
Abstract
Caveolin-1, the major structural protein of caveolae, is present in several cell types known to play a role in the development of atherosclerosis. In this study, the distribution and expression of caveolin-1 in the arterial walls were studied in hypercholesterolemic rabbits. Immunohistochemical results indicated that the staining intensity of caveolin-1 reached a high level in the arterial intima at 5 weeks after high-cholesterol-diet treatment and decreased to a very low level at 8 weeks when atheromatous plaques appeared. Western blot analysis showed that in rabbits fed a high-cholesterol diet for 5 weeks, the expression of caveolin-1 reached its highest level and then decreased from 8 to 12 weeks. The proliferative activity of smooth muscle cells (SMCs) decreased to the lowest level at 5 weeks and then increased at 8 and 12 weeks. Nitric oxide synthase activity gradually decreased in animals fed a high-cholesterol diet throughout the experiment. These studies demonstrate that the change in abundance of caveolin-1 is associated with SMC proliferation in the formation of atheromatous plaque after hypercholesterolemia insult.
Keywords
C
Another important function of caveolin-1 is its ability to negatively regulate endothelial NOS (eNOS) activity (Feron et al. 2001). eNOS becomes hyperactivated in the absence of caveolin-1 (Bucci et al. 2000). Series experiments have demonstrated that Cav-1/caveolae regulate three major vascular functions dependent on NO and Ca2+ signaling: endothelium-dependent relaxation of arteries, myogenic tone, and stimulated contractility (Drab et al. 2001). Many studies have suggested that eNOS plays a major role in the development of atherosclerosis (Drexler 1999; Feron et al. 1999). It was reported that vascular cell adhesion molecule 1 (VCAM-1) was downregulated by e-NOS and NO (Takahashi et al. 1996; Huo and Ley 2001; Kawashima et al. 2001). NO was also shown to prevent SMC proliferation and migration in culture (Garg and Hassid 1989). Therefore, regulation of eNOS by caveolin-1 in endothelial cells may indirectly cause SMCs to develop into atherosclerosis. Recently, in vivo studies demonstrated that decreased expression of caveolin-1 was observed in dogs with hypertrophic cardiomyopathy (Piech et al. 2002). Dissociation of caveolin from caveolae is also associated with aging and myocardial infarction (Ratajczak et al. 2003). In this study, we established an experimental hypercholesterolemic rabbit model to analyze the distribution and expression of caveolin-1 in the arterial walls. In addition, we investigated whether alterations in caveolin-1 distribution and expression were related to SMC proliferation and eNOS dysfunction during hypercholesterolemia.
Materials and Methods
Animals and Diet
Forty New Zealand White rabbits weighing ~4kg were fed a 2% cholesterol diet for 12 weeks. A control group consisted of 40 rabbits fed a standard diet (Fu-Shou Co.; Taichung, Taiwan). Rabbits were weighed and total cholesterol, HDL-cholesterol, and LDL-cholesterol contents were determined weekly.
Analysis of Plasma Cholesterol Levels
Blood (1.5 ml) from rabbit ears was collected into Eppendorf vials and spun at 5000 × g for 10 min. Total plasma cholesterol and LDL cholesterol were measured spectrophotometrically using LDL-Cholesterin and Cholesterin Kits (Merck KgaA; Darmstadt, Germany).
Immunohistochemistry
Animals were anesthetized with a mixture of ketamine (40 mg/kg) and xylazine (5 mg/kg) given IM. After sacrificing, the carotid artery, aortic arch, thoracic aorta, and coronary artery were dissected and washed with PBS, dehydrated in graded alcohol, and embedded in paraffin. Tissues were cut into 5-μm sections, deparaffinized with xylene, and then rehydrated in a graded series of ethanol for hematoxylin and eosin staining and immunohistochemistry. Caveolin-1 and α-actin were detected by the streptavidin-biotin amplification method reported previously (Elias et al. 1989). In brief, tissue sections were washed in a running water bath for 10 min and then heated in 10 mM citric acid buffer (pH 6.0) for 15 min in a microwave oven (800 W) to enhance antigen retrieval. Endogenous peroxidase activity was quenched by treating the tissue sections with 3% hydrogen peroxide for 10 min. Nonspecific background was eliminated by incubating the tissue sections with non-immune serum. After washing with PBS, tissue sections were incubated with rabbit anti-human caveolin-1 antibody [1:200 (v/v); Santa Cruz Biotechnology, Santa Cruz, CA] and mouse anti-human α-actin antibody [1:200 (v/v); Lab Vision Corp., Fremont, CA] at 37C for 1 hr, followed by the addition of either biotinylated goat anti-rabbit IgG or biotinylated goat anti-mouse IgG. Tissue sections were then washed three times with PBS and incubated with streptavidin-peroxidase conjugate, followed by the addition of H2O2-DAB substrate-chromogen solution (Zymed Laboratories; South San Francisco, CA). Tissues were then counterstained with hematoxylin, dehydrated, mounted with Leica CV 5000 (Bartels and Stout Inc.; Issaquah, WA), and photographed with a Nikon Microphot-FXA light microscope (Tokyo, Japan). Immunostaining specificity was checked by treating control slides as described above except that the primary antibody was omitted.
Apoptosis Determination
Five-μ-thick tissue sections were deparaffinized with xylene, rehydrated in a graded series of ethanol, and washed with PBS solution. Tissue sections were incubated with proteinase K (20 μg/ml) at 37C for 15 min to expose the DNA for end labeling. For a positive control, tissue sections were exposed to 5 μg/ml DNase I in PBS at 37C for 60 min. After incubation, cell death was detected in tissue sections by a terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay kit (Roche Diagnostics; Meylan, France) according to the manufacturer's instructions. After washing with PBS followed by double staining with 1 μg/ml DAPI at room temperature for 20 min in the dark for nucleus identification, tissue sections were examined under a Nikon Eclipse E400 fluorescence microscope.
Protein Extractions
Thoracic aorta were washed with PBS and homogenized in lysis buffer (10 mM Tris-HCl, pH 7.4, 1 mM benzamidine, 150 mM NaCl, 0.5 mM EDTA, 10 mg/ml phenyl methane sulfonyl fluoride) at 4C overnight. Cells were resuspended in lysis buffer and sonicated by Sonicator 3000 (Misonix Inc.; Farmingdale, NY) on ice. Supernatant was harvested by centrifugation at 900 × g at 4C for 10 min. Protein concentrations were measured by a BCA protein assay kit (Pierce; Rockford, IL), and the samples were stored at −20C until needed.
Western Blot Analysis
Proteins (5 μg) were separated by 12% SDS-PAGE and electrotransfered to a polyvinylidene fluoride membrane. Blots were probed with rabbit anti-human caveolin-1 antibody [1:10,000 (v/v); Santa Cruz Biotechnology] followed by horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG [1:15,000 (v/v); Zymed Laboratories]. SMC proliferation was detected with mouse anti-rat proliferating cellular nuclear antigen (PCNA) antibody [1:10,000 (v/v); Upstate, Charlottesville, VA] followed by HRP-conjugated goat anti-mouse IgG [1:15,000 (v/v)]. After washing with PBS containing 0.5% Tween-20, peroxidase activity was visualized using enhanced chemiluminescence (PerkinElmer Life Science; Boston, MA). The same membrane was re-probed with a monoclonal antibody directed against β-actin [1:10,000 (v/v); Sigma, St Louis, MO] and goat anti-mouse immunoglobulin [1:10,000 (v/v); Zymed Laboratories) for an internal control. Intensity of reaction bands was analyzed by the Image Gauge System (Fuji; Tokyo, Japan).
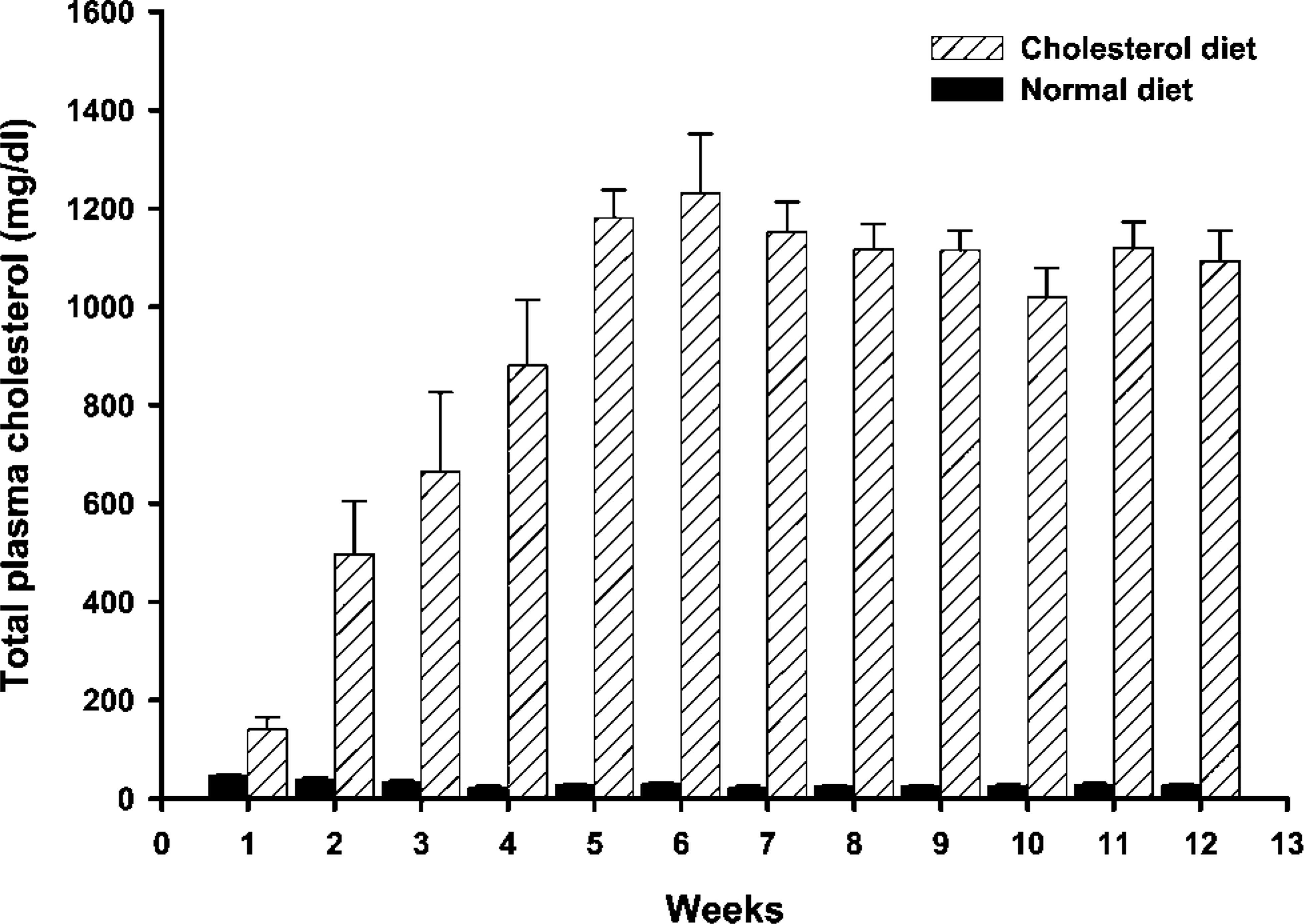
Changes in mean total plasma cholesterol levels in New Zealand rabbits fed a high-cholesterol diet and a normal diet. All values are mean ± SE,
Nitrite Production
Nitrite (NO2 -) production was measured by the Griess reaction in 5 × 10-mm specimens from the thoracic aorta (Uemura et al. 2000). Specimens were incubated in 1 ml Hanks' buffered saline solution (HBSS; Hyclone, Logan, UT) at 37C for 2 hr. Samples of the medium (80 μl) were collected, and nitrite was measured by the Griess reaction and a commercial colorimetric assay system (Promega; Madison, WI).
Results
Plasma Cholesterol Level
Total plasma cholesterol level in the controls remained in the normal range of 20-50 mg/dl. This level increased significantly from 1 to 12 weeks at all ages in high-cholesterol-fed rabbits (Figure 1).
Distribution of Caveolin-1 in the Arterial Wall
Morphological observations indicated that the small lesions appeared in the thoracic aorta of rabbits fed a high-cholesterol diet until 5 weeks into the experiments. In rabbits fed cholesterol for 7-8 weeks, atherosclerotic streaks were observed in the coronary arteries, aortic arch, thoracic aorta, and carotid arteries. Large, raised plaques consisting of foam cells, mononuclear cells, and SMCs had formed in some areas within 8-12 weeks of the high-cholesterol diet. Progression of the atherogenic lesions was consistent with the results of other studies (Ross 1986,1997,1999; Lupu et al. 1987). Endothelial denudation was not observed in the tissue sections. Immunohistochemistry detected caveolin-1 predominantly in the intima of arterial vessels in rabbits fed normal and high-cholesterol diets for 5 weeks (Figures 2A-2C). Only a small amount of caveolin-1 was observed in some SMCs after high-cholesterol-diet treatment (Figure 2B). Staining intensity was higher in the high-cholesterol diet animals than in the control animals. No reaction products were found in sections in which the primary antibody had been omitted (Figure 2D). As the plaques started to form, staining intensity of caveolin-1 in the non-lesion areas is more intense than in the lesion areas (Figures 3A-3D). More advanced plaque contains foam cells as well as large quantities of extracellular matrix in rabbits fed a highcholesterol diet for 12 weeks (Figures 4A and 4B). Many α-actin-positive cells indicated that the SMCs appeared in the plaques (Figure 4C). No reaction products were found in the sections with the primary antibody omitted (Figure 4D). Few to no TUNEL-positive cells were detected in the intima of atherogenic plaques in rabbits fed the high-cholesterol diet for 8 weeks (Figure 5A). A significant number of TUNEL-positive cells was observed in the DNAase-treated tissue section (Figure 5B). These results demonstrated that most cells in the atherogenic plaques had not undergone apoptosis. DAPI staining shows the appearance of nucleus (Figures 5C and 5D).
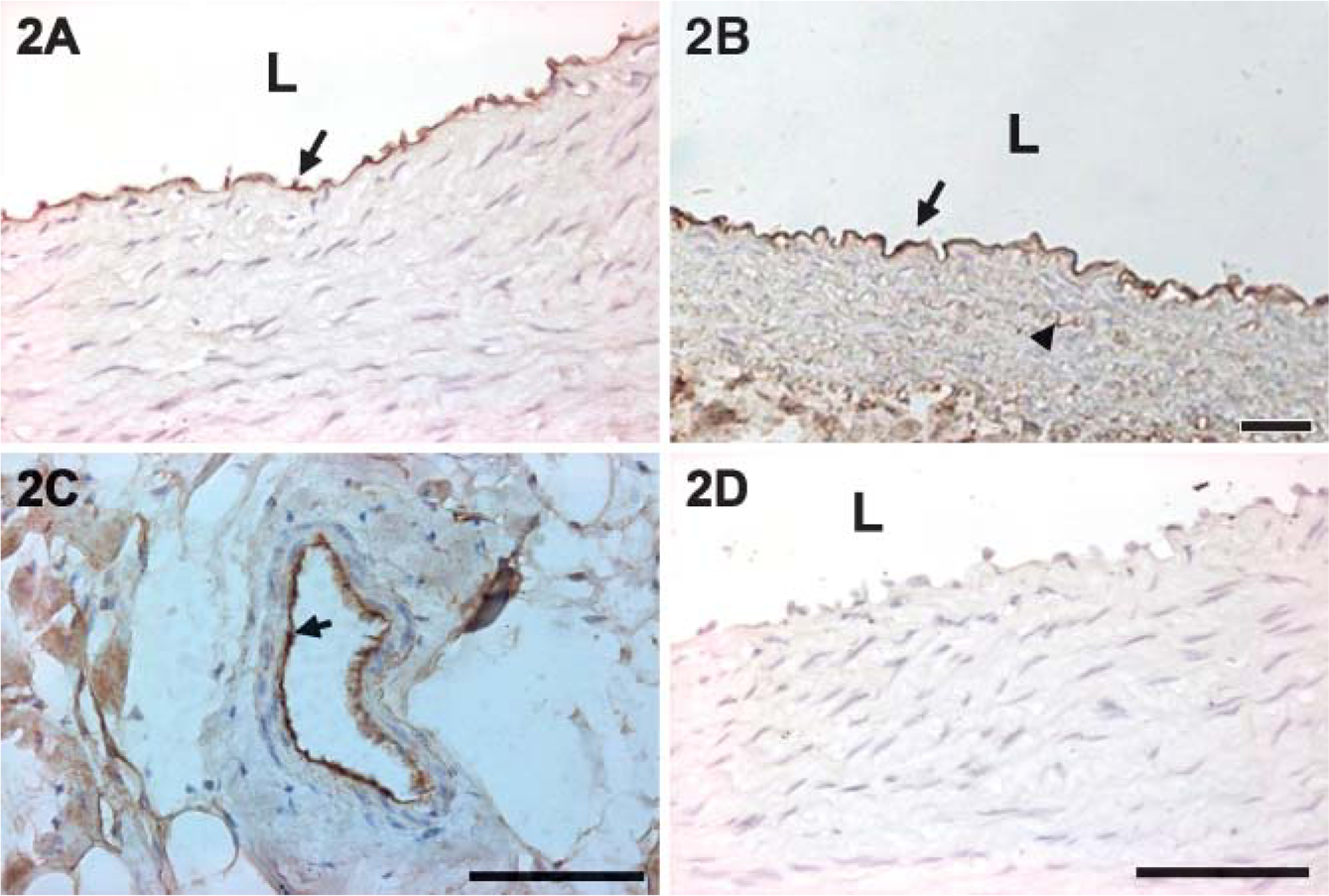
Caveolin-1 distribution in aortic arch after 5 weeks on the normal diet (
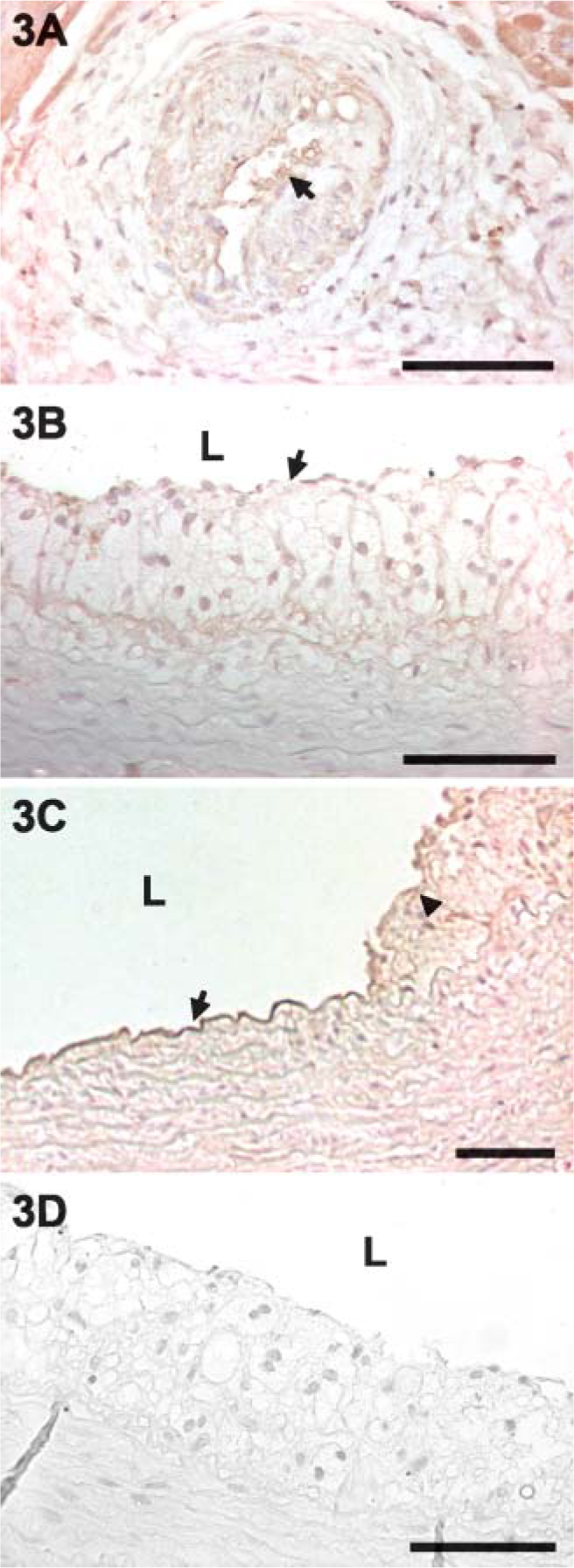
Staining intensity of caveolin-1 decreased in the plaque areas (arrows) of coronary artery (
Expression of Caveolin-1 and PCNA in the Arterial Wall
Western blot analysis showed that the level of caveolin-1 increased gradually in the vessel walls of control rabbits and reached its highest concentration at 12 weeks into the experiment (Figure 6A). It also showed that the expression of caveolin-1 reached its highest level at 5 weeks and then decreased from 8 to 12 weeks in the rabbits fed a high-cholesterol diet (Figures 6B and 6C). PCNA levels decreased in the animals fed a high-cholesterol diet at 5 weeks and then increased at 8 and 12 weeks (Figures 6B and 6D).
Nitrite Production in the Arterial Wall
Nitrite production in the arterial walls gradually decreased in cholesterol-diet groups throughout the experiment. Nitrite level was significantly lower in animals fed the high-cholesterol diet after 5 and 8 weeks than the normal controls (Figure 7).
Discussion
To the best of our knowledge, this is the first in vivo study on the distribution and expression of caveolin-1 in the arterial walls of diet-induced hypercholesterolemic rabbits. The present study demonstrated that caveolin-1 increased and PCNA decreased in the early stages (5 weeks) of ingesting a high-cholesterol diet. At 8 and 12 weeks, the level of caveolin-1 decreased while the PCNA increased. Meantime, appearance of atherogenic plaques became more severe at 8 and 12 weeks. Nitrite production gradually decreased in the cholesterol feeding groups.
Hypercholesterolemia is a crucial factor of atherosclerosis, a process histologically characterized by lesions progressing from fatty streaks to fibrous plaques, ultimately occluding the lumen of the affected artery (Ross 1993). Several studies have suggested that caveolin-1 and caveolae may play a proatherogenic role by promoting the transcytosis of LDL particles from the blood to the subendothelial space (Palade 1953; Fielding and Fielding 1996). It has been demonstrated that dietinduced hypercholesterolemia may increase the permeability of endothelium, allowing LDLs to infiltrate and accumulate in the intima (Lee et al. 2001). Furthermore, one study reported that the plasmalemmal caveolae and the open junctions increased significantly in the branched regions of arteries in hypercholesterolemic rats (Kao et al. 1995). In this study, the concentration of plasma LDL cholesterol increased from 140 mg/dl to 1250 mg/dl between 2 and 5 weeks after high-cholesterol-diet treatment. The fatty streak did not start to appear in the intima until week 5. Concomitantly, the level of caveolin-1 increased and reached its highest level at about 5-6 weeks; at that time, the level of caveolin-1 began to decrease. It has been reported that caveolin-1 is more abundant in endothelial cells than in SMCs (Frank and Lisanti 2004). Our immunohistochemical results also showed that caveolin-1 is expressed predominantly in the endothelial cells. We felt that the increased caveolin-1 shown in the result of Western blotting was mainly due to the endothelium.
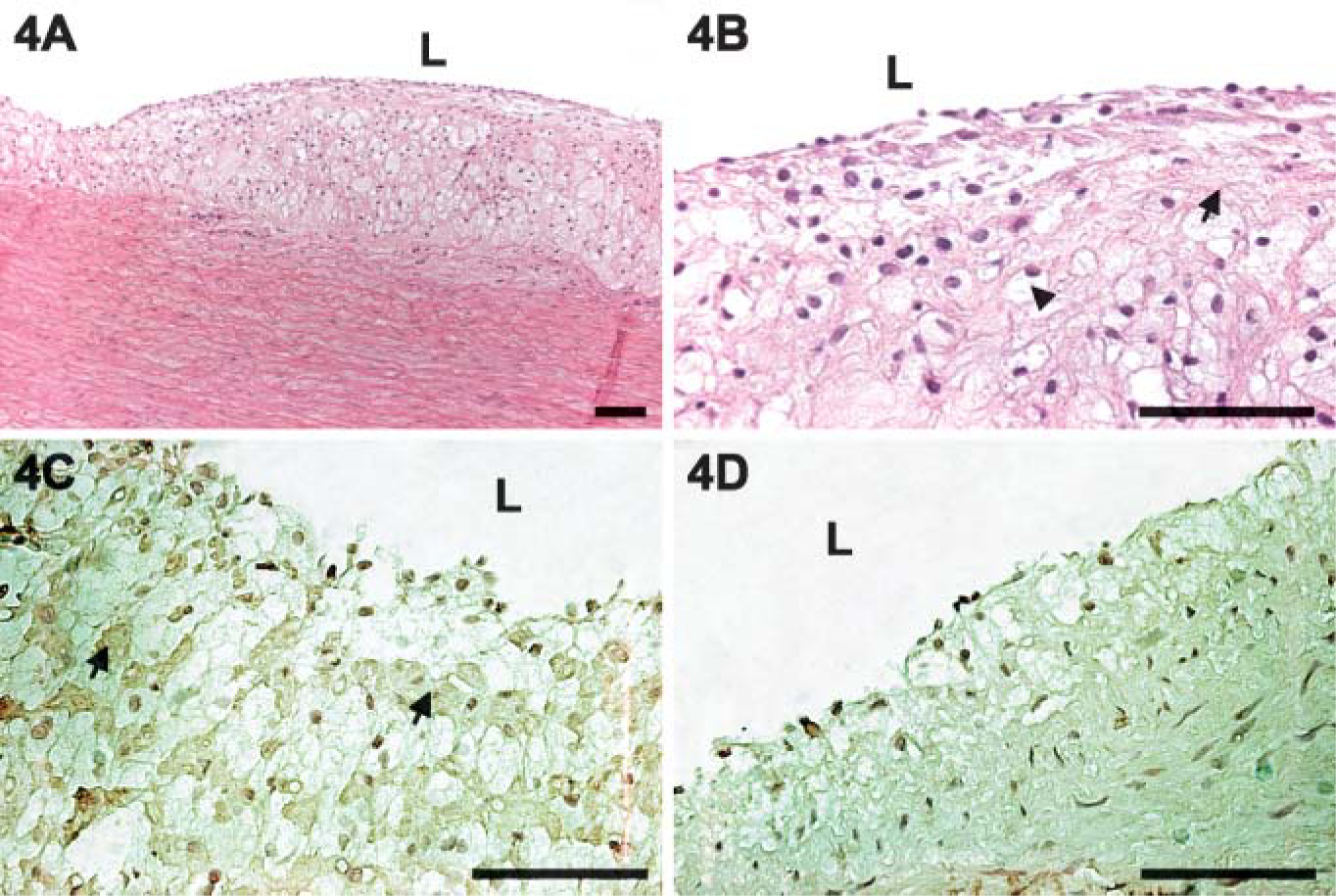
Thoracic aorta of a rabbit after 12 weeks on the high-cholesterol diet (
Several in vitro studies have shown that overexpression of caveolin-1 enhanced the cholesterol efflux in the culture system. Free cholesterol and oxysterols could modify free cholesterol efflux by means of opposite effects on the expression of caveolin (Lisanti et al. 1994; Fielding et al. 2002). In confluent fibroblast monolayers, caveolin mRNA levels are upregulated when increased levels of cholesterol enter the cell from LDL. The upregulation of caveolin-1 expression may represent a protective mechanism against cellular cholesterol accumulation in these cells. On the contrary, oxysterols reduced caveolin mRNA levels, as well as transport of cholesterol to the cell surface and cholesterol efflux (Fielding et al. 1997). Oxysterols are formed from free cholesterol when it combines with oxygen. 7-Ketocholesterol and 7-α-hydroxy cholesterol are major components of human atherosclerotic lesions (Fielding et al. 1997). Accumulation of oxidized (ox) LDL in the intima after long periods of feeding is thought to be involved in the development of atherosclerosis. Among the multiple cell types in the vascular system, the accumulation of oxLDL was chemotactic to monocytes and SMCs. We assume from our results that, in the early stage of hypercholesterolemia, caveolin-1 synthesis increased to remove the excess cholesterol from the cells. However, after long periods of feeding, caveolin-1 synthesis was repressed. It has been reported that reactive oxygen species (ROS) are profound activators of signal transduction events that may alter eNOS phosphorylation, resulting in the dysfunction of endothelial function. Additional work will be needed to clarify the role of long-term hypercholesterolemia in the regulation of caveolin-1 expression.
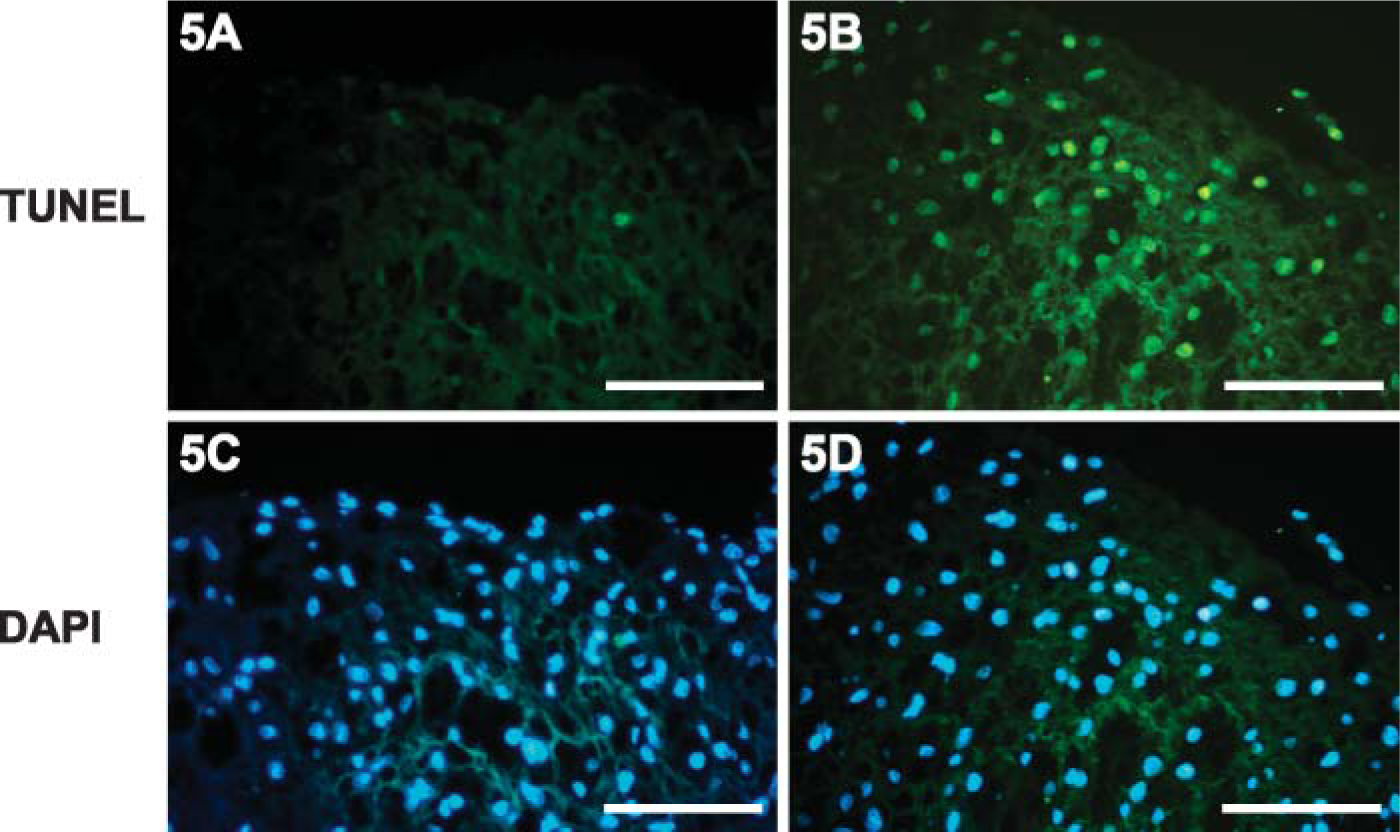
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay and DAPI staining were performed. Few to no TUNEL-positive cells were detected in the lesion area of aortic intima (
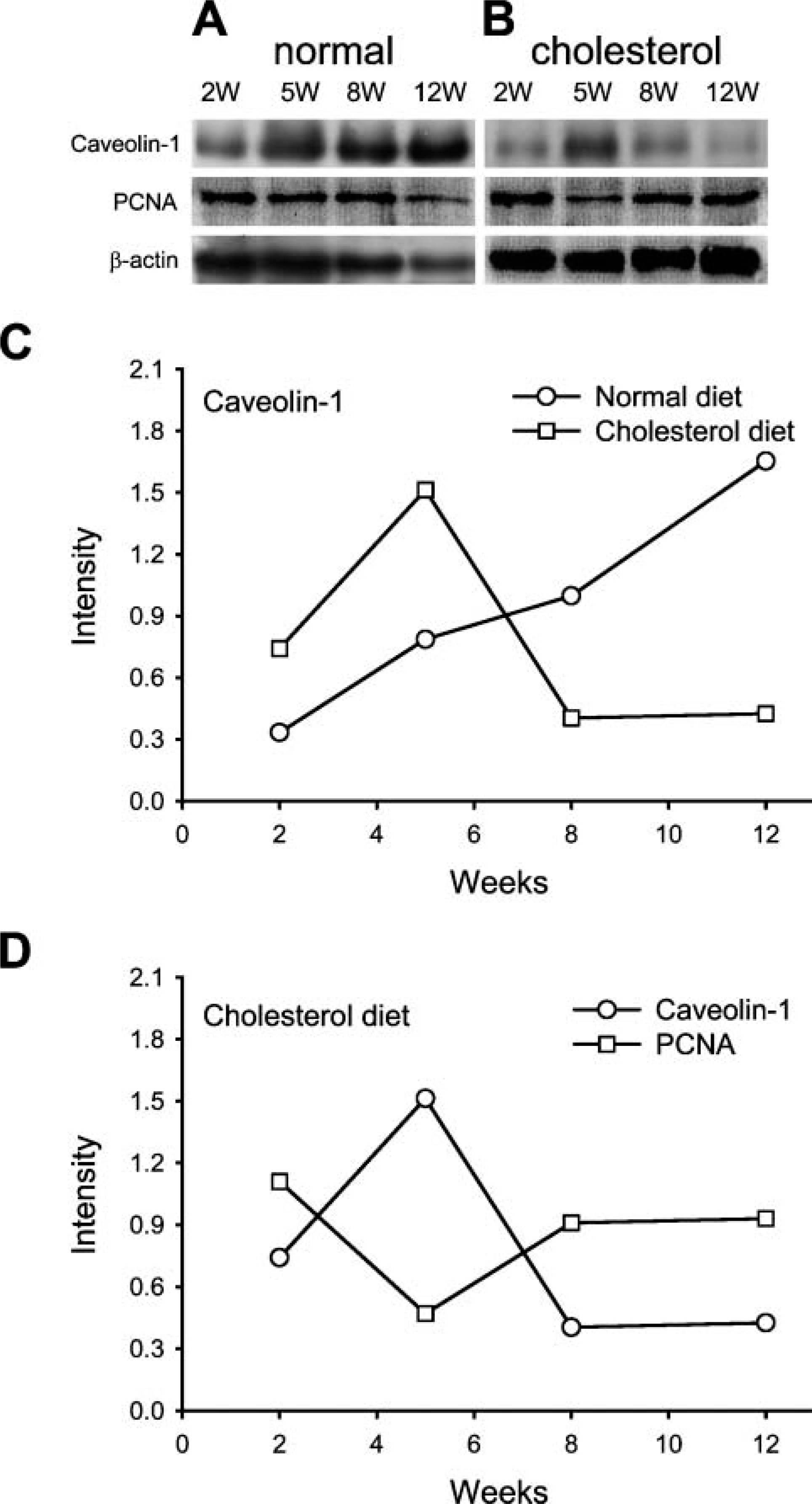
Abundance of caveolin-1 in thoracic aorta of rabbits after 2-12 weeks on the normal diet and high-cholesterol diet. Caveolin-1 level increased gradually and reached its highest level at 12 weeks in the rabbits on the normal diet (
A recent study by Zulli et al. (2006) indicated that the expression of caveolin-1 in the endothelium-over-lying plaques is increased. They used a combination diet (0.5% high-cholesterol diet plus 1% methionine) to induce hyperhomocysteinemia and hypercholesterolemia in the rabbit for 12 weeks. On the contrary, we used 2% high-cholesterol diet to induce hypercholesterolemia. The morphology of plaques in Zulli's paper was different from what we observed in our experiment. For example, we found foam cells and extracellular matrix in the plaques. Results similar to ours have been reported in previous studies (Ross 1986,1993,1997,1999). We assume that the different experimental design may have caused the different results.
Endothelium-derived NO modulates vasodilation and prevents platelet adhesion and SMC proliferation. NO has also been shown to have antioxidant effects by enhancing the expression of superoxide dismutase, preventing lipid-chain reactions, and reacting with the superoxide anion. Thus, NO seems to play a major role as an endogenous protective factor against atherosclerosis. Caveolin-1 in endothelial cells has been reported to negatively regulate eNOS activity (Razani et al. 2001). For example, NOS became hyperactivated in the absence ofcaveolin-1 (Shauletal. 1996). The ability of caveolin-1 to inhibit eNOS might also contribute to the regulation of vascular permeability (Fukumura et al. 2001). In this study, NOS activity decreased gradually in high-cholesterol diet groups. In other words, after a long period of ingesting a high-cholesterol diet, both the level of caveolin-1 and the activity of NOS decreased. It has been reported that dietary cholesterol reduced endothelium-dependent relaxations and decreased NO production (Boger et al. 1995). Enhanced superoxide anion (O2 -) production may further contribute to the decreased biological activity of NO in hypercholesterolemia. For example, ROS were shown to inhibit cholesterol-mediated increases in caveolae number in bovine aortic endothelial cells (Peterson et al. 2003).
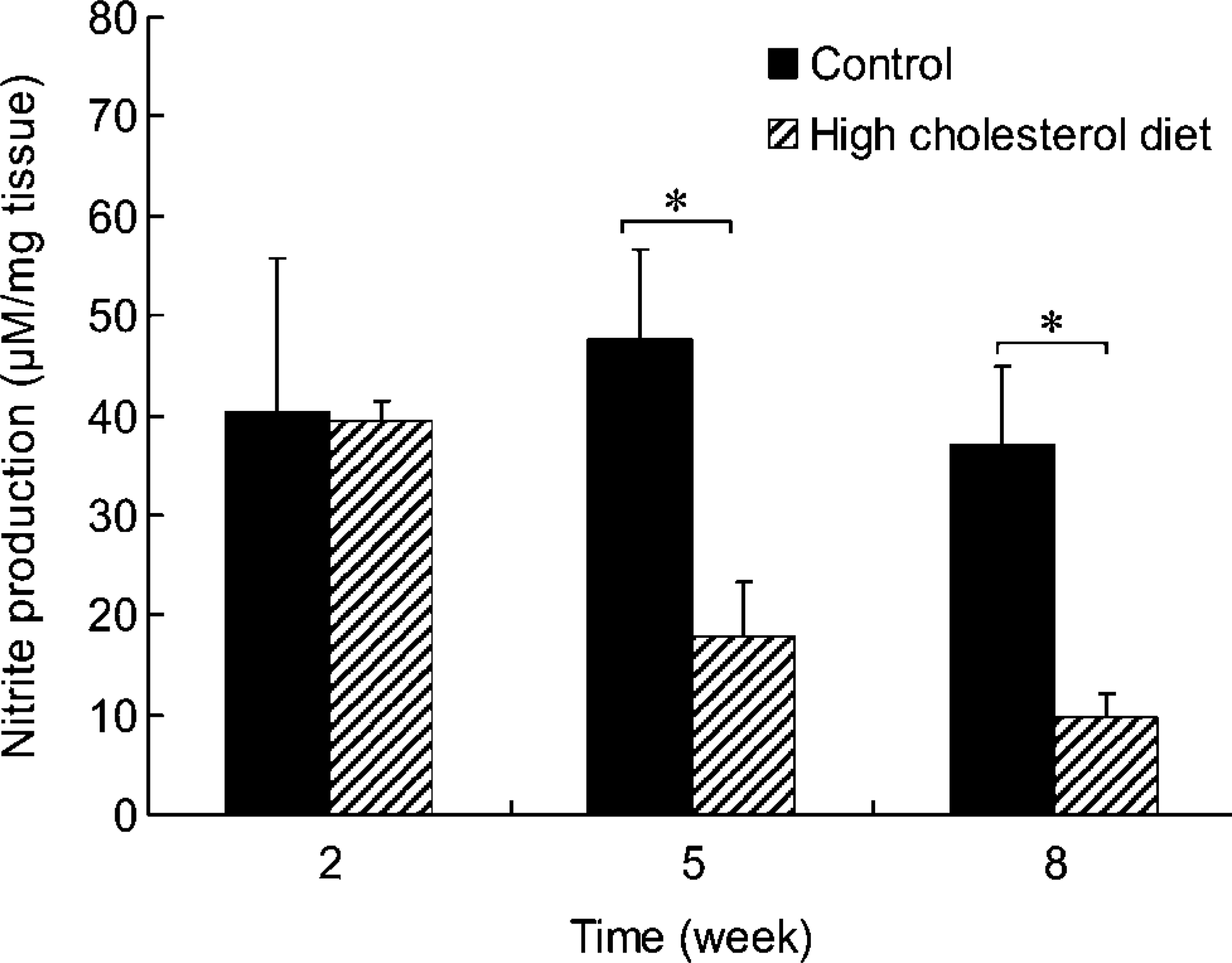
Nitrite production in the segments of thoracic aorta from rabbits fed normal and high-cholesterol diets for 8 weeks. Nitrite production gradually decreased in tissues from animals fed a high-cholesterol diet. Lower production of nitrite was observed in tissues from animals fed a high-cholesterol diet than in tissues from the control animals at 5 and 8 weeks.
SMC proliferation is one of the important events in the development of atherosclerosis. The regulation of eNOS by caveolin-1 in endothelial cells may indirectly affect neighboring SMCs in the arterial walls that are prone to the development of atherosclerosis. One study has reported that genetic ablation of caveolin-1 in mice stimulates the proliferation of SMCs (Hassan et al. 2004). It has also been shown that the transition of SMCs from a contractile to a synthetic phenotype is associated with a loss of plasma membrane caveolae. Several reports have proposed an antiproliferative and possibly antimigratory role for caveolin-1 (Peterson et al. 2003). In this study, we demonstrated that the decreasing level of caveolin-1 was associated with increasing SMC proliferation in the arterial walls of hypercholesterolemic rabbits. This finding provides new information about the role caveolin-1 plays in atherosclerotic formation in vivo. Regulation of caveolin-1 expression may be important in the prevention and treatment of atherosclerosis.
Footnotes
Acknowledgements
This work was supported by research grant TCVGHT-T-927802 from the Taichung Veterans General Hospital and research grant THU 2208-74-62 from the Tunghai University, Taiwan, Republic of China.
The technical assistance of S.H. Tsai is gratefully acknowledged.