
Abstract
Infections with enterotoxigenic Escherichia coli (ETEC) are a major cause of travelers' diarrhea worldwide. Colonization of the small intestine mucosa is dependent on specific colonization factor antigens (CFA) and coli surface (CS) antigens. CFA/1, CS3, and CS6 are the most prevalent fimbrial antigens found in clinical isolates. The goal of our study was to visualize the morphology of CS3 and CS6 fimbriae in wild-type and recombinant E. coli strains by means of transmission electron microscopy in conjunction with negative staining and immunolabeling. Corresponding ETEC genes were cloned into E. coli K12 strain DH10B. Expression of fimbriae was dependent on culture conditions and sample handling. Specific immunolabeling of fimbriae unequivocally demonstrated the presence of all types of surface antigens investigated. Negative staining was effective in revealing CS3 but not CS6. In addition, this technique clearly demonstrated differences in the morphology of genetically and immunologically identical CS3 surface antigens in wild-type and recombinant strains. This paper provides a basis for the assessment of recombinant vaccines.
S
The goal of the present study was to establish appropriate methods to demonstrate the expression of surface antigens in recombinant strains of E. coli. A method of negative staining and immunogold labeling of a recombinant acceptor E. coli-K12 strain DH10B expressing either CS3 or CS6 was adopted to monitor the presence and morphology as well as the antigenic potential of fimbriae as compared with selected wild-type strains. The present study clearly demonstrates the value of this approach for the validation of recombinant strains in view of their potential as vaccine candidates.
Materials and Methods
Investigated wild-type strains and recombinant strains, as well as plasmids used for their transformation, are specified in Table 1.
Culture Conditions and Collection of Bacteria in Suspension
After testing various protocols, bacteria were grown in CFA medium (Evans et al. 1979) overnight at 37C. Antibiotics were included as appropriate according to Table 1 (34 μg/ml chloramphenicol, 50 μg/ml kanamycin, and 50 μg/ml ampicillin either alone or in combination). Bacteria were then rinsed off the culture plates with 0.85% NaCl, which had been prewarmed to 37C. Bacterial suspension was fixed immediately by adding an equal volume of paraformaldehyde fixative (final concentration: 0.1% paraformaldehyde in 0.01 MPBS, pH 7.4) or diluted Karnovsky's fixative (final concentration: 2.5% glutaraldehyde, 2% paraformaldehyde in 0.1 M cacodylate buffer, pH 7.4), respectively. Later experiments showed that fixation is unnecessary, and that it can be omitted without detrimental effects. Bacteria were adsorbed onto parlodioncoated copper or nickel grids by placing a grid upside down onto a drop of bacterial suspension. Adsorbed bacteria were washed once or twice on a drop of dH2O for 5 min.
Negative Staining
Negative staining was performed by floating the grids on a drop of 0.25% phosphotungstic acid (PTA) with 0.01% BSA in dH2O for 30-40 sec. Excess liquid was blotted off with filter paper, and samples were air dried.
Immunolabeling
Samples were initially blocked for 20 min with PBS containing 0.2% BSA and 0.05% Tween 20 (PBS-BT). As this step was shown to be unnecessary to prevent background labeling, it was omitted in later experiments. Antibodies used included mouse monoclonal anti-CS3 as well as rabbit polyclonal anti-CS3 and anti-CS6 antibodies. All antibodies were raised against purified antigens (kindly provided by Dr. F. Cassels, National Institutes of Health, Bethesda, MD). Antigens were either fimbriae purified according to Yu et al. (2002) or whole cells producing the recombinant fimbriae. Rabbit sera were preadsorbed against a panel of strains to remove irrelevant antibodies. Specificity of antibodies used was then assessed on Western blots (Burnette 1981). Host bacteria lacking expression plasmids were used as negative controls.
Properties of bacterial strains
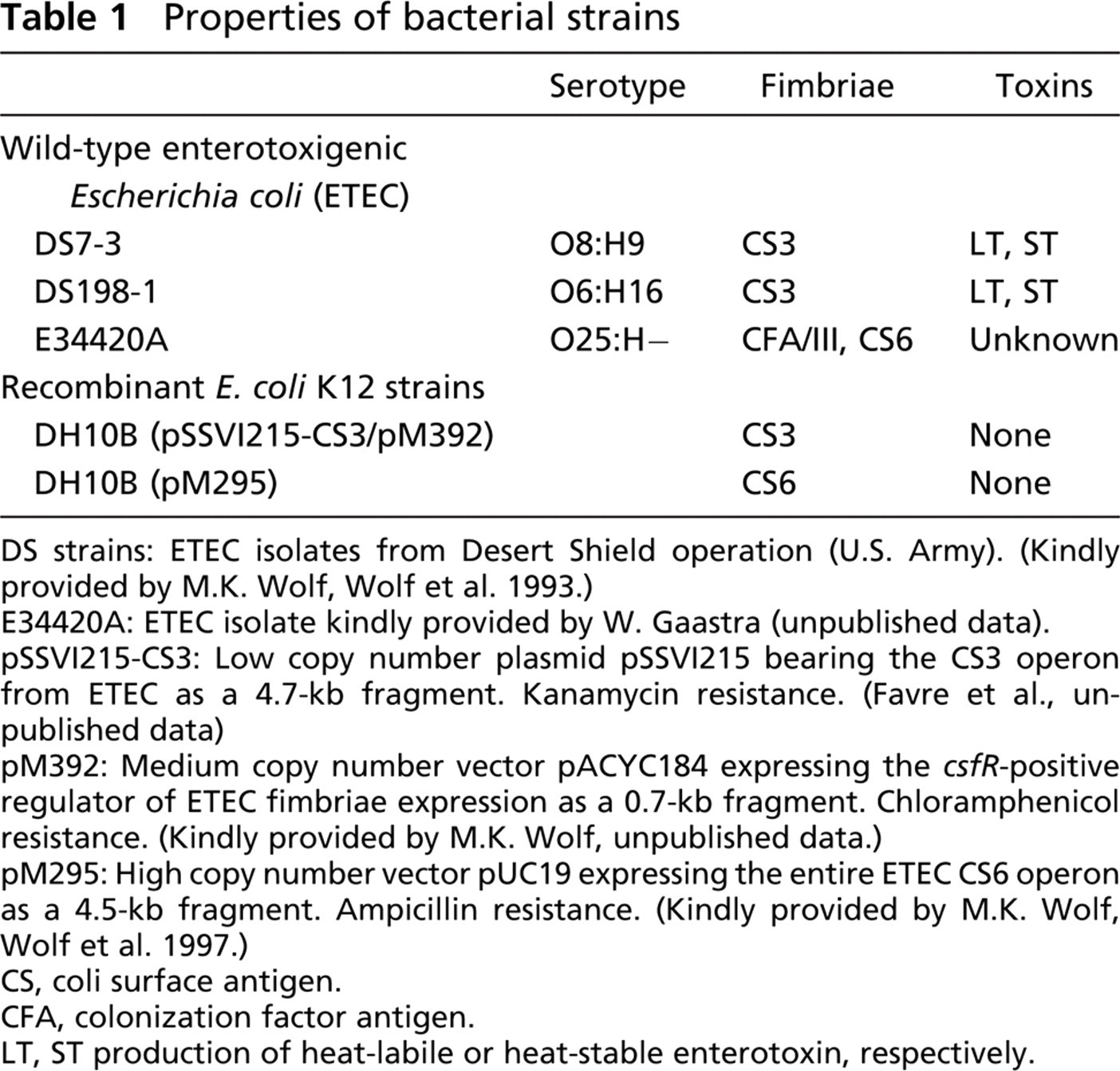
DS strains: ETEC isolates from Desert Shield operation (U.S. Army). (Kindly provided by M.K. Wolf, Wolf et al. 1993.)
E34420A: ETEC isolate kindly provided by W. Gaastra (unpublished data).
pSSVI215-CS3: Low copy number plasmid pSSVI215 bearing the CS3 operon from ETEC as a 4.7-kb fragment. Kanamycin resistance. (Favre et al., unpublished data)
pM392: Medium copy number vector pACYC184 expressing the csfR-positive regulator of ETEC fimbriae expression as a 0.7-kb fragment. Chloramphenicol resistance. (Kindly provided by M.K. Wolf, unpublished data.)
pM295: High copy number vector pUC19 expressing the entire ETEC CS6 operon as a 4.5-kb fragment. Ampicillin resistance. (Kindly provided by M.K. Wolf, Wolf et al. 1997.)
CS, coli surface antigen.
CFA, colonization factor antigen.
LT, ST production of heat-labile or heat-stable enterotoxin, respectively.
For immunogold labeling, grids were floated on corresponding primary antibodies for 60 min (diluted 1:100 or 1:50 in PBS-BT). Grids were washed in PBS-BT for 5 min and incubated for another 60 min on a drop of either 12 nm- or 15 nm-gold-conjugated goat anti-mouse IgG or goat anti-rabbit IgG (both from British Biocell International; Braunschweig, Germany) diluted 1:100 in PBS-BT, respectively. Finally, grids were washed twice in dH2O for 5 min. After blotting off excess liquid with filter paper, negative staining, and air drying, grids were examined in a Zeiss (Oberkochen, Germany) 109 transmission electron microscope equipped with a wide-angle slow scan CCD Camera 689 (GATAN GmbH; Munich, Germany).
Control experiments included negative staining of unaltered host strain DH10B and labeling of recombinant strains, whereby monoclonal and polyclonal primary antibodies were substituted with an irrelevant primary antibody.
Results
Negative Staining and Immunolabeling of CS3 ETEC DS7-3 (Figures 1A and 1B) and DS198-1 (Figures 1C and 1D) were used as reference strains for the characterization of the CS3 surface antigen. CS3 surface antigens showed a thin and curly morphology (Figures 1A and 1C). Likewise, immunogold labeling with both monoclonal and polycolonal antibodies was highly specific and, thus, unequivocally confirmed these hair-like structures to be CS3 (Figures 1B and 1D).
In contrast to their appearance in wild-type strains, CS3 surface antigens produced variable electron-dense clusters in recombinant strain DH10B(pSSVI215-CS3/pM392) (Figure 1E) as demonstrated by negative staining. However, their presence was clearly demonstrated by immunolabeling (Figure 1F). Furthermore, labeled debris was commonly seen in between the bacteria (Figure 1F).
Negative Staining and Immunolabeling of CS6
As for CS6 fimbriae, negative staining only provided evidence of a narrow margin of PTA surrounding the bacteria. Consequently, only one type of surface antigen, i.e., the long and wavy CFA/III surface antigen, was depicted morphologically in bivalent wild-type strain E34420A (Figure 2A). However, immunogold labeling with polyclonal antibodies yielded convincing evidence of CS6 surface expression. Interestingly, labeling of CS6 was superimposed over the cell body, and gold particles were only occasionally seen away from the bacteria (Figure 2B).
In accordance with the observations made on wild-type strains, CS6 fimbriae also could not be demonstrated on recombinant strain DH10B(pM295) (Figure 2C). Notwithstanding, the specific labeling signal obtained with polyclonal rabbit anti-CS6 antibodies (Figure 2D) was similar to the labeling density seen in wild-type strain E34420A (Figure 2B).
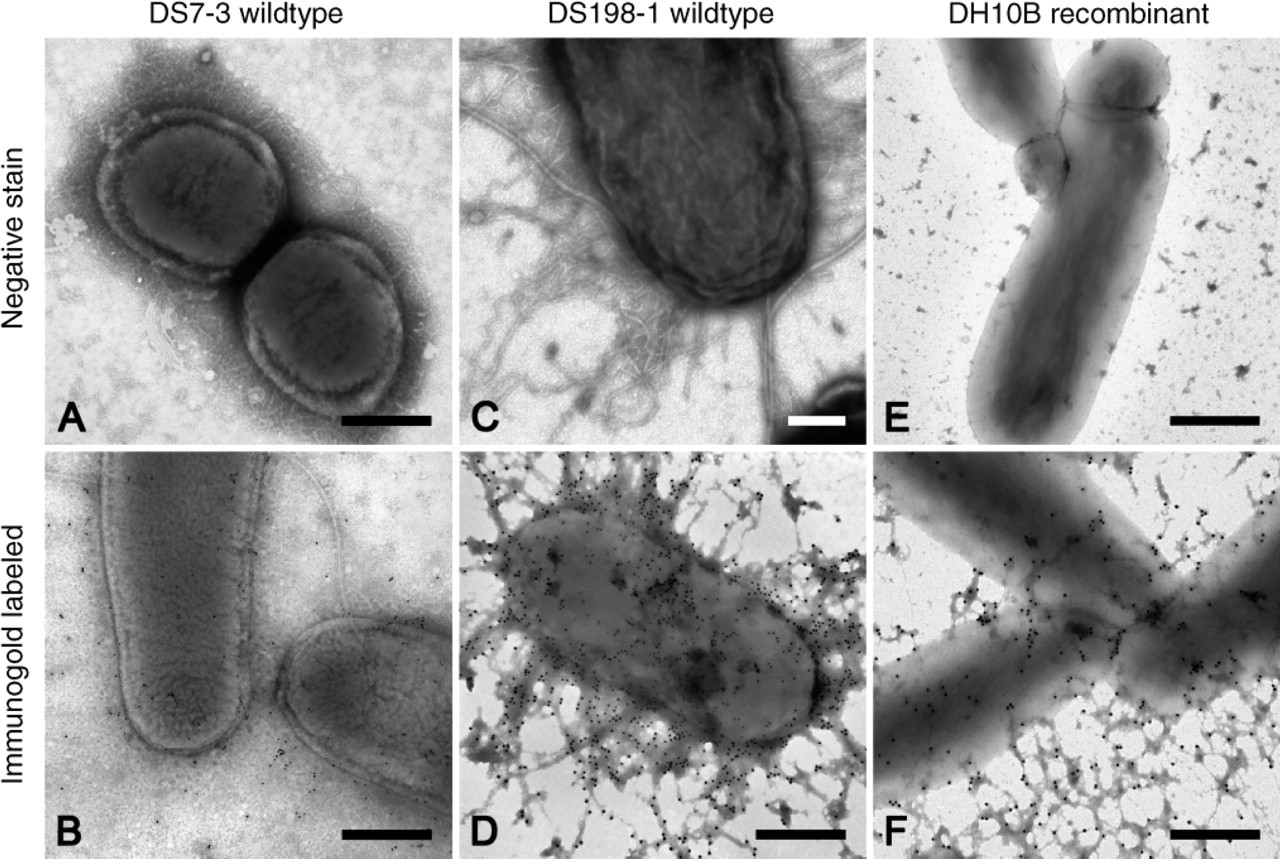
Expression of CS3 antigens in wild-type strains DS7-3 (
Controls
All control strains were devoid of background labeling (Figure 3).
Discussion
Evidence for the presence of a gene, its translation into a protein, and indication of surface expression can be provided by molecular biology methods, slide agglutination, immunospots, immunofluorescence, and pili ELISA. However, the ultimate proof for proper expression in the correct location to elicit an immune response can only be granted by structural and immunologic analysis. Therefore, we used transmission electron microscopy in combination with negative staining and immunogold labeling to assess the expression of coli-specific CS3 and CS6 in recombinant E.coli-K12 strain DH10B. Expression of fimbriae was dependent on culture conditions and sample handling. Factors affecting morphological appearance of fimbriae included composition of culture medium, temperature, growing time, and centrifugation. These parameters were adjusted for maximum expression and optimal preservation of fimbriae and led to the protocol reported above.
Bacteria were grown in CF medium (Evans et al. 1979). Inclusion of lactose to this medium did not improve the expression of CS3 and CS6 fimbriae. Growing the bacteria overnight was adequate because experiments with incubation times of 5, 6, and 7 hr did not yield satisfactory expression of fimbriae. Both NaCl and PBS were adequate to preserve bacterial morphology. However, use of distilled water, 0.5 M Tris, or a protein-free culture medium did not provide satisfactory results.
Interestingly, small fimbriae such as CS3 and CS6 turned out to be more vulnerable in wild-type strains as compared with CFA I pili and even more so in recombinant strains. Obviously, expression of corresponding genes produces frailer fimbriae than in donor strains. Thus, avoiding any centrifugation steps by simply rinsing off the bacteria from the agar plates with NaCl substantially improved the preservation of fimbriae. But even after the most gentle treatment, CS3 fimbriae were partly lost in recombinant strain DH10B(pSSVI215-CS3/pM392). Obviously, shear stress caused by blotting off excess liquid from the grid with filter paper is enough to detach fimbriae from the bacterial body.
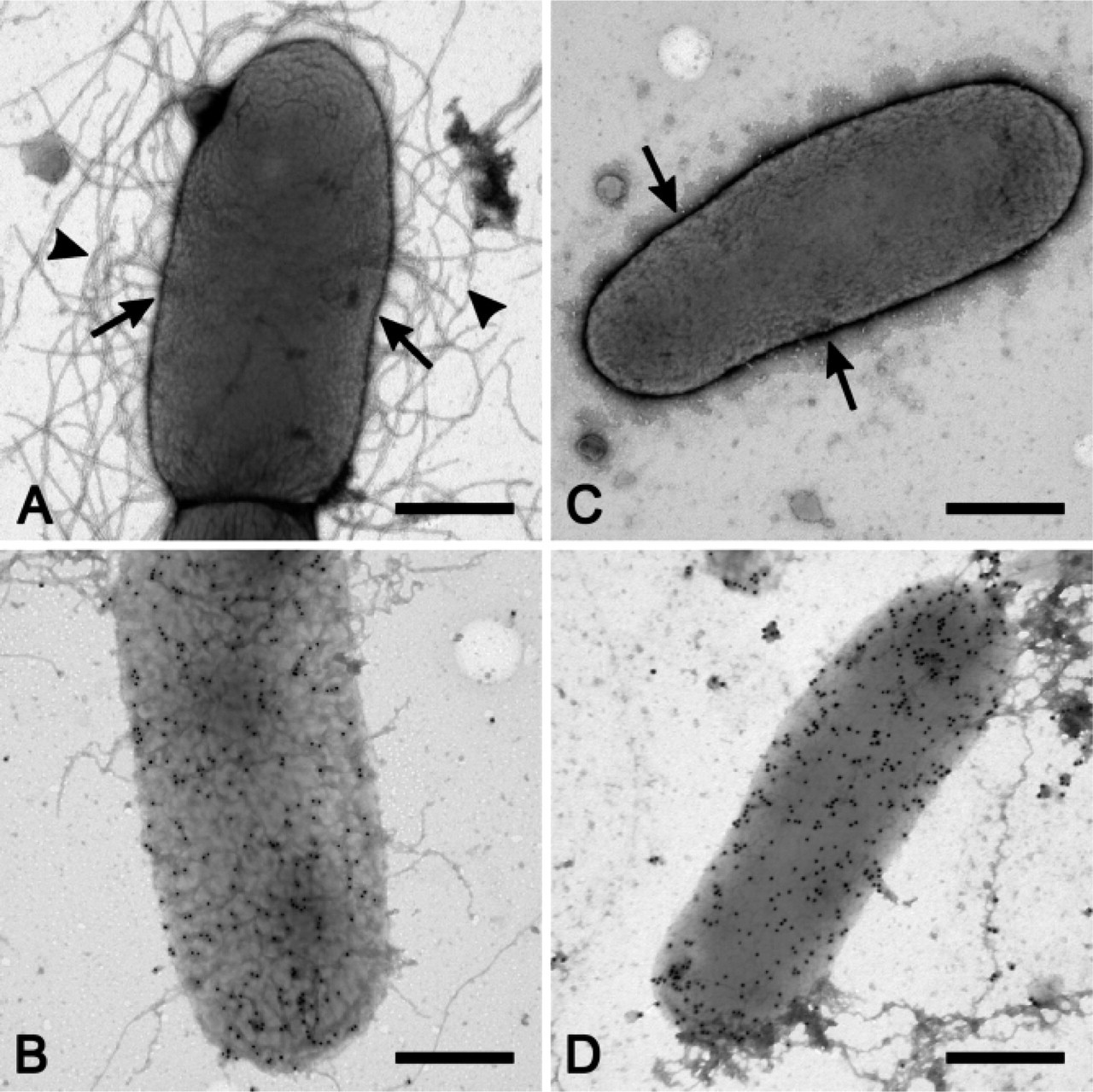
Expression of CS6 antigens in bivalent wild-type strain E34420A (A,B) and recombinant E. coli K12 strain DH10B(pM295) (
On the other hand, CS6 fimbriae seem to be so short that they do not protrude enough from the bacterial surface to be detectable by negative staining. However, a small fringe of PTA was retained, thus providing suggestive evidence for the presence of fimbriae. Presence of CS6 surface antigens was unequivocally demonstrated by conspicuous immunolabeling on strains E34420A and DH10B(pM295). This provided compelling evidence for the expression of immunologically identical CS6 on recombinant strains as compared with wild-type ETEC.
The current study presents an effective approach allowing the expression of surface antigens to be assessed both morphologically and immunologically. This phenotypical and immunological appraisal provides a valuable tool to investigate various factors affecting the expression of genes encoding surface antigens. In addition, morphological assessment of gene expression may be a crucial step in the development of new vaccines. Some live attenuated bacterial vaccine strains have been used for decades and, thus, have been characterized comprehensively. Such strains represent promising carriers into which the genes encoding potentially immunogenic proteins might be introduced. Any resulting new vaccine, however, needs to be carefully assessed with respect to proper gene expression at the protein and structural level. The method presented in this paper thus provides a basis for the validation of potential recombinant bacterial vaccines prior to clinical trials.
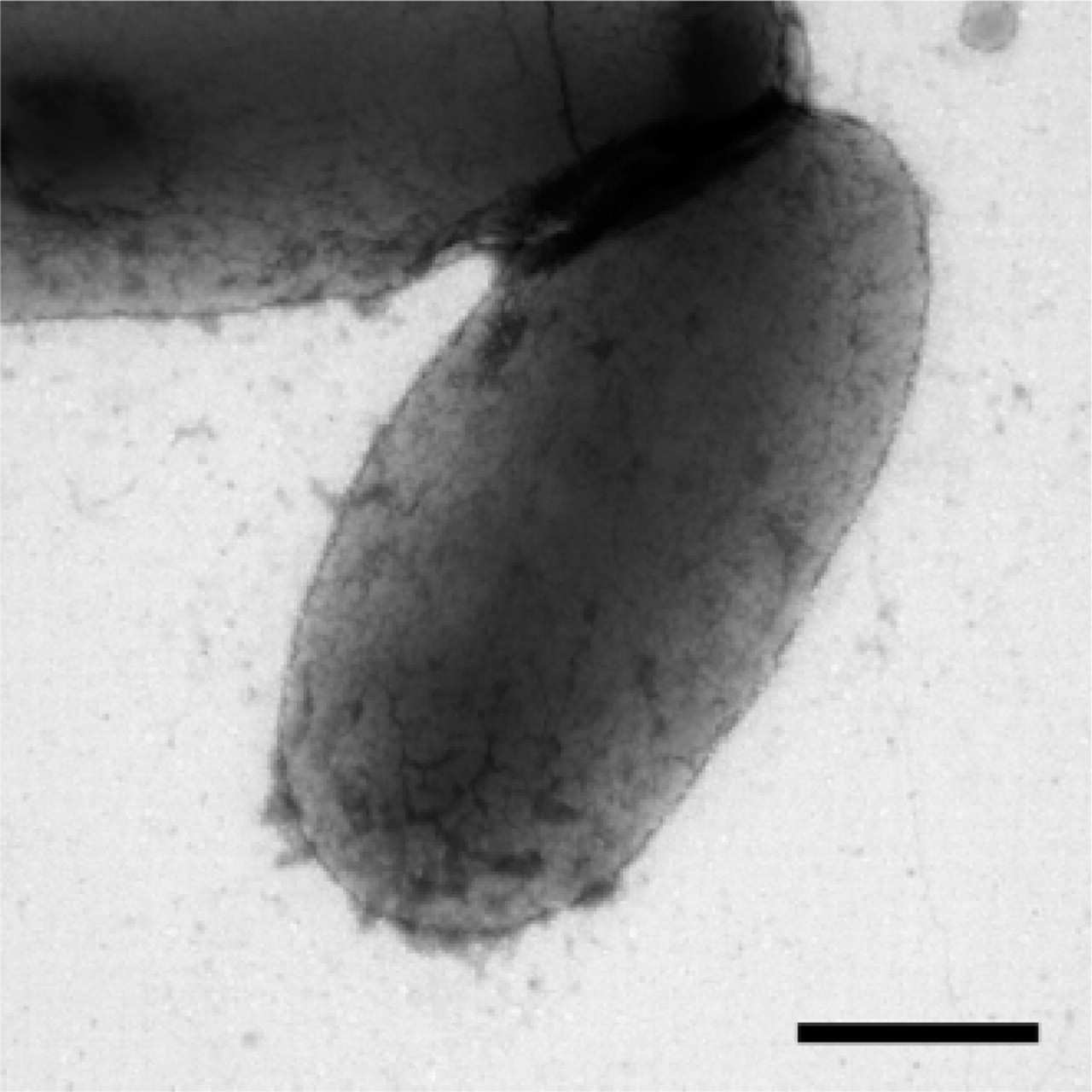
E. coli K12 strain DH10B(pSSVI215-CS3/pM392): substitution of primary antibody with rabbit anti-calcitonin antibody (negative control). Bar = 0.5 μm.
Footnotes
Acknowledgments
This study was supported by the Federal Office for Professional Education and Technology (OPET Grant KTI 7304.1 LSPP-LS).
The technical support of Cynthia Furer, Yvonne Schlatter, Corinne Hug, and Simon Knig is gratefully acknowledged. We are also indebted to M.K. Wolf for the generous gifts of ETEC strains DS7-3 and DS198-1 and for plasmids pM295 and pM392. ETEC strain E34420A was kindly and generously made available by W. Gaastra and the antisera against CS3 and CS6 by F. Cassels.