at room temperature. After treating the samples with 1.25% NaHCO3, 2% OsO4, and 1% NaCl, they were dehydrated with ethanol. Dried samples were incubated with propylene oxide, fixed in propylene oxide/araldite (2:1), covered with pure araldite, and hardened at 60C for 2 days prior to cutting into 60-nm ultrathin slices (Ultracut S; Leica, Wetzlar, Germany). Samples were transferred onto coated copper grids and analyzed with a Tecnai 12 microscope (FEI Electron Optics; Eindhoven, The Netherlands).
Electron Immunogold Labeling. Primmorph samples were treated in 0.1% glutaraldehyde/3% paraformaldehyde buffered in 0.1 M phosphate buffer, pH 7.4. After 2 hr the material was dehydrated in ethanol and embedded in LR-White Resin. Sixty-nm-thick slices were cut and blocked with 5% BSA in PBS and then incubated with the primary antibody PoAb-aSILIC (1:1000) or PoAb-aGALEC (1:1000) for 12 hr at 4C. In controls, preimmune serum was used. After three washes with PBS/1% BSA, sections were incubated with 1:100 dilution of the secondary antibody (1.4-nm nanogold anti-rabbit IgG; diluted 1:200) for 2 hr. Sections were rinsed in PBS, treated with 1 % glutaraldehyde/PBS for 5 min, washed, and dried. Subsequently, immunocomplex enhancement was performed with silver as described (Danscher 1981). Samples were examined as above with the Tecnai 12 microscope.
Scanning Electron Microscopic (SEM) Analysis. Sponge specimens were analyzed by SEM, and samples were mounted on stubs (carbon adhesive Leit-Tabs #G 3347; Plano, Wetzlar, Germany) and observed with a Zeiss DSM 962 digital scanning microscope (Zeiss; Aalen, Germany).
In Situ Localization Studies
The method applied bases on a described procedure (Polak and McGee 1998; Perović et al. 2003). Eight-μm-thick frozen sections through tissue were prepared, fixed, treated with proteinase K, and subsequently fixed again with paraformaldehyde. After rehydration, sections were hybridized with the labeled probe, a 200-nt-long SDOSTF cDNA portion. After blocking, sections were incubated with an anti-digoxigenin antibody conjugated with alkaline phosphatase. The dye reagent NBT/X-Phosphate was used for visualization of the signals. Antisense and sense ssDNA digoxigenin-labeled probes were synthesized by PCR using the PCR DIG Probe synthesis Kit (Roche; Mannheim, Germany). Sense probes were used in parallel as negative controls in the experiments.
The cells surrounding and associated with spicules were isolated from sponge tissue as described and hybridized under the same conditions with the collagen probe. A recently described cDNA encoding fibrillar collagen-3 (SDCOLL3) was used (Schröder et al. 2006), accession number AM157177, choosing a 400-nt portion comprising the collagen triple helix repeats.
Immunohistology
Primmorph/tissue samples were embedded and 8-μm frozen sections were obtained as described (Perović et al. 2003). Cryosections were fixed, washed and then incubated with the first antibody (PoAb-aSILIC or PoAb-aSIA; 1:1000 dilution), blocked and then subjected with the second antibody (Cy3-conjugated F(ab')2 goat anti-rabbit IgG (1:2000 dilution). Immunofluorescence analysis was done with an Olympus AHBT3 light microscope (Olympus; Tokyo, Japan) together with an AH3-RFC reflected light fluorescence attachment at the emission wavelength 546 nm (filter G).
Results
Co-localization of Silicatein and Silicase in Primmorphs
Cryosections through primmorphs were prepared and stained with PoAbs raised against recombinant silicase (PoAb-aSIA) and silicatein (PoAb-aSILIC) from S. domuncula. Inspection by fluorescence light microscopy revealed that these two antibodies stained fibrillar structures in primmorphs (Figures 1A and 1C). In parallel, these long-fibered structures had been visualized by Nomarsky optics (Figures 1B and 1D).
To support that silicase and silicatein proteins are recognized by antigens associated with the spicules, sections through tissue were performed. After immunostaining, strong signals resulting from reactions with PoAb-aSIA (Figure 1E) or with PoAb-aSILIC (Figure 1F) could be detected. Immunocomplexes were seen on the surface of the spicules and in the axial filaments residing in the axial canals.
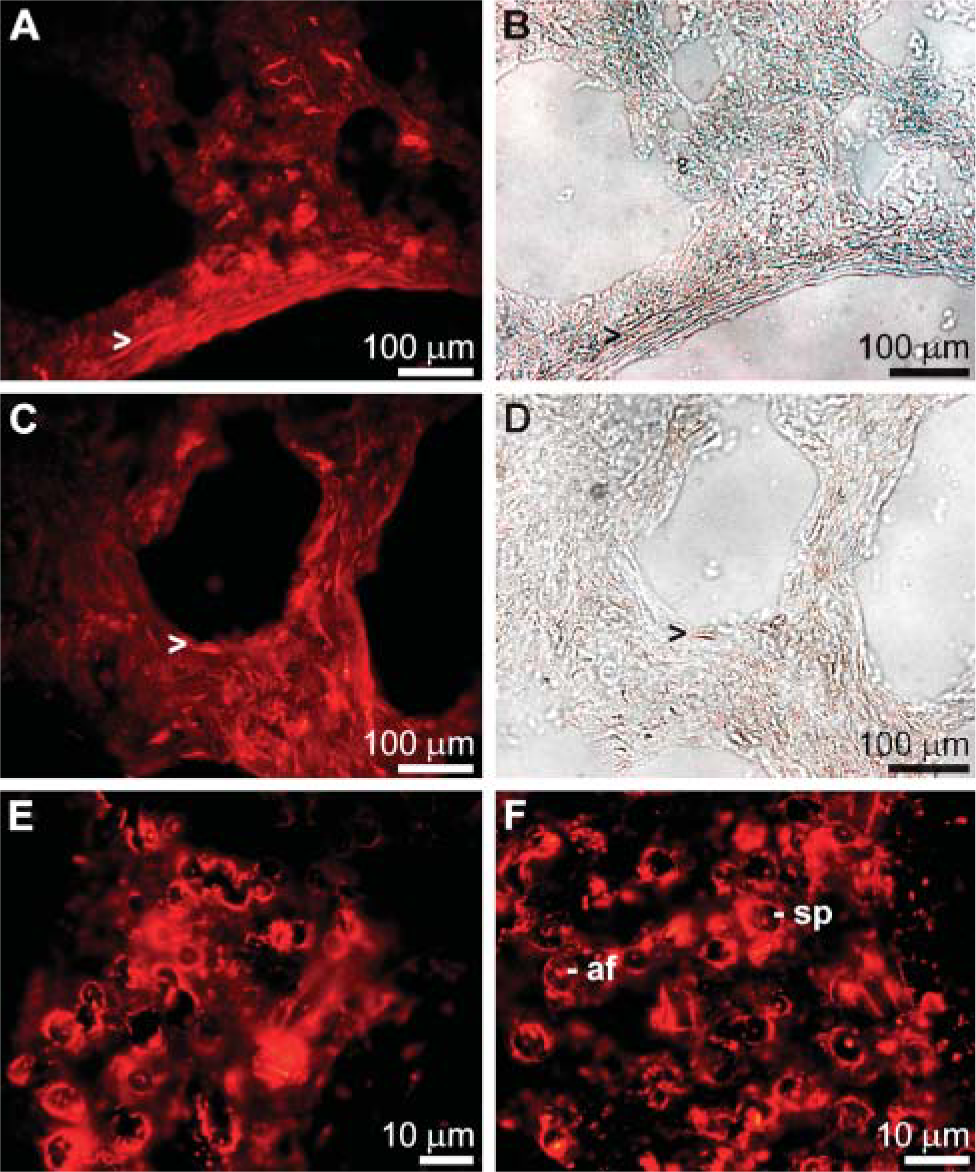
Immunostaining of cryosections through primmorphs (
Osteoclast-stimulating Factor, the Gene/Protein, and Its Expression
The S. domuncula polypeptide encoding the putative osteoclast-stimulating factor was identified and cloned from the cDNA library of S. domuncula. The 739-nt-long sequence (SDOSTF) has an open reading frame nt70/72 to nt703/705 (stop) that codes for the 211-aa-long protein (OSTF_SUBDO; Figure 2A). The size of this molecule is 23,049 Da. Motif scan analysis revealed three ankyrin repeats (aa76 to aa108, aa109 to aa142, and aa143 to aa175) as well as one SH3 motif. The highest sequence similarity is seen with the human protein, the osteoclast-stimulating factor 1, with 66% similar and 53% identical amino acids. The similarity score to the protostomian sequence from Caenorhabditis elegans (osteoclast-stimulating factor 1) is lower. Similarities to the Drosophila melanogaster-deduced protein (CG4719-PA) and the probable ankyrin protein from Arabidopsis thaliana are <50%. This relationship is also reflected by the rooted phylogenetic tree (Figure 2B).
In situ hybridization studies were performed to elucidate in which cells the gene for the osteoclast-stimulating factor is expressed. Applying the antisense probe (SDOSTF), strong signals are seen in cells surrounding the spicules (Figures 2C-2C' and 2C”). Controls have been performed with the sense probe, which gave no signal after hybridization (Figures 2C-2C'”).
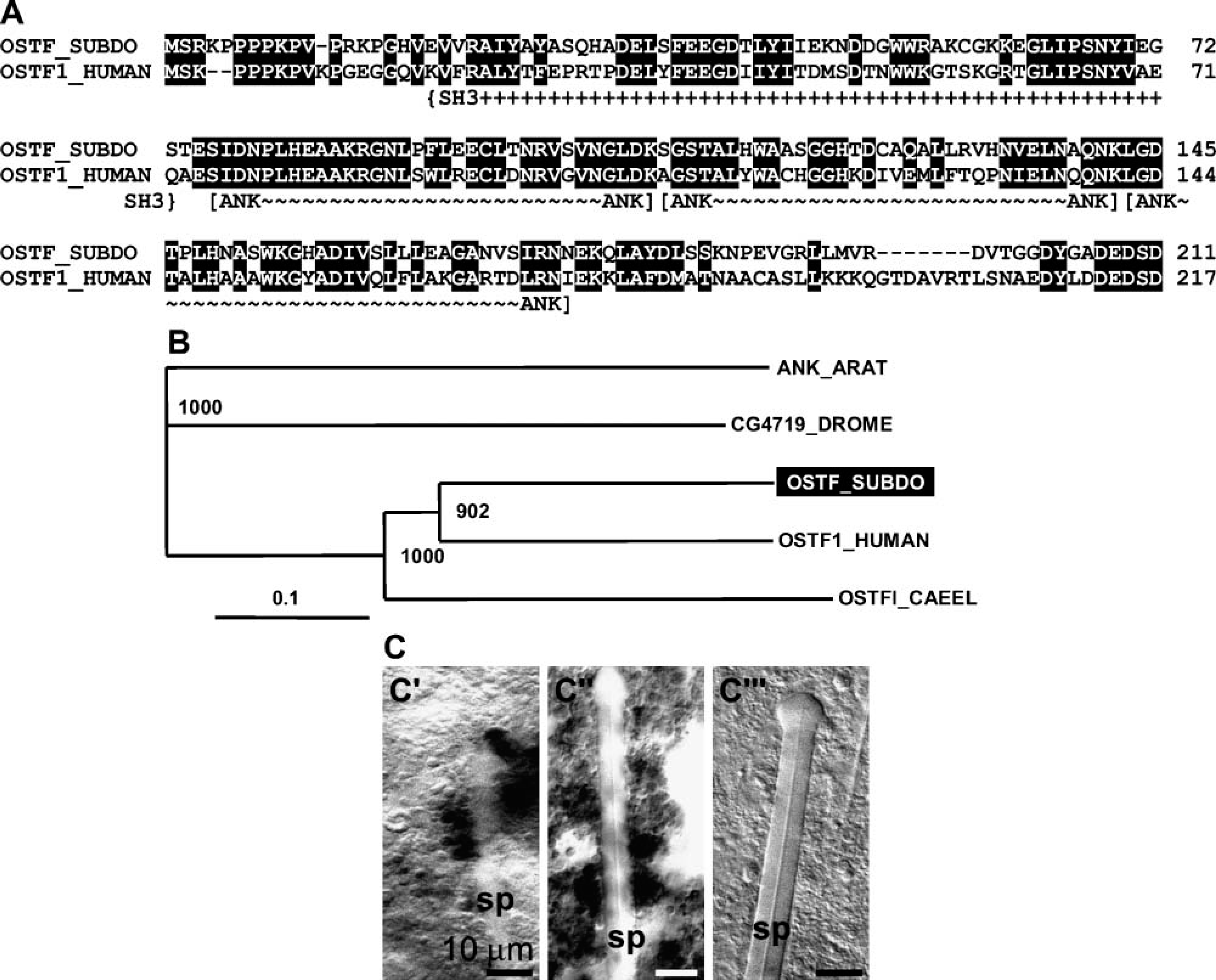
Osteoclast-stimulating factor in S. domuncula. (
Extra-spicular Association of Silicatein with Galectin
Using antibodies against silicatein (PoAb-aSILIC) and galectin (PoAb-aGALEC), the association of silicatein with galectin in the extraspicular space in primmorphs was determined by electron immunogold labeling. Sections through primmorphs were reacted with antibodies against silicatein (Figures 3A-3C) and galectin (Figures 3E-3H). The gold signals coming from the PoAb-aSILIC/gold complexes are arranged around the cross-section through spicules in primmorphs in concentric patterns (Figures 3A and 3B). At higher magnification it can be distinguished that the rings are composed of strings/bundles that react with the PoAb-aSILIC/gold complexes (Figure 3C). The siliceous shells of the growing spicules are visible as electron-dense deposits around the axial filament (Figure 3B). In sections that were performed at the tip of a growing spicule, little of the siliceous shell can be recognized (Figure 3A). To demonstrate that the immunoreactions are specific, preimmune serum was used. In those images no gold complexes are seen (Figure 3D).
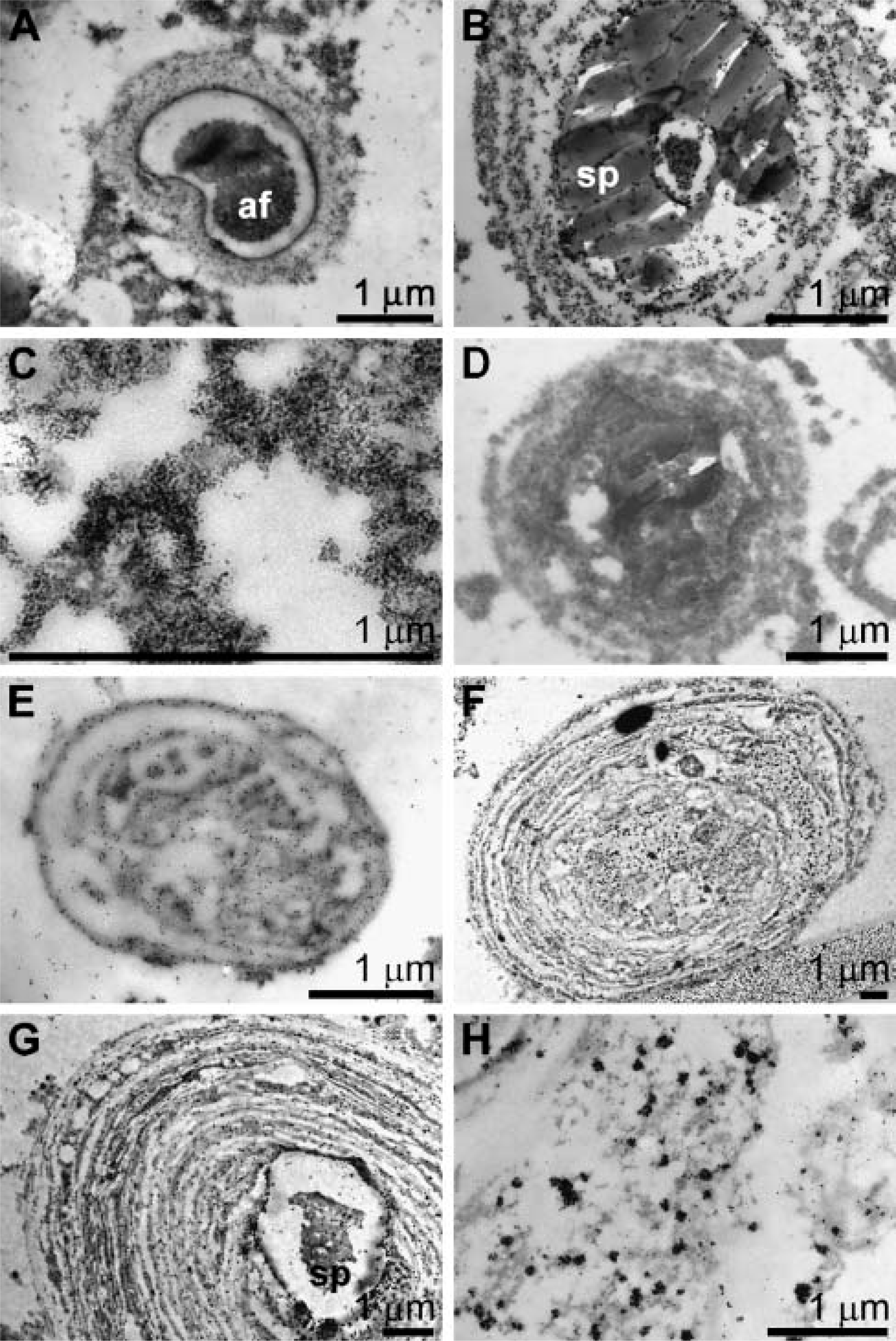
Association of silicatein with galectin in the extraspicular space in primmorphs; analysis by electron immunogold labeling. Sections through primmorphs were performed and reacted with antibodies against silicatein (PoAb-aSILIC) (
In parallel to the anti-silicatein antibodies, antibodies against galectin were applied. The reaction of these antibodies is visible in those structures that surround the spicules concentrically (Figures 3E-3H). At higher magnification the accumulated gold complexes, associated with electron dense strings/bundles, are clearly detected (Figure 3H). Control studies with preimmune serum gave no signals here (not shown).
Localization of Collagen in the Vicinity of Spicules
TEM of osmium-fixed samples was performed to visualize the collagen network (Figure 4). Again the primmorph system was used to analyze spicules during their growing phase. After release into the mesohyl, spicules are only occasionally associated with cells (Figure 4A). In most images the spicules are surrounded by the bulky extracellular matrix of the mesohyl into which the fibers are embedded (Figures 4B-4C). At an average magnification an organized network of collagen fibers can be seen, especially at the tip of growing spicules (Figure 4B) arranged longitudinally to the spicules. In the center of the spicules both cross-sections through collagen fibers and through longitudinally orientated fibers are seen (Figure 4C). At higher magnification the less fibrous structured strings/bundles are visualized, located between the collagen fibers and the spicules (Figures 4D-4F). Finally, at highest magnification it is apparent that the strings are closely associated with the siliceous spicules, whereas collagen fibers end at a distance of 150 μm in front of the spicules (Figures 4G-4I).
Organized Collagen Web Surrounds Spicules
SEM analysis was applied to demonstrate the complex collagen network surrounding the spicules within tissue from adult specimens (Figure 5). An organized collagen web surrounds the spicules (Figure 5A); the images show fibers that follow the orientation and the shape of the spicules. Only at the ends of the spicules, which in adult specimens are tylostyles where the knobs have formed, a mesh-like arrangement of the collagen fibers is seen (Figures 5B and 5C). In the midportion of the spicules, less filamentous strings are seen, which are layered between the collagen fibers and the surface of the spicules (Figures 5D and 5E).
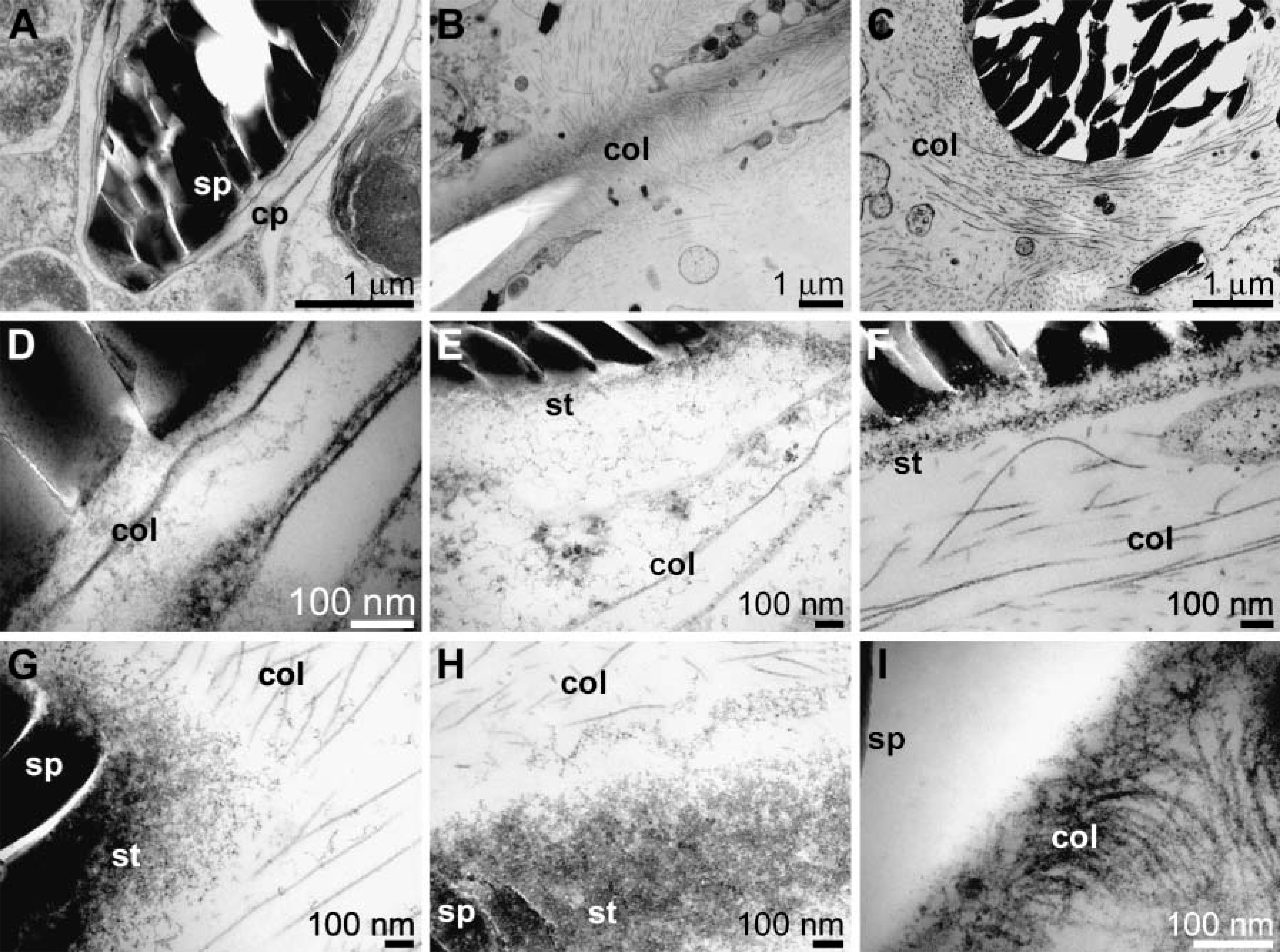
Transmission electron microscopy analysis of primmorphs. Osmium-fixed samples were processed as described under Materials and Methods. (
Expression of Collagen in Cells Associated with Spicules
To demonstrate that the cells surrounding the spicules express collagen, spicule samples were prepared with their associated cells. After in situ hybridization with a probe for the fibrillar collagen-3 gene from S. domuncula, strong signals are seen after the reaction with the antisense probe (Figure 6A), whereas the sense probe reacted only weakly (Figure 6B).
Discussion
Mineralization of those structures in metazoans that have a species-specific form is biologically controlled; this implies that a differential gene activity controls the nucleation, growth, and final localization of the mineral deposits (Weiner and Dove 2003). It is known from vertebrate bones that the individual morphology of the bones is the result of tuned modeling and remodeling processes under genetic as well as epigenetic control (Garant 2003). The earliest metazoans, the siliceous sponges, are composed of species-specific and highly complex skeletal elements, the spicules. During the past few years, the first clues on the genomic regulatory systems that control spiculogenesis have been described. These animals having a distinct body plan and, in at least some species, a pronounced axis do not contain homeobox gene clusters. However, the genetic repertoire of Porifera is complex enough to allow the formation of amazingly diverse spicules, the structural basis for the species-specific phenotypes (Müller 2005). Axis formation, e.g., in branches from some species, is controlled by differential expression of an apoptotic gene, as demonstrated for L. baicalensis (Wiens et al. 2006). At present, however, the processes through which the cells control the spatial and temporal pattern of spicule formation are not known. Recent results of in vitro studies suggested that anabolic (silicatein) as well as catabolic enzymes (silicase) are involved in the establishment of the well-structured inorganic skeletal elements, at least in demosponges (Schröder et al. 2003). In addition, it can be postulated that matrix-mediated signaling and growth/differentiation factors regulate spiculo-/skeletogenesis as well.
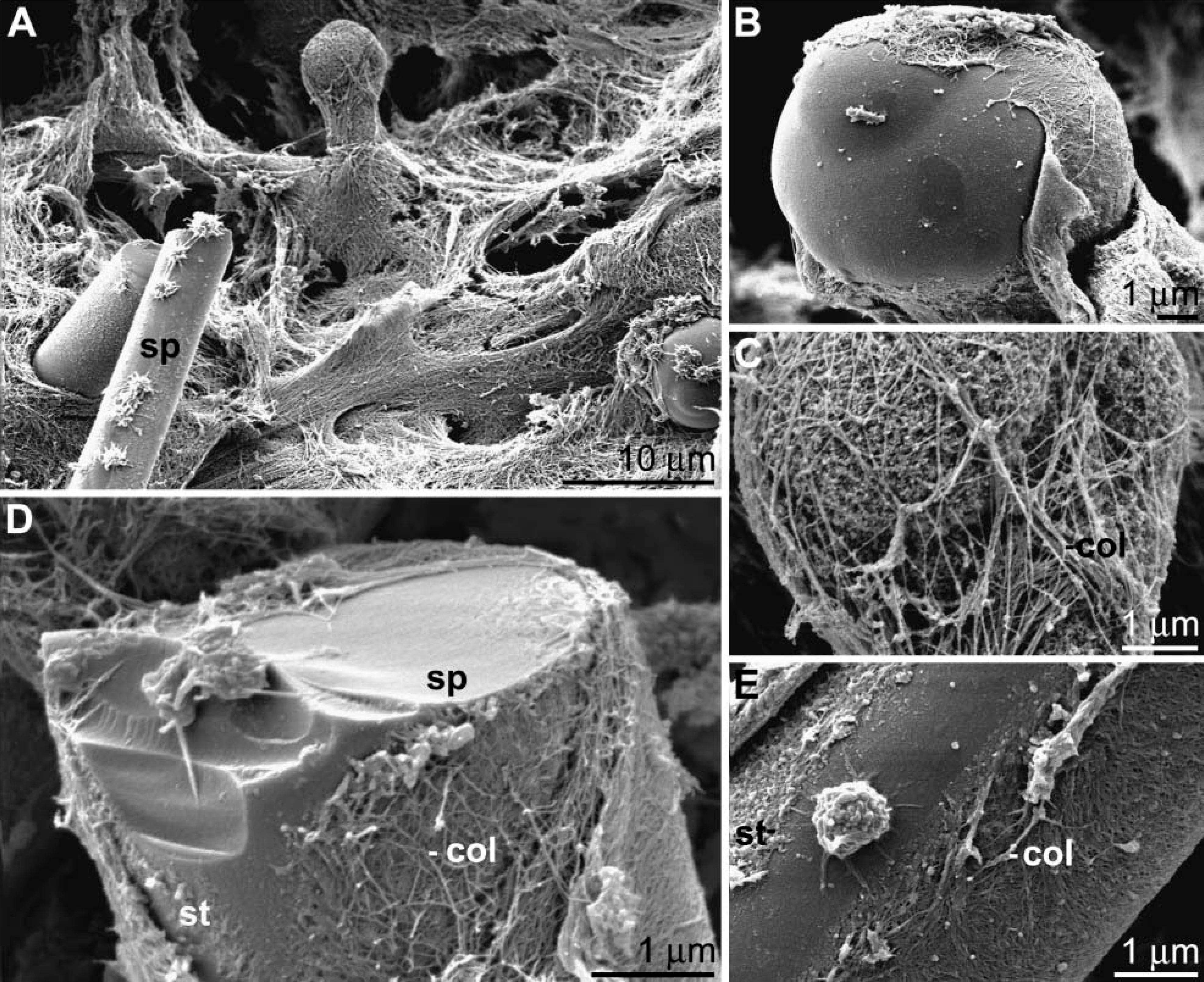
Scanning electron microscopy (SEM) analysis through tissue. Collagen fibers surround the spicules (sp) in an organized manner (
The studies presented herein show that during spicule formation in primmorphs of S. domuncula, expression of both silicatein and silicase is seen at those sites within the three-dimensional cell aggregates where the growth/building of these siliceous structures starts. Silicateins present in the axial filament of the spicules are enzymes that synthesize biosilica in vitro (Cha et al. 1999; Krasko et al. 2000,2002) and belong to the family of proteolytic enzymes (cathepsins). The role of the silicateins as synthesizing enzymes in vivo was elucidated by in situ hybridization studies (Schröder et al. 2004b). In the present study, immunofluorescence analyses have been applied to demonstrate that in primmorphs silicatein is associated with the fibrous structures that represent the primordial spicules. In parallel with the application of anti-silicatein it could be shown that the silica-catabolizing enzyme (silicase) is also localized at the same structures within the primmorphs. This result was supported by immuno-fluorescence studies of sections through tissue samples. Like silicatein(s), silicase belongs to a protein family known as carbonic anhydrases (Hatch 1980; Martin and Udagawa 1998; Schröder et al. 2003). Silicase has been shown to mediate dissolution of biosilica (Schröder et al. 2003). From this finding, a functional aspect can be deduced, suggesting that as in other inorganic skeletal structures such as in the osteons of the vertebrate bones (Garant 2003), mineralization/deposition of biosilica of the spicules is a result of synthesizing and catabolizing enzymes.
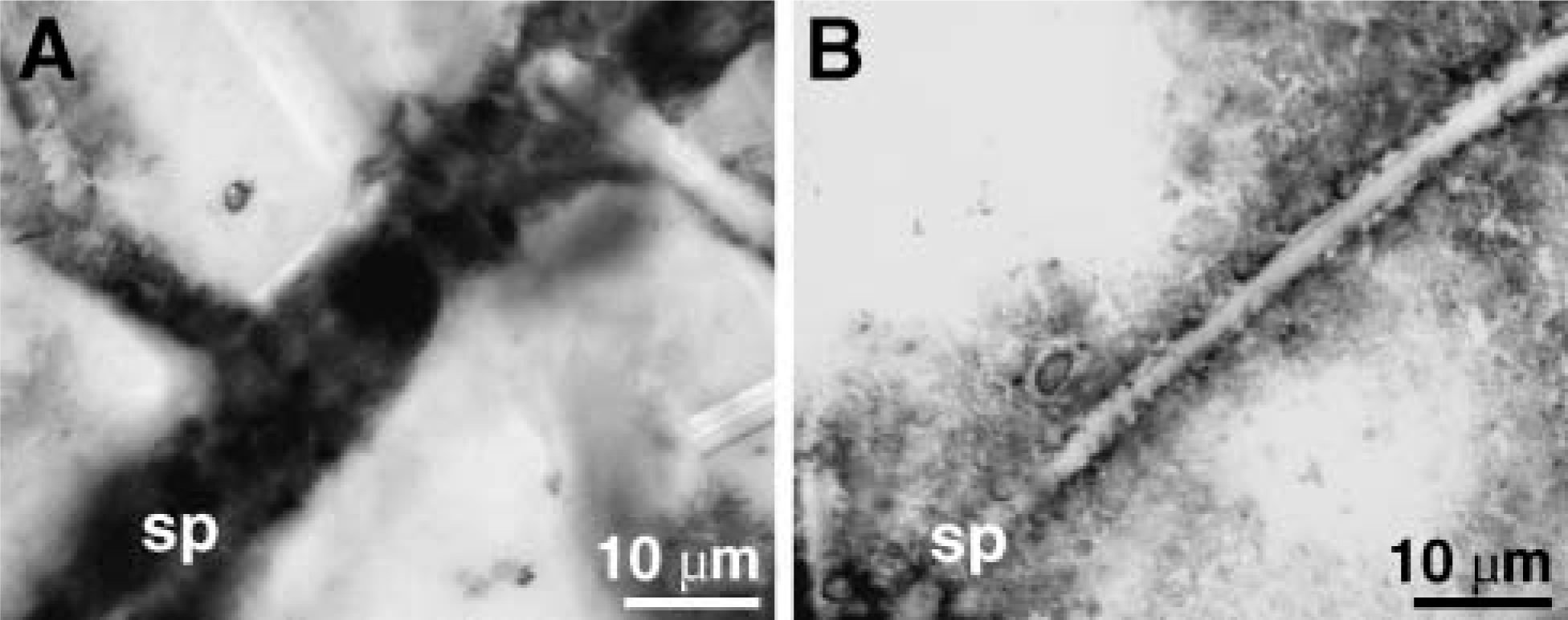
Expression of the gene encoding fibrillar collagen-3 in cells surrounding the spicules (sp); analysis by in situ hybridization. (
It remains enigmatic which factors/structural macro-molecules guide biosilica deposition to its functional location and allow the formation of the species-specific appearance of the spicules. Advances in tissue culture techniques in sponges and the analysis of the genome complexity of sponges, especially in S. domuncula, allowed for the first time the identification of a putative factor, presumably involved in spicule formation, the osteoclast-stimulating factor (OSTF_SUBDO). Osteoclast-stimulating factor is involved in the regulation of osteoblast/osteoclast activity together with cathepsin and carbonic anhydrases (Martin and Udagawa 1998; Quinn and Gillespie 2005). The cDNA/gene of this protein was identified and cloned in S. domuncula. The deduced protein comprises three ankyrin domains and one SH3 (Src homology 3) domain. Both domains are involved in protein-protein interactions in those molecules involved in signal transduction and cellular localization (McPherson 1999). Similarity of the sponge protein is highest to the corresponding human protein; similarity scores to related proteins from protostomians are lower, e.g., to the osteoclast-stimulating factor 1 from C. elegans. Using in situ hybridization, it could be demonstrated that cell expression of the gene for OSTF_SUBDO is highest in those cells surrounding the spicules and the sclerocytes. Our immunofluorescence studies with antibodies against silicatein and silicase experimentally support the earlier hypothesis of Uriz et al. (2003) to some extent by implying that spiculogenesis and spicule arrangement are under tuned genetic control by the synthesizing molecules, e.g., the different forms of silicatein, as well as hormone-like factors and/or transcription factor (homeodomain proteins).
Subsequently, the question of which structural molecules in sponges allow the formation of the species-specific diverse structures was approached by immunoelectron microscopy using antibodies against silicatein and galectin. Galectins, galactose-specific lectins, are dominant proteins existing in the mesohyl of the demosponges Geodia cydonium (Diehl-Seifert et al. 1985; Müller et al. 1997) and also S. domuncula (Wiens et al. 2003). In the mesohyl, “unstructured” extracellular matrix or ground substances classified to the carbohydrate-rich compounds (Garrone 1978) are also highly abundant. As in G. cydonium, the S. domuncula galectin forms three-dimensional clumps in the presence of Ca2+ (Diehl-Seifert et al. 1985; Wiens et al. 2003). To clarify if the biosilica-forming enzyme silicatein and the structural protein galectin are colocalized in the extraspicular space of the mesohyl, immunoelectron microscopic analyses were performed. This series of experiments showed that silicatein is associated with the galectin meshwork in concentric rings around the spicules. In a recent study (Schröder et al. 2006), we showed that spicule formation occurs in the final steps extracellularly in the mesohyl where an appositional growth of spicules can be demonstrated. Galectin molecules associate in the presence of Ca2+ to strings (nets) that allow binding of silicatein molecules (Schröder et al. 2006). Subsequently, collagen fibers seem to orient the silicatein-galectin strings concentrically around the growing spicules. Finally, as reported recently by Schröder et al. (2006), biosilica deposition is mediated in two directions, originating both from the silicatein-galectin strings and from the surface of the spicules (centripetal and centrifugal). Additional fibrous molecules might very likely be involved in the formation of the concentric rings as well, e.g., spongin (Wheeler and Mendel 1909), whose chemical composition and gene(s) are currently not defined (Simpson 1984), or collagen (Garrone 1978).
In the present study the spatial coexistence of the concentrically arranged galectin/silicatein superstructures with the collagen fibers is demonstrated. Because galectin is arranged in the mesohyl of sponge around the spicules in strings/bundles displaying no apparent organization pattern, it might be proposed that collagen serves as a major fibrillar network/matrix during the size- and form-specific mineralization of spicules (Simpson et al. 1985; Schröder et al. 2006). In the present study, organization of collagen fibrils in adult specimens was analyzed by SEM. It is documented that a collagen web surrounds the spicules in a highly organized manner. Along the longitudinal direction of the spicules the fibers are primarily arranged parallel to their surfaces. At the tips of the spicules, collagen fibers surround the knobs of the tylostyles as a mesh. It is not yet known if the slight swelling/knob of the tylostyles in S. domuncula is initially present during the synthesis of the spicules or secondarily apposed onto the spicules. In view of the available data (Schröder et al. 2006), the latter possibility is more realistic because oxea spicules, pointed at both ends, are seen almost exclusively in primmorphs. SEM analyses also show that a less fibrous material covers the intimate surfaces of the spicules, very likely the silicatein/galectin strings as identified by TEM. To the best of our knowledge, such a highly ordered organization of fibrils/fibers composed of galectin and collagen has not been previously reported in demosponges. This is in contrast to reports on the hexactinellid sponges, Euplectella marhalli (Ijima 1901) and Monorhaphis chuni (Schulze 1904).
Together the presented results show that during spiculogenesis the two enzymes silicatein (anabolism of biosilica) and silicase (dissolution) mediate biosilica deposition, a process that is matrix-guided primarily by galectin and secondarily also by collagen. Future experiments should address the question on the mechanism and specificity of interactions of these molecules and will try to elucidate the controlling genetic machinery switched on during spicule formation. The osteoclast-stimulating factor described here is a first step toward an understanding of the regulatory system controlling the size and shape of the spicules.
Footnotes
Acknowledgements
This work was supported by grants from the European Commission, the Deutsche Forschungsgemeinschaft, the Bundesministerium für Bildung und Forschung Germany, and the International Human Frontier Science Program.
We thank Ms. E. Sehn (Zoological Institute; University of Mainz, Mainz, Germany) for valuable technical assistance.