
Abstract
Localization and coordinate phosphorylation/dephosphorylation of endothelial nitric oxide synthase (eNOS) are critical determinants for the basal and stimulated production of nitric oxide. Several phosphorylation sites in eNOS have been identified as targets of the cAMP-dependent protein kinase A (PKA). Basal eNOS activity is also regulated by interaction with caveolin-1, the major coat protein of caveolae. In the present study we have examined in rat aorta endothelium the subcellular steady-state distribution of eNOS, the catalytic subunit of PKA (PKA-c), and caveolin-1. Basal eNOS expression was found in two distinct locations, the endothelial cell surface and the Golgi complex. Cell surface eNOS was equally distributed over caveolar and non-caveolar membranes but was 2.5-fold enriched on luminal lamellipodia located at endothelial cell contacts. PKA-c colocalized with eNOS in the lamellipodia, whereas caveolin-1 was absent from these membrane domains. PKA-c was also found associated with cell surface caveolae and with tubulovesicular membranes of Golgi complex and endosomes. The topological proximity of eNOS with the catalytic subunit of PKA in restricted intracellular locations may provide mechanisms for differential PKA-mediated eNOS regulation.
Keywords
N
We here present data on the subcellular steady-state distribution of eNOS in relation to two potential regulatory proteins, caveolin-1 and the catalytic subunit of PKA (PKA-c), in rat aorta endothelium. We demonstrate that an important pool of cell surface eNOS is outside caveolae and principally located in endothelial cell junctions, where it is closely associated with PKA-c, and not with caveolin-1. Distinct pools of PKA-c were also associated with cell surface caveolae and with the trans site of the Golgi. These restricted sites of colocalization may provide mechanisms for PKA-dependent eNOS regulation in different endothelial subcellular compartments.
Materials and Methods
Cells and Antibodies
Primary bovine aortic endothelial cells (BAECs) were obtained from Clonetics (Walkersville, MD). Monoclonal anti-eNOS (#N300200), monoclonal anti-caveolin-1 (#C37120), and polyclonal anti-caveolin-1 (#13630) were from Transduction Laboratories (San Diego, CA). Rabbit polyclonal anti-eNOS antibody, directed to the COOH-terminal domain of the molecule, was raised in rabbits. Monoclonal anti-phospho eNOS Ser 1177 (#9571) was from Cell Signaling (Beverly, MA). In BAECs, the antibody reacted with eNOS only when phosphorylated at Ser1177. Rabbit polyclonal antibody directed to the catalytic subunit of PKA-c was from Santa Cruz Technology (Santa Cruz, CA; sc-903). On immunoblots of human platelet lysates, the antibody reacted with the three subunits, α, β, and γ, of PKA-c (den Dekker et al. 2002). Fluorescent conjugated secondary antibodies were from Jackson Immunochemicals (West Grove, PA).
Cell Culture and Tissue Preparation
The procedures involving the use of animals followed the Institute of Laboratory Animal Resources (1996) Guide for the Care and Use of Laboratory Animals. Rats were anesthetized and perfusion-fixed through the left ventricle of the heart with a mixture of 2% paraformaldehyde (PFA) and 0.2% glutaraldehyde (GA) in 0.1 M sodium phosphate buffer (pH 7.4). After perfusion, small aorta rings were excised, postfixed for 2 hr at room temperature, and stored in 1% PFA at 4C. After washing, the samples were embedded in 10% gelatin, cooled in ice, and cut into 1-mm3 blocks in the cold room. The blocks were infused with 2.3 M sucrose at 4C for 24 hr, frozen in liquid nitrogen, and stored until cryoultramicrotomy. Primary BAECs were grown in EGM BulletKit medium (Clonetics) according to the manufacturer's instructions, fixed in 2% PFA and 0.2% GA in 0.1 M sodium phosphate buffer (pH 7.4), infused with 2.3 M sucrose at 4C, and frozen in liquid nitrogen.
Immunofluorescence
Semithin cryosections were prepared from the luminal area of the aorta. The sections were transferred to microscope slides and incubated with primary antibodies for 30 min, followed by incubation with Cy3-conjugated secondary antibodies. In a similar fashion, BAECs were permeabilized, and immunostained for 60 min with primary antibodies, washed three times with PBS, followed by a 60-min incubation with Cy3-conjugated secondary antibodies. Samples were analyzed by a Leica TCS 4D confocal laser-scanning microscope (Leica; Lasertechnik, Heidelberg, Germany).
Immunoelectron Microscopy
Cryosections 50 nm thick were cut at −120C using an Ultracut S ultramicrotome (Leica). The sections were collected on formvar-coated grids using a mixture of 1.8% methylcellulose and 2.3 M sucrose (Liou et al. 1996) and incubated with primary antibodies and protein A–gold (Slot et al. 1991). For monoclonal antibodies (MAbs), rabbit anti-mouse was used as bridging antibody. Immunogold double labeling was performed using 10-nm (eNOS) and 15-nm (Cav-1 and PKA-c) gold particles (Slot and Geuze 1985). After labeling, the sections were fixed with 1% glutaraldehyde, counterstained with uranyl acetate, and embedded in methyl cellulose-uranyl acetate. The sections were viewed in a JEOL 1200CX electron microscope.
Quantitative Analysis
For quantitative evaluation, cryosections were prepared from three different areas of the aorta lumen and 25 electron micrographs were taken randomly from the endothelium. The endothelial cell surface was subdivided into luminal, basal, and lateral membranes and the peripheral junctional membranes (lamellipodia). Gold particles over these categories were attributed to caveolar and non-caveolar membranes. The relative membrane length was determined using a transparent overlay with lines on the electron micrographs and counting the number of membrane intersections over the categories. The linear labeling density, defined as the ratio of gold particles and membrane intersections, was expressed per μm membrane. The intracellular gold label distribution of eNOS was determined over endoplasmic reticulum, Golgi complex, non-identified membranes, Weibel-Palade bodies, and cytosol. eNOS surface density was defined as the ratio of gold particles and random points over the compartments, and expressed per μm2. Random points were counted using a transparent overlay with points. The relative distribution of PKA-c was determined over caveolar and non-caveolar plasma membrane, peripheral cytoskeletal network, and intracellular membranes. Gold particles within 20 nm of a membrane were considered membrane-associated. Statistical ANOVA of linear label density was performed using the Student's t-test.
Results
Immunofluorescence (IF) of semithin cryosections of rat aorta endothelium showed a focal eNOS staining at the luminal site of the endothelial cell layer (Figure 1A). Caveolin-1 staining was peripheral, consistent with localization on basal and luminal surfaces of the endothelial cells (Figure 1B). The labeling pattern, however, was not uniform and showed distinct areas where caveolin-1 was absent. This typical staining pattern suggested that caveolin-1 was particularly absent in areas where endothelial cells made contact. IF staining of PKA-c appeared more diverse, showing a distinct focal distribution pattern often located at the endothelial cell borders (Figure 1C).
Immunoelectron microscopic analysis of immunogold-labeled cryosections revealed that as much as 48 ± 3.3% of total eNOS was found at the cell surface, whereas the remaining 52 ± 3.3% was intracellularly located. A fraction of the enzyme was located in luminally and basally oriented caveolae, which made up ~40% of the total endothelial cell surface. A substantial proportion of cell surface eNOS was found outside caveolae and was located in lateral endothelial contacts, particularly associated with peripheral membranes (lamellipodia) that overlaid adjacent endothelial cells (Figure 2A). Because these endothelial cell contacts are made up of a 2-fold membrane density, we quantified the label density (defined as gold particles per μm membrane) over the peripheral lamellipodia and other plasma membrane categories (Table 1). eNOS was 2.5-fold enriched on the lamellipodia compared with the average plasma membrane (p<0.005, Student's t-test) and was equally distributed over caveolar and non-caveolar membranes. Consistent with studies in cultured cells, we found a dominant pool of eNOS in the Golgi complex (Figure 2B; Table 1). We found no qualitative differences in the eNOS localization using two independent anti-eNOS COOH-terminal antibodies (not shown).
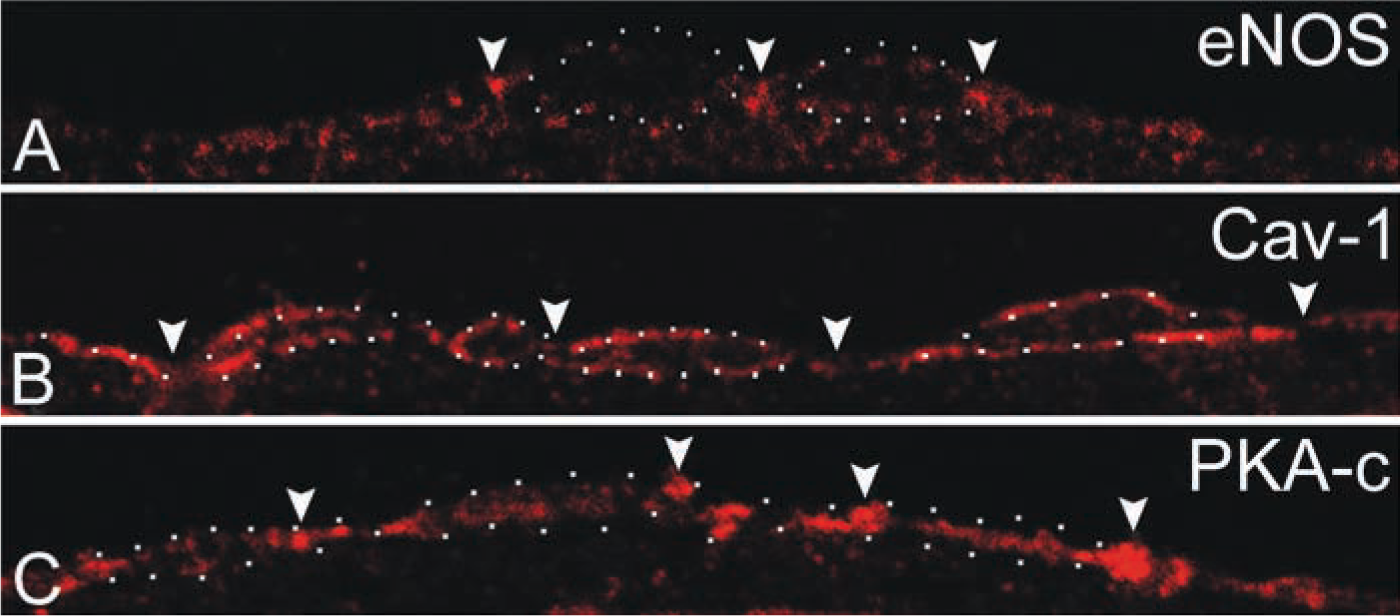
Immunofluorescence staining of eNOS, caveolin-1, and PKA in rat aorta endothelium. Semithin cryosections of rat aorta endothelium were labeled with antibodies directed to eNOS, caveolin-1, and PKA-c. (
On the basis of these observations, we conclude that at steady state, eNOS is present in three distinct locations, (a) a caveolae-associated pool, (b) a Golgi-associated pool, and (c) a non-caveolar plasma membrane pool enriched in junctional lamellipodia. Because caveolin-1 is a negative regulator of eNOS and several PKA target sites have been identified in eNOS that regulate its activity, we next analyzed the subcellular distribution of caveolin-1 and the catalytic sub-unit of PKA. Caveolin-1 was abundantly associated with luminally and basally oriented caveolae but was completely absent from the lamellipodia at endothelial contacts (Figure 3). A majority of PKA-c was associated with peripheral actin fibrils (Figure 4A), representing the previously reported apical and basolateral focal adhesions (White and Fujiwara 1986). A conspicuous labeling of PKA-c was found at cell–cell contacts and was particularly associated with the junctional lamellipodia where eNOS was enriched (Figure 4A). Specific labeling of PKA-c was also found within cell surface caveolae (Figure 4B) and within non-caveolar plasma membrane (Table 2). Immunogold double labeling confirmed the close proximity of eNOS and PKA-c in the peripheral lamellipodia and in lateral cell–cell contacts (Figure 5). Distinct intracellular pools of PKA-c were also detected in the Golgi area and in close proximity to tubulovesicular endosomes (Figure 4B). The specific localization of PKA-c in the Golgi area was also confirmed in cultured BAECs, where IF staining was typically found in the perinuclear region (not shown) and immunogold labeling specifically associated with tubulovesicular membranes at the trans site of the Golgi complex (Figure 6A). This small but distinct pool of PKA-c associated with the Golgi complex is of particular interest because both BAECs and rat aorta endothelium possess a large pool of eNOS in the Golgi, and it has been suggested that PKA-dependent phosphorylation of eNOS in the Golgi leads to the local release of NO (Fulton et al. 2002). Indeed, using an anti-phosphorylated eNOS Ser-1177 MAb, we found low but specific labeling in the Golgi area (Figure 6B).
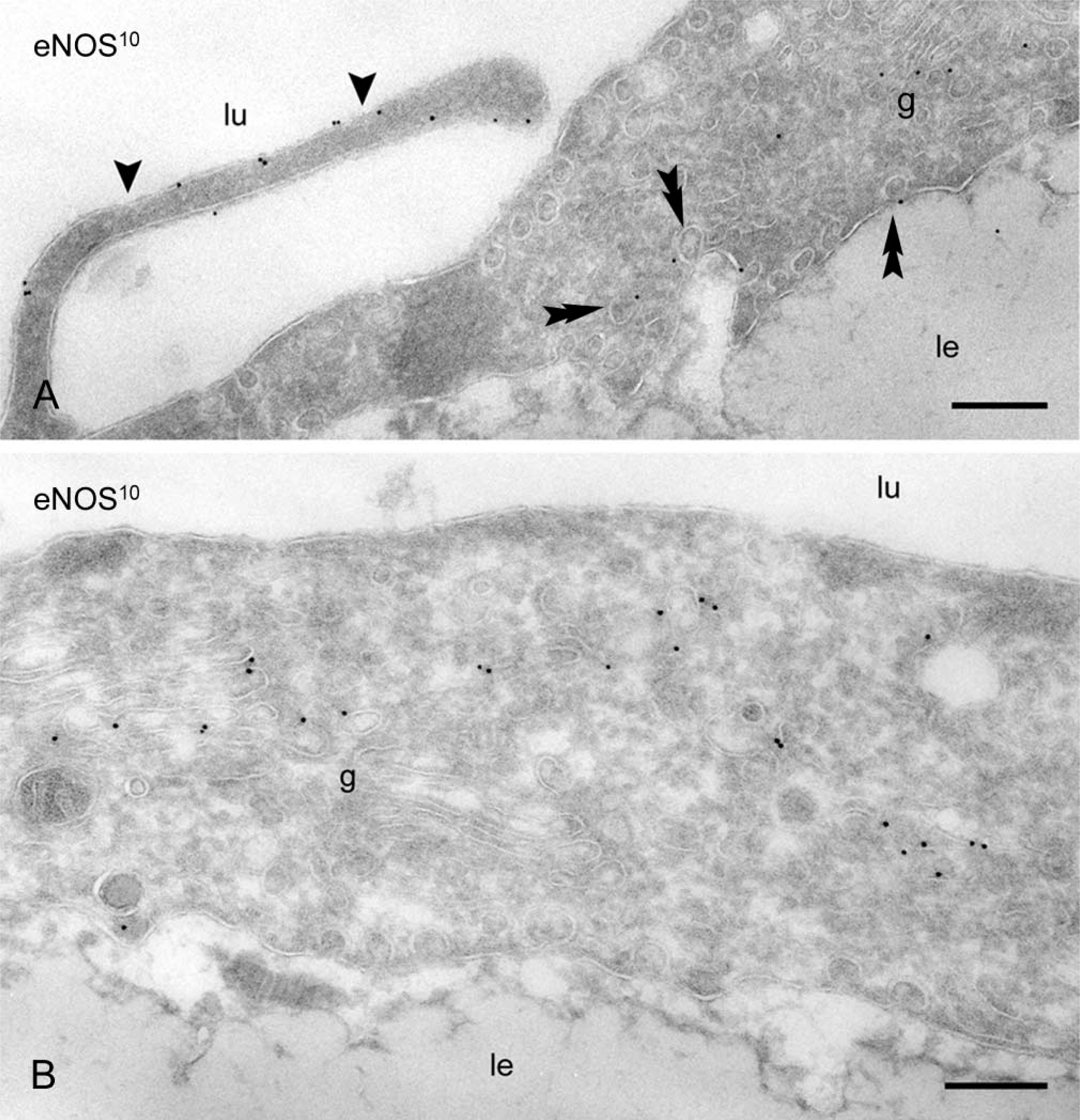
Immunoelectron microscopic localization of eNOS in rat aorta endothelium. Thin frozen sections of rat aorta endothelium. (
Discussion
Thus far, only limited morphological approaches have been applied to study the steady-state distribution of eNOS in vivo in endothelium. On the basis of biochemical fractionation data and immunohistochemistry of subconfluent cultured cells, it has been proposed that eNOS activity is mainly associated with cell surface caveolae. We here present data on the subcellular distribution of eNOS in relation to two regulatory proteins, caveolin-1 and PKA-c, in rat aorta endothelium. We found that at steady state, eNOS is present in three distinct locations, (a) cell surface caveolae, (b) Golgi complex, and (c) a pool associated with endothelial cell junctions. The latter pool was 2.5-fold enriched over caveolar and non-caveolar membranes, and thus represents a dominant pool of cell surface eNOS that is located outside caveolae and that is typically associated with lamellipodia that make up the endothelial cell junctions.
Subcellular distribution of eNOS in rat aorta endothelium a
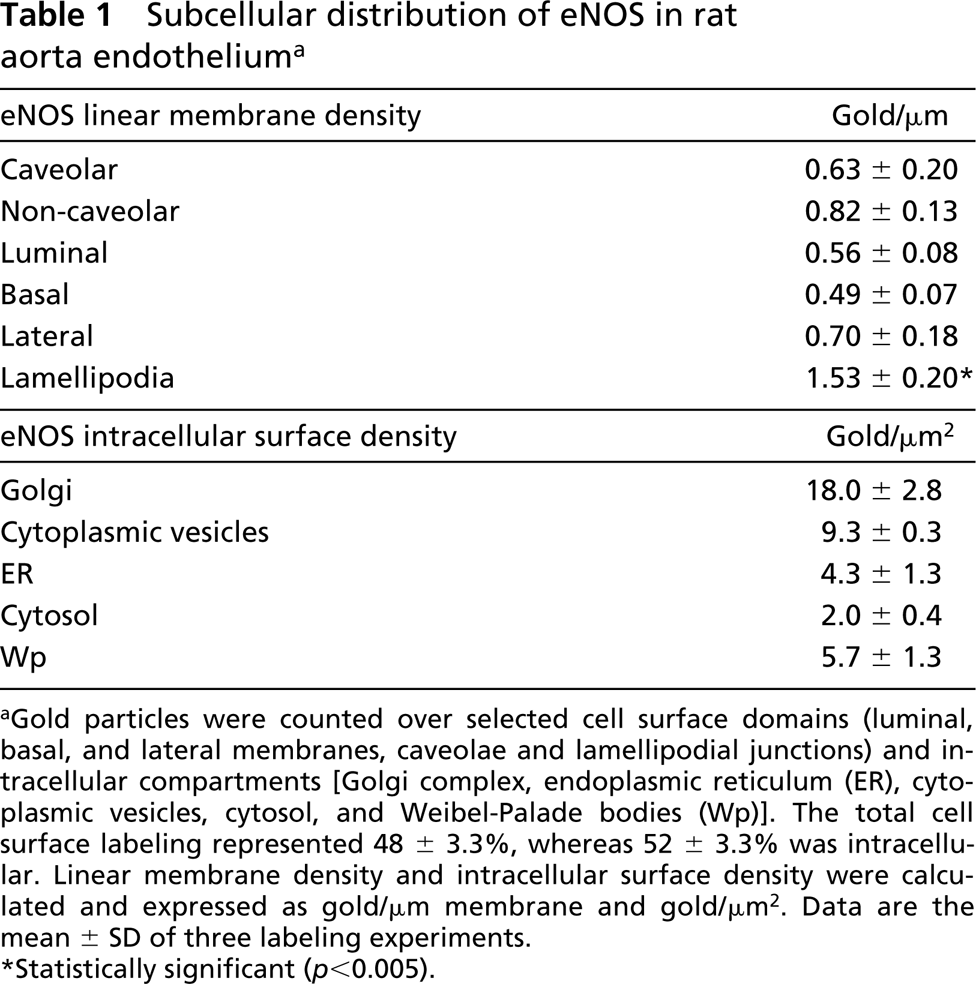
aGold particles were counted over selected cell surface domains (luminal, basal, and lateral membranes, caveolae and lamellipodial junctions) and intracellular compartments [Golgi complex, endoplasmic reticulum (ER), cytoplasmic vesicles, cytosol, and Weibel-Palade bodies (Wp)]. The total cell surface labeling represented 48 ± 3.3%, whereas 52 ± 3.3% was intracellular. Linear membrane density and intracellular surface density were calculated and expressed as gold/μm membrane and gold/μm2. Data are the mean ± SD of three labeling experiments.
∗Statistically significant (p<0.005).
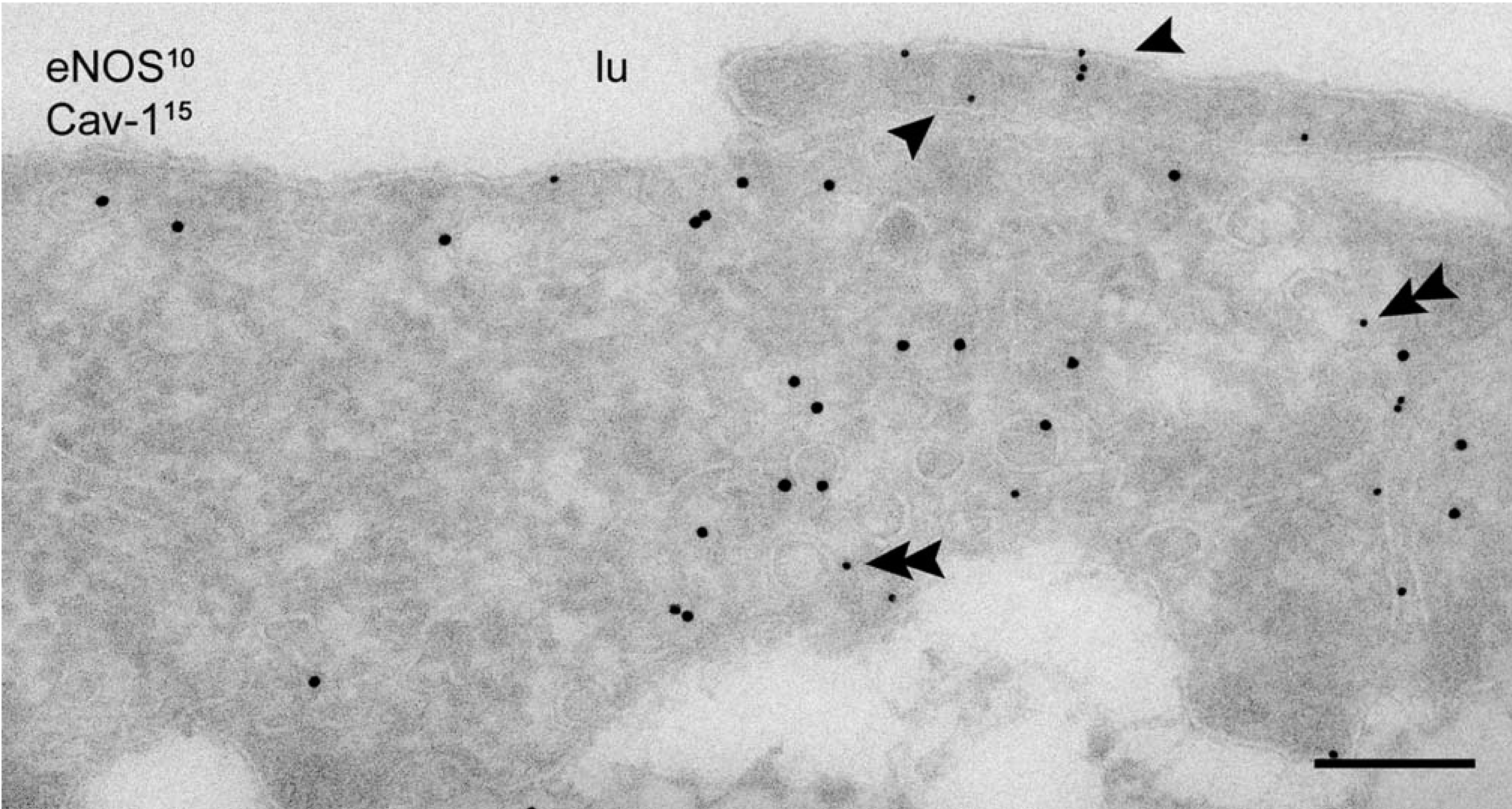
Codistribution of eNOS and caveolin-1 in rat aorta endothelium. Caveolin-1 localization in basal and luminally oriented caveolae of the endothelium. Peripheral lamellipodia at endothelial cell borders are completely devoid of caveolin-1 but show preferential localization of eNOS (arrowheads). Double arrowheads indicate eNOS associated with caveolae. lu, lumen of the aorta. Bar = 200 nm.
There is increasing evidence that eNOS activity is regulated by PKA-mediated phosphorylation (Michell et al. 2001,2002; Boo et al. 2002a, b). Several PKA target sites have been identified in bovine eNOS (Ser 1179, Ser 635, and Ser 617). Selectivity of PKA signaling is mainly provided by its subcellular location. Targeting of PKA catalytic activity to restricted intracellular locations depends on the binding of the PKA-RII subunit to the so-called A-kinase anchoring proteins (Edwards and Scott 2000; Skalhegg and Tasken 2000). The diverse locations of PKA-c that we found in our present study are consistent with the concept that PKA-c action is compartmentalized in restricted areas in the aortic endothelial cell, suggesting that different PKA pools may be operational in the regulation of eNOS. eNOS and PKA-c are in close proximity in the peripheral lamellipodia, at endothelial cell–cell contacts, and in the Golgi area. The 2.5-fold enrichment of eNOS in the lamellipodial membranes and the absence of caveolin-1 therein indicate that the local eNOS activity in these peripheral contact areas is not regulated by caveolin-1 interaction. The topological proximity of PKA-c and eNOS instead highlights a possible PKA-dependent regulation of eNOS activity in these locations. Our finding that a proportion of PKA-c is associated with caveolae is not quite unexpected. In vitro, caveolin-1 can interact with the catalytic subunit of PKA and negatively regulate PKA-c action (Razani et al. 1999; Razani and Lisanti 2001). This may have implications for the shear-dependent eNOS activation in these domains. Luminal caveolae are believed to be the mechanical sensors of blood flow variations (Rizzo et al, 1998). Shear-induced alterations in the interaction of caveolin-1 with PKA-c may affect the phosphorylation state of eNOS in caveolae and have downstream effects on the eNOS activity in these domains. Support for such a mechanism is obtained from recent studies showing that shear-induced phosphorylation of eNOS at Ser 1179 and Ser 635 is fully PKA-dependent (Boo et al. 2002a, b). However, the peripheral lamellipodia are also luminally oriented, and changes in pulsatile blood flow may therefore be sensed in these membrane domains as well. Because eNOS activity and PKA catalytic activity are probably not regulated by caveolin-1 interaction in these locations, the shear-induced regulation of eNOS may be directly under the control of PKA. Our data further suggest that at steady state, a portion of eNOS is phosphorylated in the Golgi area. A site-specific role of PKA in the Golgi may particularly be connected to the regulation of functional eNOS at the cell surface. cAMP-raising agents such as forskolin prevent protein export from the trans-Golgi network (Muniz et al. 1996), and more specifically prevent the trafficking of mature eNOS to plasma membrane caveolae (Belhassen et al. 1997).
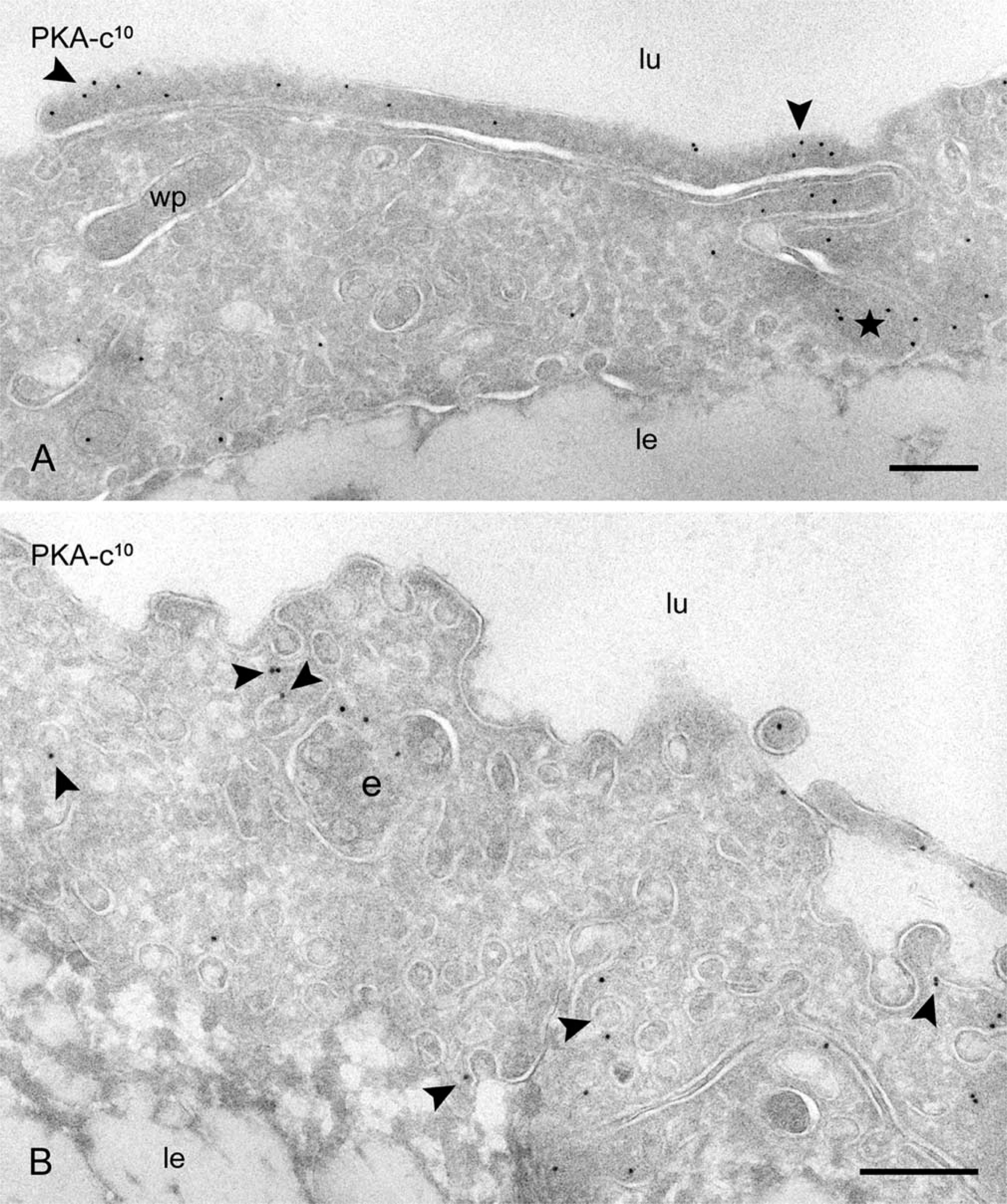
PKA-c is compartmentalized in distinct subcellular locations. (
Relative distribution of PKA-c a
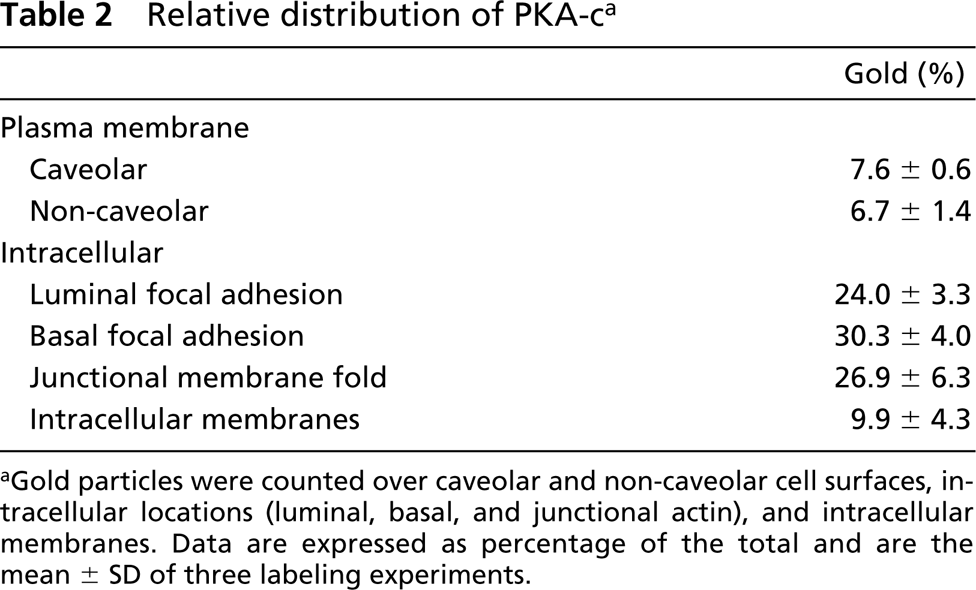
aGold particles were counted over caveolar and non-caveolar cell surfaces, intracellular locations (luminal, basal, and junctional actin), and intracellular membranes. Data are expressed as percentage of the total and are the mean ± SD of three labeling experiments.
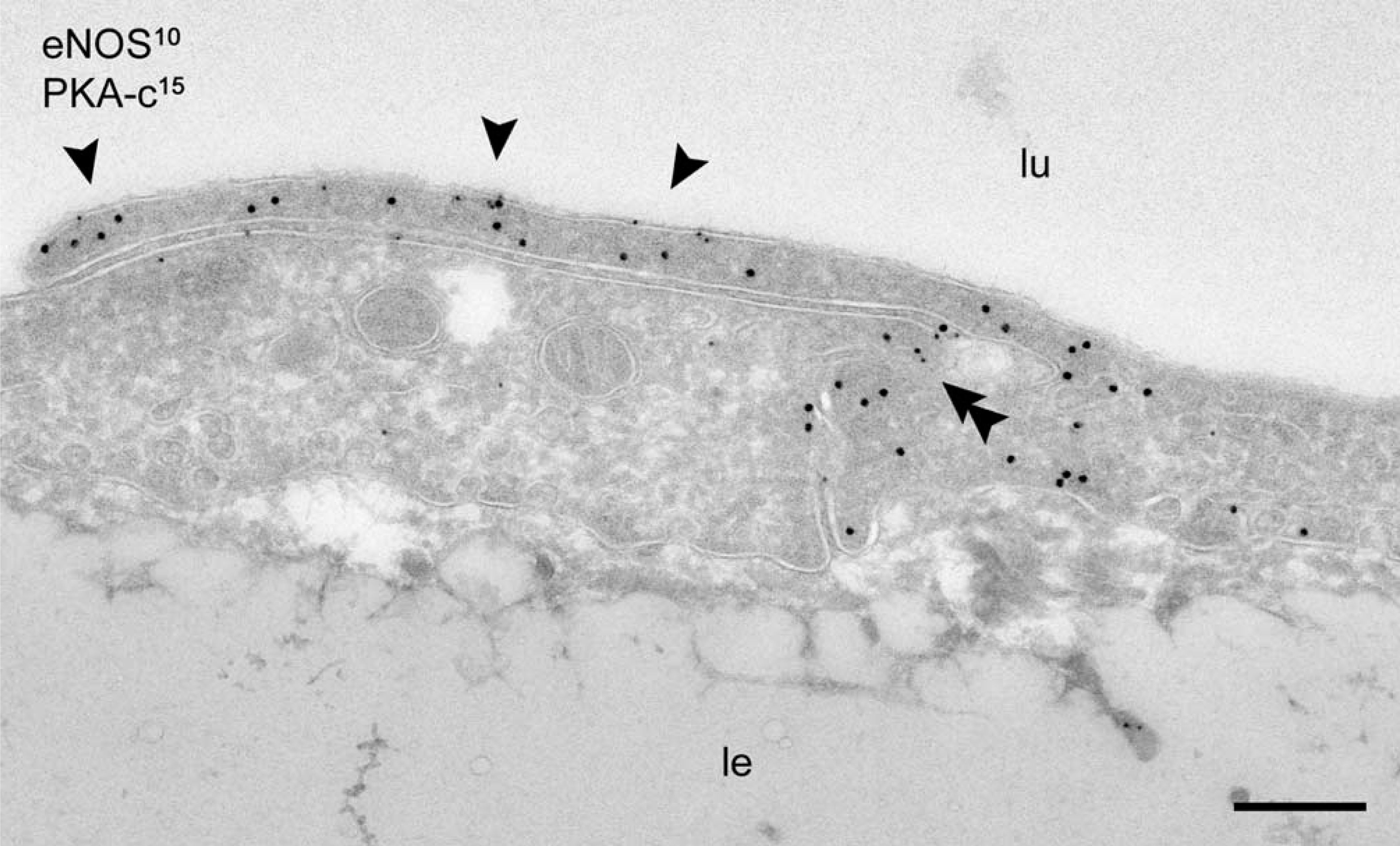
Colocalization of eNOS with PKA-c. Close proximity of PKA-c (15-nm gold) and eNOS (10-nm gold) in the peripheral lamellipodia (arrowheads) and at lateral endothelial cell contacts (double arrowhead). lu, lumen of aorta; le, lamina elastica, Bar = 200 nm.
In conclusion, we have demonstrated that, in rat aorta endothelium, a large portion of cell surface eNOS is located outside caveolae, in distinct endothelial cell junctions. Our findings are in agreement with those reported in rat endocardiac endothelium, where eNOS was predominantly found on peripheral cell borders (Andries et al. 1998). The absence of the inhibitor caveolin-1 and the close proximity of the catalytic form of PKA in these locations suggest a site-specific regulation of the enzyme independent of caveolin-1. In addition to luminal caveolae, changes in vascular tone and blood flow may also be sensed in these specialized endothelial cell junctions and hence regulate the flow-responsive NO production in a PKA-dependent manner. Such a mechanism is consistent with recent findings in cultured endothelial cells where eNOS localization and NO production have been reported to dramatically depend on the existence of cell–cell contacts (Sowa et al. 1999; Govers et al. 2002a). Further studies are required to determine which pool of PKA is responsible for the shear-related NO production.
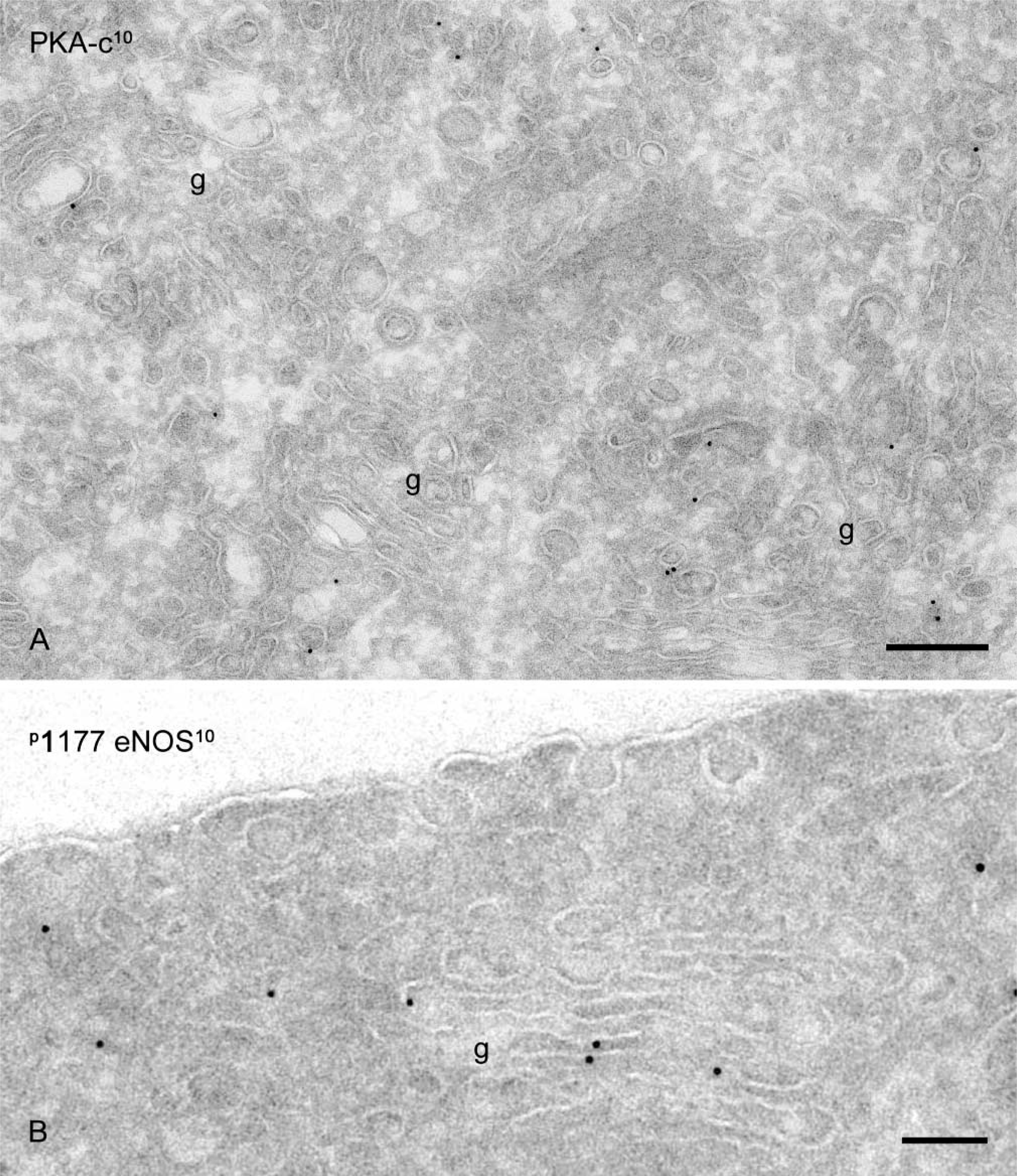
Golgi association of PKA-c and phospho-eNOS. (
Footnotes
Acknowledgements
We thank Dr Catherine Rabouille for critical reading of the manuscript, and Rene Scriwanek and Marc van Peski for excellent photographic work.