
Abstract
This study examines the relationship between inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) in the control of cervical ripening and parturition under normal (normal term pregnancy) and abnormal (preterm labor and prolongation of pregnancy) conditions by (a) measuring changes in the collagen both visually and quantitatively, (b) localizing and characterizing iNOS and COX-2 under normal conditions, and (c) characterizing the changes in iNOS and COX-2 under abnormal conditions. Cervices are obtained from estrus and timed pregnant Sprague-Dawley rats (n=4-10 per group). Preterm labor is induced with Onapristone (3 mg/rat; progesterone antagonist) and the prolongation of pregnancy with progesterone (2.5 mg, twice daily). Collagen changes are measured and visualized with the picrosirius polarization method. RT-PCR is used to characterize the mRNA expression (p<0.5), and immunohistochemistry is used to localize the protein expression for iNOS and COX-2. The organization and birefringence of the collagen during pregnancy decreased and is supported by changes in the luminosity (p<0.001). The iNOS and COX-2 enzymes were localized in cervical smooth muscle, vascular smooth muscle, and epithelium. Under normal conditions, iNOS mRNA levels decreased as COX-2 mRNA levels increased demonstrating an inverse correlation (Spearman r = −0.497; p=0.00295). Onapristone stimulated preterm labor, increasing the iNOS and COX-2 mRNA (p<0.05). The increase demonstrated a positive correlation (Spearman r = 0.456; p=0.03). Progesterone prolonged pregnancy, decreasing the iNOS and COX-2 mRNA (p=0.036). In conclusion, there may be an interaction between the nitric oxide and prostaglandin pathways in cervical ripening and parturition.
Keywords
C
Treatment with NO donors, such as isosorbide-mononitrate and glyceryl-trinitrate in humans, stimulate the ripening process (Thomson et al. 1997; Norman et al. 1998; Thomson et al. 1998). Inducible nitric oxide synthase (iNOS) is present in the cervix of many different species during pregnancy and increases during labor, suggesting a role for NO in the ripening process and parturition (Nathan 1992; Gilbert and Herschman 1993a,b; Kitajima et al. 1994; Vane et al. 1994; Buhimschi et al. 1996; Ali et al. 1997; Ledingham et al. 2000). Local applications of NO in animals and humans produce characteristics associated with the ripening process (Chwalisz et al. 1997; Thomson et al. 1997; Thomson et al. 1998; Shi et al. 2000b). N-nitro-L-arginine methyl ester, a nonspecific NOS inhibitor, prolongs time cervical dilation and decreases cervical extensibility. N-nitro-L-arginine methyl ester also prolongs delivery (Buhimschi et al. 1996; Shi et al. 2000b). These studies would suggest that NO plays an important role in the ripening of the cervix and parturition.
PGs are used to interrupt pregnancy in humans in the second trimester and also to stimulate cervical ripening at term (Szalay et al. 1981; Theobald et al. 1982; Ekman et al. 1983a,b,d,e; Calder and Greer 1992; Calder, 1994). Cyclooxygenase-2 (COX-2) is found in the cervix and sharply increases during labor or just before labor (Calder et al. 1977; Szalay et al. 1981; Ekman et al. 1983a-c,e; Calder and Greer 1992; Calder 1994; Dong et al. 1996; Chwalisz et al. 1997). Local applications of PGs in animals and humans produce characteristics associated with the ripening process (Ekman et al. 1983a; Chwalisz et al. 1997). PG inhibitors such as Indomethacin, a nonspecific COX inhibitor, prevent spontaneous abortions or preterm labor (Calder 1994; Hearne and Nagey 2000; Shi et al. 2000b; Bukowski et al. 2001). These studies would suggest that PGs play an important role in the ripening of the cervix and parturition.
These and other studies suggest that there may be an interaction between the NO and PG pathways in regulating the NOS and COX expression and this interaction may play a critical role in the control of cervical ripening and parturition (Salvemini et al. 1993; Appleby et al. 1994; Tetsuka et al. 1994; Zhang et al. 1998; Ekerhovd et al. 2002). Specifically, such interactions have been observed in various tissues and models. L-arginine, the substrate for NO production, produces an increase in PGE2 levels. Both NG-monomethyl-L-arginine, a nonspecific NOS inhibitor, and aminoguanidine, a specific iNOS inhibitor, decrease PGE2 levels after lipopolysaccharide stimulation in rat mesangial cells, whereas d-arginine had no effect. Aminoguanidine abolished NO production and decreased PGE2 production after interleukin (IL)-1β stimulation (Tetsuka et al. 1994). In RAW264.7 cells, PGE2 levels were also decreased in the presence of NG-monomethyl-L-arginine and aminoguanidine and in the absence of L-arginine. In RAW264.7 cells with an inactivated L-arginine NO pathway, NO donor sodium nitroprusside and glyceryl trinitrate increased PGE2 production after arachidonic acid stimulation (Salvemini et al. 1993). Isosorbide 5-mononitrate, another NO-donating compound, stimulated a significant increase in PGE2 in human cervical specimens (Ekerhovd et al. 2002). Hemoglobin, which binds and inactivates NO, blocked the increase in PGE2 production induced by L-arginine, NO, sodium nitroprusside, and glyceryl trinitrate. The activation site of COX contains an iron-heme center that NO may interact with to stimulate an increase in COX activity (Salvemini et al. 1993). These studies suggest a potential role for NO in the regulation of COX expression or activation and PG production.
Similarly, PGs, depending on type and concentration, may stimulate NOS expression or activation and NO production. PGs may influence NO activity through secondary messenger systems. Forskolin, which activates adenylate cyclase, increases nitrite or nitrate production, the byproduct of NO metabolism, in IL-β-induced rat mesangial cells. PGI2 activates adenylate cyclase, converting ATP to cAMP, which in turn may stimulate iNOS expression and NO production (Tetsuka et al. 1994). PGE2 may downregulate iNOS expression through changes in Ca+2, cAMP, or the protein kinase C pathways. Indomethacin enhances IL-1β-induced nitrite production by inhibiting PGE2 release, and increases IL-1β-induced steady-state iNOS mRNA levels and NO production. The effect of Indomethacin is reversed by exogenous PGE2 (Tetsuka et al. 1994). However, other studies show that PGE2 increases iNOS expression and NO production, suggesting that the effect of PGs on NOS expression and NO production may be concentration-dependent (Corbett et al. 1993; Salvemini et al. 1993; Franchi et al. 1994; Salvemini et al. 1994; Murakami et al. 1997). Carba-prostacyclin, a stable analog of PGI2, increased iNOS mRNA levels and IL-1β-induced NO production. The action of PGI2 may be mediated through the activation of adenylate cyclase. In mesangial and vascular smooth muscle cells, agents that increase cAMP also enhanced the cytokine-induced iNOS induction (Tetsuka et al. 1994). It has been shown that IL-1β stabilizes COX-2 mRNA by phosphorylation of cytosolic factors that bind to the AUUUA-rich 3′ end of COX-2. The iNOS mRNA has an AUUUA 3′ motif in the untranslated region that is considered to be an mRNA instability determinate. PGs may have similar effects to that of IL-1β stabilizing iNOS (Srivastava et al. 1994; Tetsuka et al. 1994). These studies suggest a possible interaction between NOS and COX expression. Similar interactions may be involved in the control of cervical ripening and parturition.
The objective of these studies is to examine the relationship between iNOS and COX-2 in the control of cervical ripening and parturition under normal (normal term pregnancy) and abnormal (preterm labor and prolongation of pregnancy) conditions. First, under normal conditions, we characterize the change in the collagen both visually and quantitatively (picrosirius red polarization method). We also localize (immunohistochemical staining) and characterize (RT-PCR) the expression patterns of iNOS and COX-2 during the ripening process and parturition in the rat cervix. Localization may suggest a spatiotemporal relationship between the iNOS and COX-2 enzymes. Characterization of the expression pattern quantitatively may suggest a relationship between the iNOS and COX-2 mRNA expression. Then, under abnormal conditions, we characterize the expression patterns of iNOS and COX-2: first, under preterm laboring conditions induced with the antiprogestin, Onapristone (ZK98299) and, second, after the prolongation of pregnancy with progesterone. Onapristone is used as a model for progesterone withdrawal and preterm labor (Edwards et al. 1995). Characterization of the expression pattern quantitatively may suggest a relationship between the iNOS and COX-2 mRNA expression under preterm laboring conditions. In contrast, progesterone is used as a model for the prolongation of pregnancy in which progesterone withdrawal does not occur. Characterization of the expression pattern quantitatively may suggest a relationship between the iNOS and COX-2 mRNA expression during the prolongation of pregnancy. Finally, the iNOS and COX-2 mRNA is compared under term vs preterm laboring conditions examining the role or importance of NO and PGs in regulating or initiating parturition. A better understanding of the mechanisms that regulate the ripening process and parturition may result in improved therapeutic and diagnostic options in both preterm cervical insufficiency and failure of cervical ripening at term.
Materials and Methods
Animal Treatment
Nonpregnant and timed-pregnant Sprague-Dawley rats (Charles River Laboratories; Wilmington, MA) were housed separately and allowed free access to food and water. Animals were maintained on a constant 12 hr light and dark cycle. Animals were killed by CO2 inhalation according to the protocols approved by the Animal Care and Use Committee at the University of Texas Medical Branch.
Tissue Preparation
Normal Term Pregnant Conditions. Cervices were obtained on days 14 (mid-gestation), 18 (late gestation), 22 nonlabor, and labor (one to three pups expelled) of pregnancy, and days 1 and 3 postpartum. Nonpregnant rat cervices obtained at estrus were used as control.
Preterm Labor. Cervices were obtained from timed pregnant Sprague-Dawley rats at 6, 12, 24, and 28 hr after Onapristone (3 mg/rat SC) or vehicle control (200 μl of benzyl benzoate [1]: sesame oil [4] solution) (Sigma; St. Louis, MO) treatment on day 17 of gestation (n=5-6 per group).
Prolongation of Pregnancy. Timed-pregnant rats were chronically treated with either progesterone (2.5 mg SC twice daily) or vehicle starting on day 18 of pregnancy. The animals were sacrificed 1 (control only, reference), 2, 3, 4, or 6 days after the initial treatment (n=6 per group).
Tissue Embedding for Immunohistochemistry (IHC) and the Picrosirius Red Polarization Method. The tissue was fixed in 4% paraformaldehyde in 0.1M phosphate buffer saline with CaCl2 for 30 min to an hour, trimmed, cut into longitudinal sections, and stored overnight in 0.1M phosphate buffer saline with CaCl2 at 4C. The tissue was then dehydrated, embedded in paraffin, sectioned at 5-6 μm, and placed on Superfrost/Plus microscope slides (Fisher Scientific; Fairlawn, NJ). The charged slide surface retains the tissue sections. Slides were dried at 60C for 30–45 min and stored at room temperature.
Preparation of RNA. Other cervical tissues were dissected, frozen in liquid nitrogen, and stored at −70C. Total RNA was extracted from the frozen tissue using the acid-guanidinium thiocyanate-phenol-chloroform method (Chomczynski and Sacchi 1987). All samples were treated with DNase I (Stratagene; La Jolla, CA) to remove any contaminating genomic DNA. The amount of RNA was calculated using the A260 value.
Visualizing and Measuring the Changes in Rat Cervical Collagen Using the Picrosirius Polarization Method
For picrosirius staining, sections were deparaffinized in xylene (EM Science; Gibbstown, NJ), rehydrated in a series of ethanol solutions (Fisher Scientific), and washed in running water for 10 min. Thorough washing of the sections before staining is essential because it allows the large Sirius red F3BA molecule to penetrate the connective tissue fibers.
Sections are stained with picrosirius red F3BA solution (Direct Red 80; Aldrich, Milwaukee, WI) (0.1% Sirius red F3BA in saturated aqueous picric acid) for 30 min. The dye solution must stand for 24 hr before being used. Positive connective tissue staining, except for elastic connective tissues, is indicated with a deep red colorization. All other tissue structures stain bright yellow (Figure 1) (Sweat et al. 1964). Tissues were dehydrated, cleared, and mounted using Cytoseal 60 (Fisher Scientific).
Sections were viewed with an Olympus U-STP polarizing light microscope (Olympus America; Melville, NY) (Figure 1; Sweat et al. 1964; Junqueira et al. 1979). An OLY-200 digital camera (Olympus America) was used with Adobe Premier 5.1 software (Adobe Systems Inc; San Jose, Ca) to obtain 10 (100x) low-power field images from the circular collagen of each cervix (Figure 1). The shutter speed (1/250 sec) and light source were fixed to compare brightness of the collagen birefringence between groups. As the crosslinks break down, the long strings of collagen are lost, reducing the ability of the collagen to reflect the polarized light. Images were analyzed for mean luminosity using the Adobe Photoshop 5.0 software (image: histogram; Adobe Systems Inc).
Localization of iNOS and COX-2 in Rat Cervical Tissue Using Immunohistochemical Staining
Standard IHC procedures were conducted. Sections were deparaffinized in xylene (EM Science), and ethyl alcohol (reagent grade; Pharmco, Brookfield, CT), and rehydrated in a graded series of ethanol solutions (95%, 70%, 50%) (Fisher Scientific). Sections were treated with 0.3% hydrogen peroxide to quench endogenous peroxidase activity and permeabilized with 0.1% Triton X-100 (Fisher Scientific) in PBS (pH 7.5). Nonspecific binding was blocked by incubating sections in PBS buffer (0.1 M) containing 0.025% BSA Type C (Sigma) and 5% normal horse serum (GIBCO Laboratories; Chagrin Falls, OH).
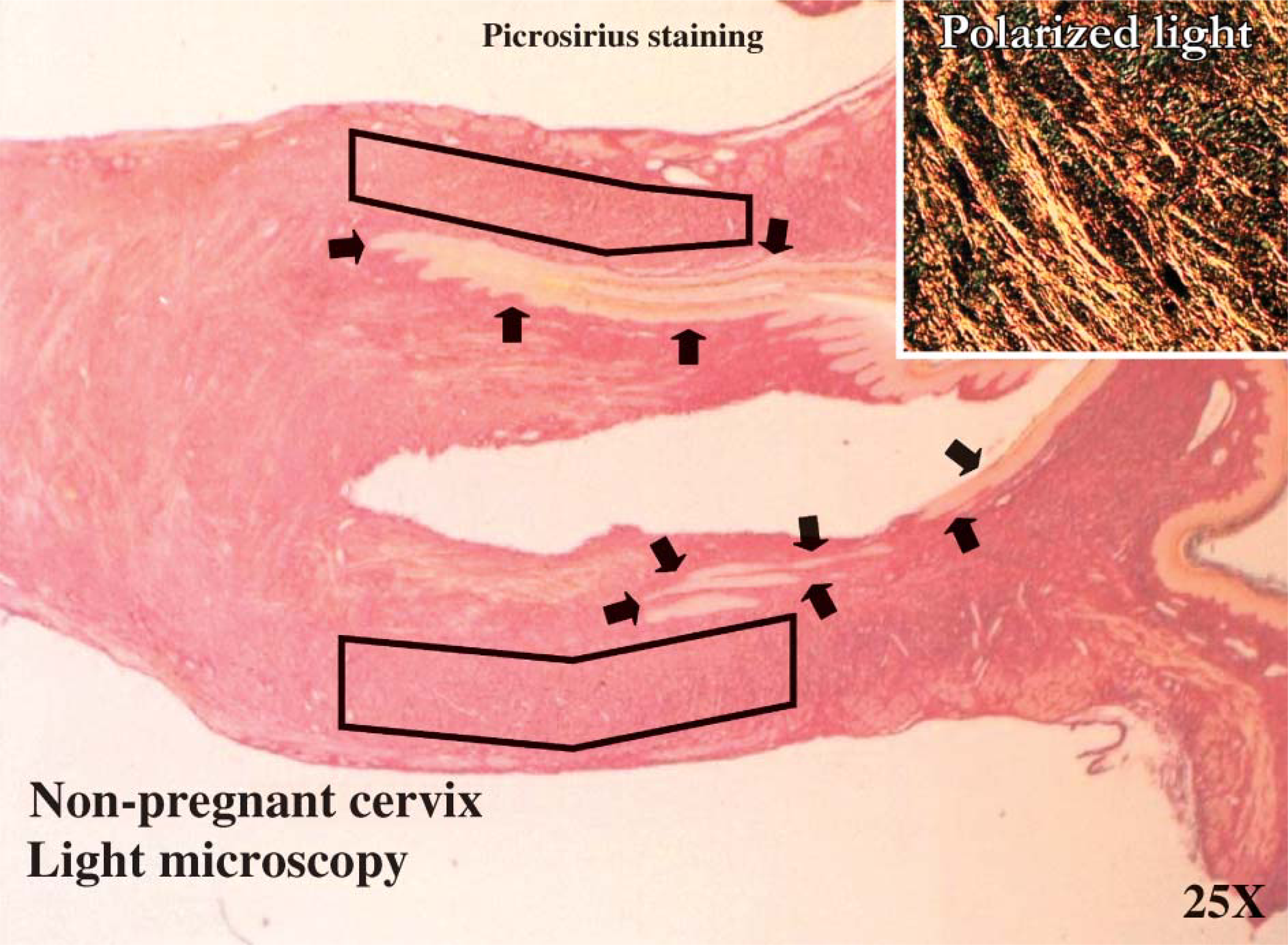
Picrosirius red staining under light microscopy (25x) and polarized light (100x). This is a digital image of the collagen stained with the picrosirius red that provides a representative indication of where the samples were obtained in the rat cervical tissues throughout gestation. The figure is of the estrus cervix. The arrows indicate where the two cervical canals are located. The black boxes indicate the approximate region within the tissue from which the samples were obtained.
iNOS. Sections used to detect iNOS expression were incubated overnight at 4C in rabbit polyclonal anti-iNOS antibody (Transduction Laboratories; Lexington, KY) at a 1:50 dilution. Sections were rinsed in PBS and then incubated with a biotinylated secondary antibody (Zymed Laboratories; San Francisco, CA). Aminoethyl carbazole (Zymed Laboratories) served as the chromagen to localize the iNOS enzyme.
COX-2. Sections used to localize the COX-2 enzyme expression were incubated for one hour at 37C with a goat polyclonal anti-COX-2 antibody (Santa Cruz Biotechnology; Santa Cruz, CA), at a 1:50 dilution. Sections were rinsed in PBS and then incubated with a biotinylated secondary antibody (Vector Laboratories; Burlingame, CA). Sections were then rinsed in PBS and incubated in an avidin-biotinylated peroxidase complex using the Vectastain Kit. 3,3′-Diaminobenzidine served as a chromagen to localize the COX-2 enzyme.
Sections of both iNOS and COX-2 preparations were lightly counterstained with Harris Modified Hematoxylin containing acetic acid (Fisher Scientific). The excess hematoxylin was removed with an acid-alcohol dip (0.5 ml HCl to 49.5 ml 70% ETOH) and then rinsed in ammonia water (2-3 drops of NH4OH to 50 ml of distilled water). Tissues were dehydrated in graded alcohol, placed in xylene, and mounted with either GVA mounting solution (Zymed Laboratories Inc.) for iNOS or Cytoseal 60 (Fisher Scientific) for COX-2.
IHC was performed for descriptive, but not quantitative purposes. Numerous sections were examined and representative tissues were photographed using a Nikon Microscope (FXA EP1-FL3; Nikon Microphot, Tokyo, Japan). Tissues from a minimum of four animals were examined at each time point with the distal portion of the cervix evaluated.
Using Real-time RT-PCR to Characterize the iNOS and COX-2 Cervical mRNA Expression
Primers and probes (Table 1) specific for either iNOS or COX-2 were designed using the Primer Express software (Applied Biosystems, Foster City, CA) with the rat iNOS (Acc # L12562) and COX-2 (Acc #L20085) as reference sequences. The selected primers and probes served as queries in blast analyses to confirm specificity (reference identification number: iNOS–forward primer 988309628-20826-3898, reverse primer 988310400-814-12100, and probe 1049152419-02113-5929; COX-2 - forward primer 961013803-12918-21846, reverse primers 1049149992-02684-14705, and probe 1049153173-010389-12316).
Primers and probe concentration and sequence information specific for iNOS and COX-2
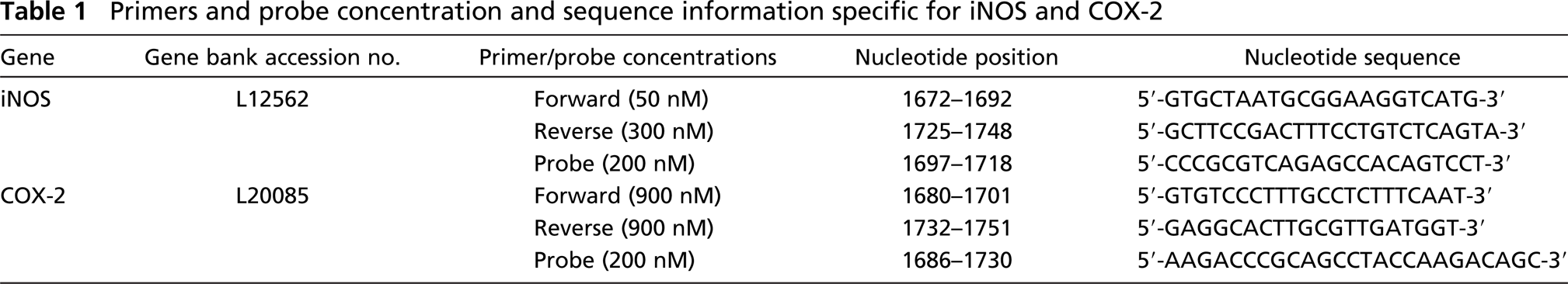
One-step RT-PCR assays were carried out using reagents provided in a TaqMan PCR Core Kit along with an RNase inhibitor and a Multiscribe Reverse Transcriptase (Applied Biosystems; Foster City, CA). Final primer and probe sequences and concentrations for iNOS and COX-2 are listed in Table 1. The iNOS 18SrRNA concentration was 25 nM each for the forward and reverse primers and probe. The COX-2 18SrRNA concentration was 50 nM each for the forward and reverse primers and probe. Each sample contained 100 ng of total RNA in a final volume of 50 μl and was reverse transcribed for 30 min at 48C, followed by AmpliTaq Gold activation (Applied Biosystems) at 95C for 10 min. Real-time PCR amplification was measured by the ABI Prism 7700 Sequence detection System (Applied Biosystems) under the following conditions: denaturation at 95C for 15 sec and annealing/extension at 60C for 1 min for 40 cycles.
Tissues from 4 to 10 animals were analyzed in each experimental group. Relative RNA levels were calculated using the ΔCT method described in Taqman Users Bulletin #2 (Applied Biosystems). The data were normalized against the internal 18SrRNA control (Applied Biosystems). Results are expressed as the average % of cervical iNOS or COX-2 mRNA expression relative to the levels of the estrus cervix (normal term pregnant study); the 6-hr control cervix (Onapristone study/preterm labor); the day 19 control cervix (progesterone study/prolongation of pregnancy); or the estrus cervix (term labor), the 28-hr control cervix (preterm labor) and the cervix at estrus (comparing term vs preterm labor) ± SEM.
Statistical Analysis
Statistical comparison between groups was performed using a one-way ANOVA followed by either the Student-Newman-Keuls test (picrosirius polarization study) or a Tukey test for multiple comparisons and a Spearman's correlation (normal term pregnant study and Onapristone/preterm labor study); a two-way ANOVA followed by a Tukey test (progesterone/prolongation of pregnancy) (significance: p<0.05); or a t-test or a Mann-Whitney Rank Sum Test (term versus preterm labor comparison study-normal/term: iNOS p=0.022, COX-2 p=0.01; Onapristone/preterm: iNOS p=0.005, COX-2 p=0.004; term versus preterm: iNOS p=0.004, COX-2 p=0.874 (Sigma Stat; Jandel Scientific Software, San Rafael, CA).
Results
Visualizing and Measuring the Changes in the Collagen
Figure 1 provides a representative indication of where the samples were obtained in the rat cervical tissues throughout gestation. It also demonstrates staining with the picrosirius red under light microscopy (25x) and polarized light (100x).
The changes in the organization of the collagen are visualized based on changes in the birefringence of the collagen (Figure 2). Luminosity is used to measure changes in the collagen based on the brightness of the birefringence (Figure 3). There is a progressive decrease in the organization reflected by the loss of birefringent intensity of the collagen from the nonpregnant state (Figure 2A), through pregnancy (Figures 2B–2D), which then appears to increase in the postpartum cervix (Figures 2E and 2F). In samples from the estrus (Figure 2A) and day 14 (Figure 2B) cervix, the collagen appears to be dense and highly organized with a high birefringence (Figure 3). On day 18 of gestation (Figure 2C), term (Figure 2D) and postpartum day 1 (Figure 2E) cervix, the collagen appears to be dispersed and disorganized with a significantly low birefringence (Figure 3). The birefringence (Figure 3) and the organization of the collagen within the cervix appeared to return to estrus levels (Figure 2A) in the postpartum day 3 cervix (Figure 2F).
Localization of Cervical iNOS and COX-2 during Gestation
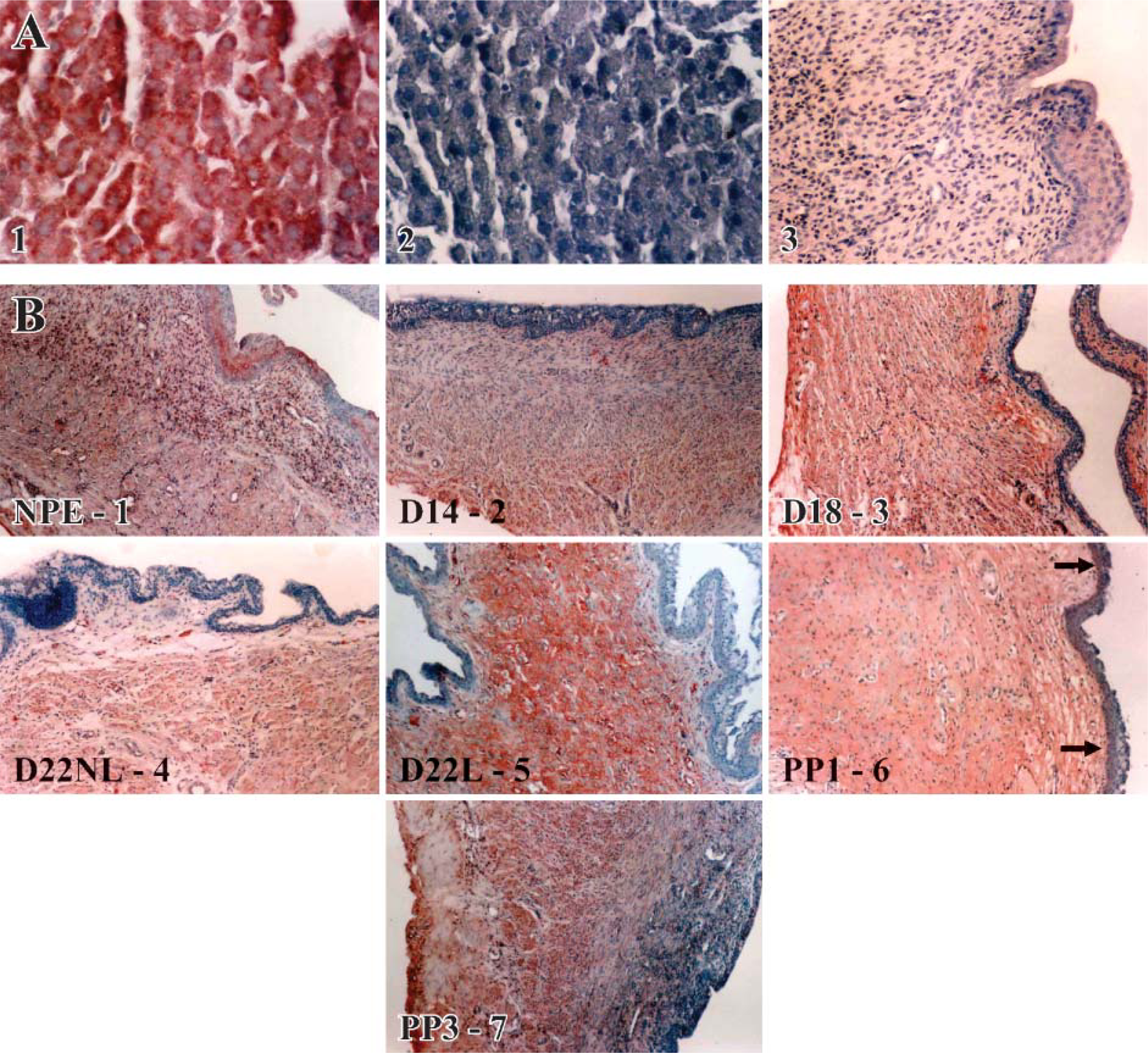
Controls for iNOS and COX-2. Figures 4A1–4A3 is the control tissues for the localization of the iNOS enzyme using immunohistochemical staining. Rat liver was used for the iNOS IHC positive control (Figure 4A1) A red to brown reaction product is an indication of positive staining for the iNOS enzyme. Normal rabbit IgG (Santa Cruz Biotechnology) was substituted for the primary antibody (Figure 4A2, rat liver) or the primary antibody was omitted in the negative control sections (Figure 4A3, estrus rat cervix).
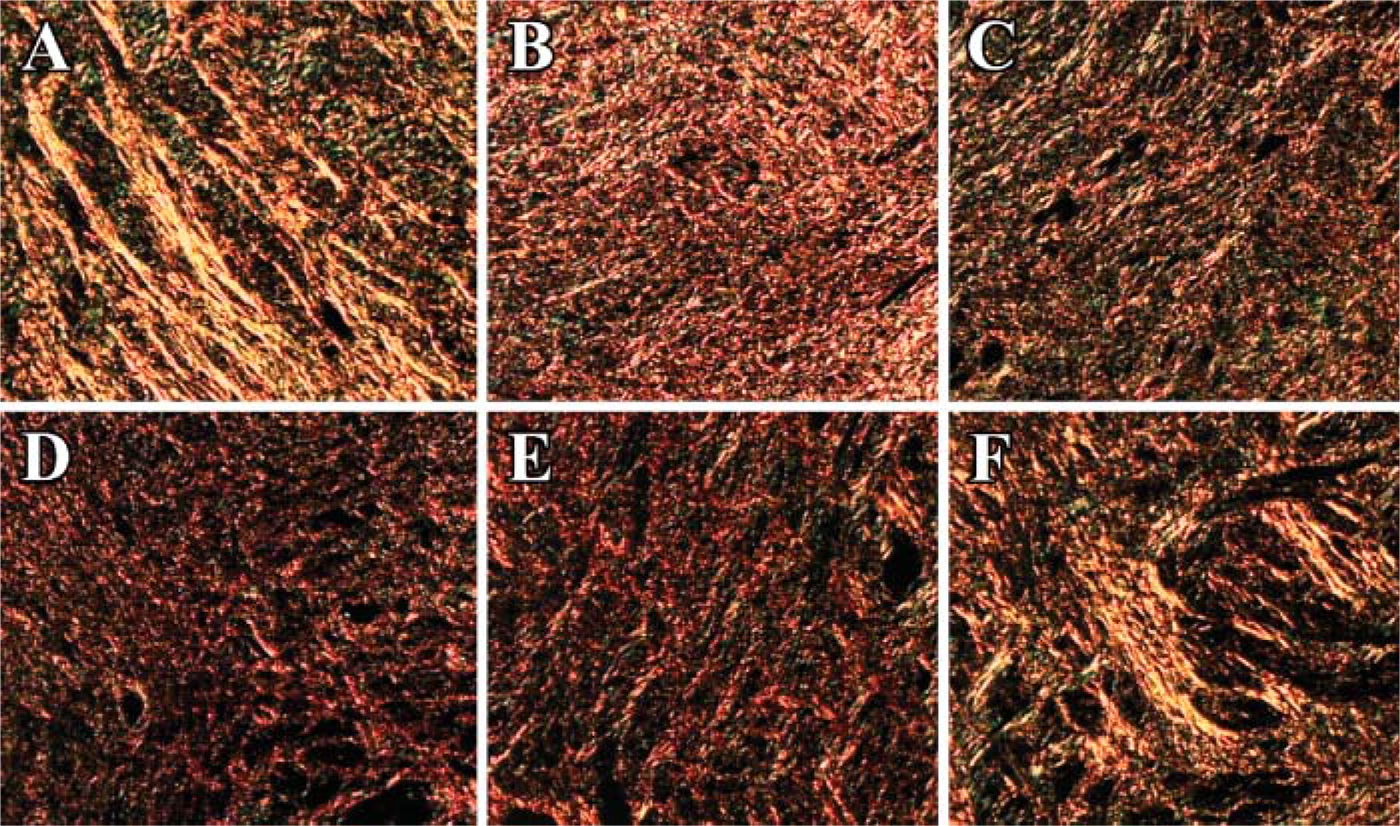
Changes observed were based on the organization and birefringence of the picrosirius red-stained collagen within the rat cervix throughout gestation using digital images (100X). (
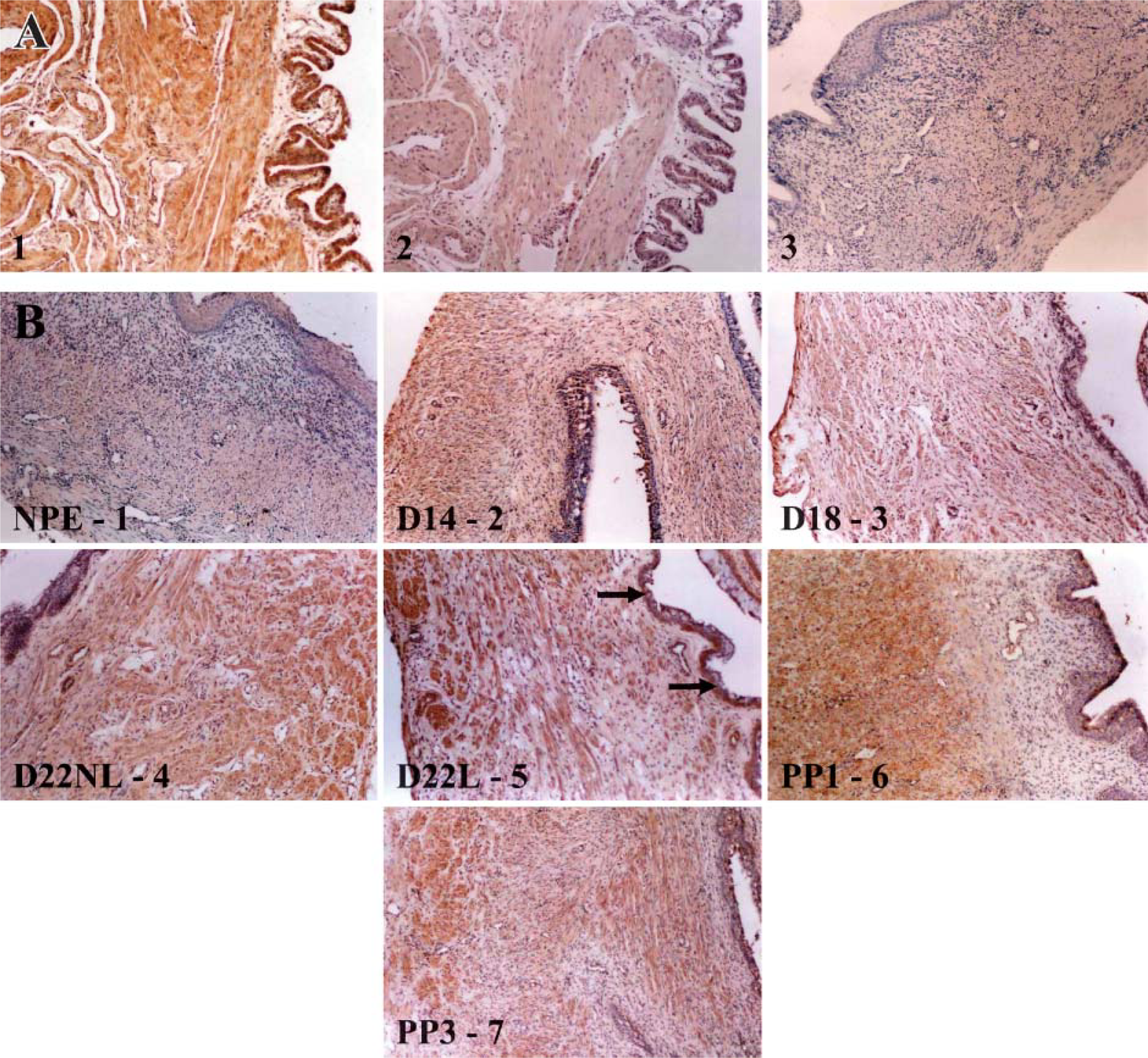
Figures 5A1-5A3 is the control tissues for the localization of the COX-2 enzyme using immunohistochemical staining. Day 22 laboring (D22L) rat uterus or kidney was used for COX-2-positive control sections (Figure 5A1, D22L rat uterus). A brown reaction product is an indication of positive staining for the COX-2 enzyme. Normal goat IgG (Santa Cruz Biotechnology) was substituted for the primary antibody (Figure 5A2, D22L rat uterus) or the primary antibody was omitted in the negative control sections (Figure 5A3, estrus rat cervix).
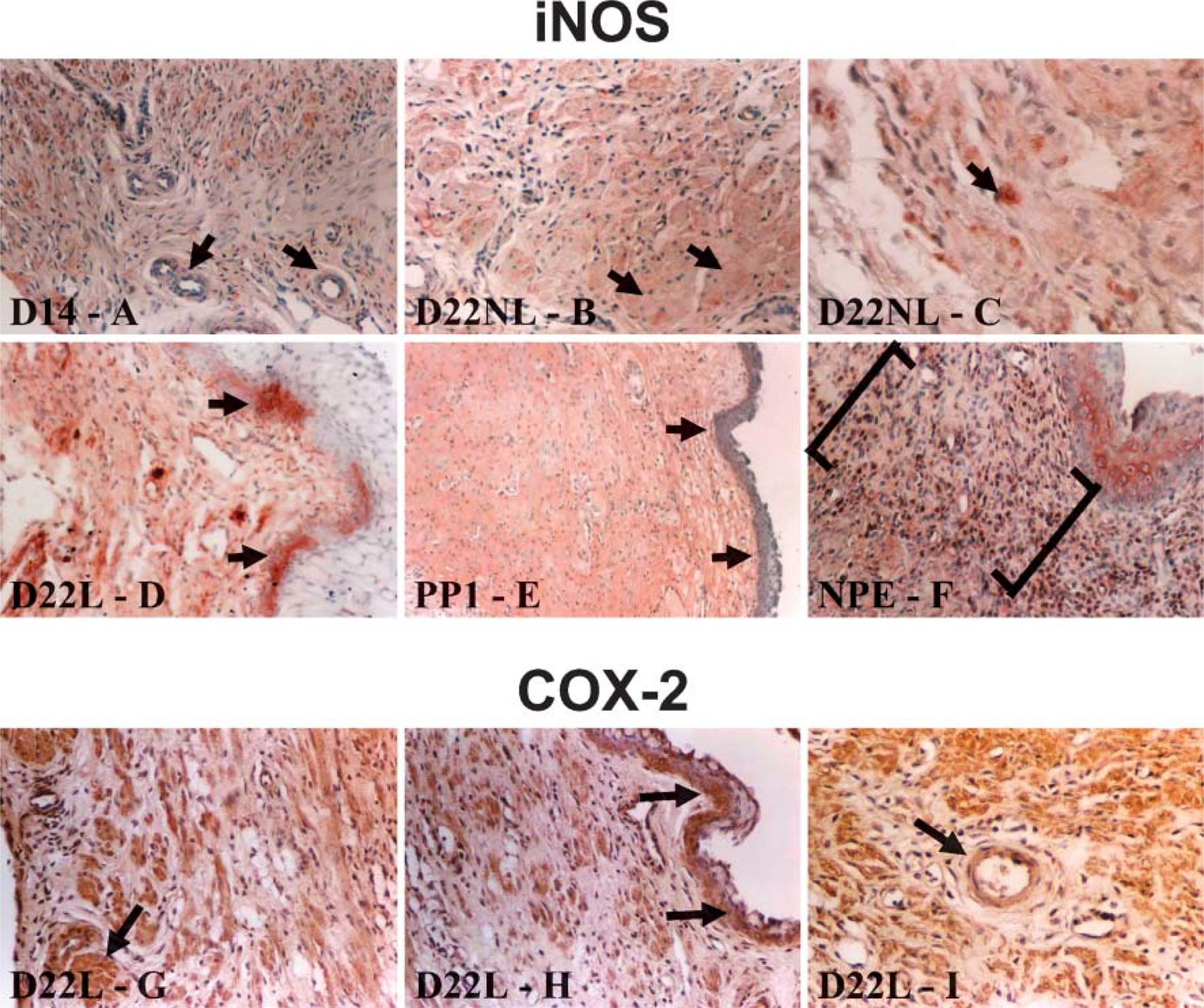
iNOS. Figures 4B1-4B7 are representative tissues demonstrating the localization sites for the iNOS enzyme throughout gestation in the rat cervix. Labeling for iNOS was evident in the cervical tissues at all time points examined. The iNOS enzyme was most prominent in the cervical smooth muscle (Figure 4B and Figure 6B). Expression of iNOS was also localized in the vascular smooth muscle (Figure 4B2 and Figure 6A). There was also labeling of large leukocyte or monocyte-like cells (Figure 6C). Epithelial labeling occurred in the postpartum day 1 cervix (Figure 4B6 and Figure 6E) and occasionally in the laboring cervix (Figure 6D). iNOS in the estrous cervix was present in the cervical smooth muscle, the vascular smooth muscle, the epithelium, and within a band of unknown cells located in the stroma, possibly fibroblasts (Figure 4B1 and Figure 6F).
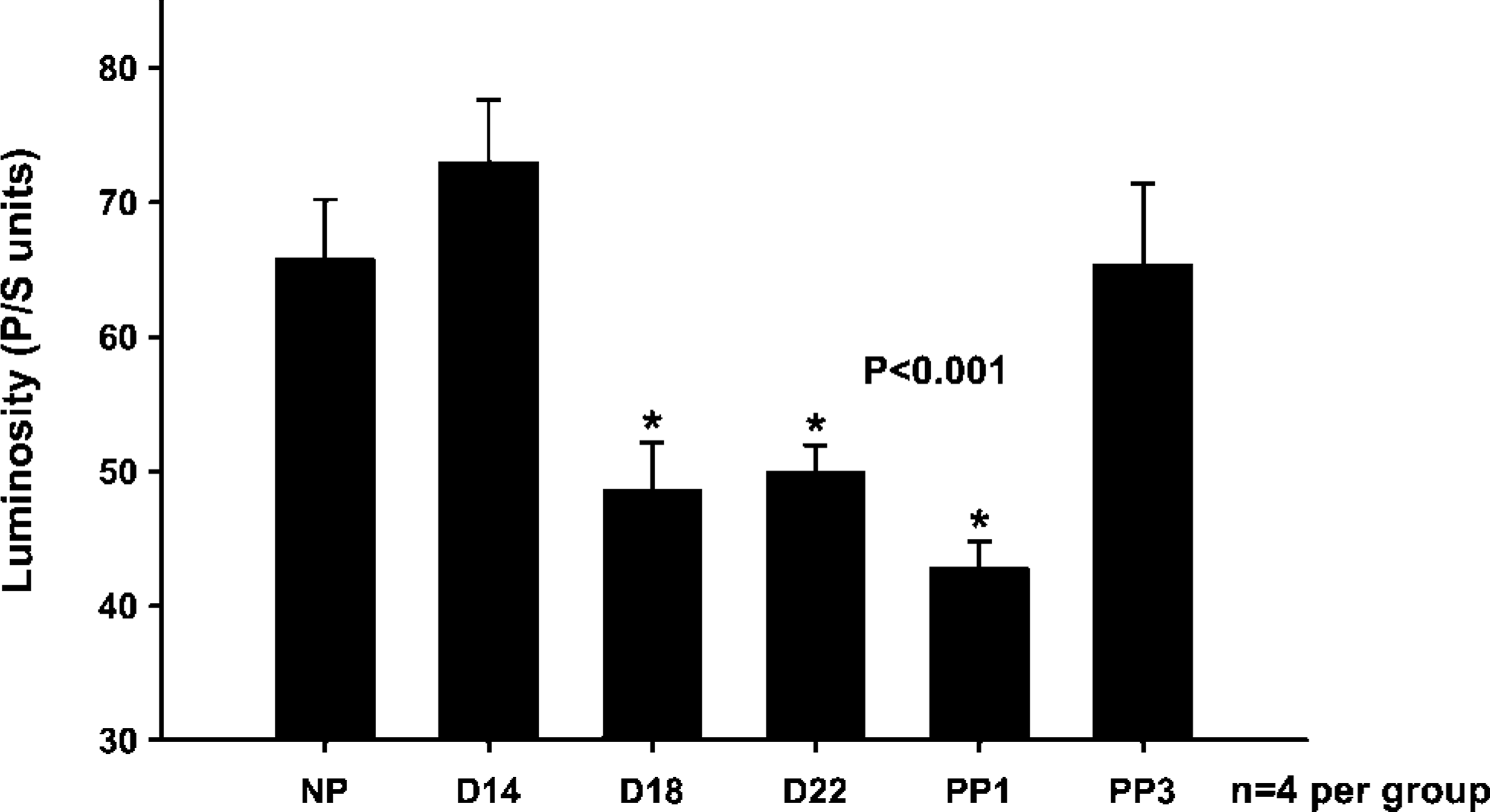
Changes in the rat cervical collagen during the ripening process measured by the picrosirius polarization method. The changes in the collagen are measured based on the luminosity of collagen as pregnancy progressed to term and the cervix ripened. Statistical comparison between the groups was performed using a one-way ANOVA followed by the Student-Newman-Keuls test. ∗Indicates significance of p<0.001. (NP: non-pregnant estrus; D14: day 14; D18: day 18; D22: day 22; PP1: postpartum day 1; PP3: postpartum day 3; n=4 per group).
COX-2. Figures 5B1-5B7 is representative tissues demonstrating the localization sites for the COX-2 enzyme throughout gestation in the rat cervix. Labeling for COX-2 was present in the cervical tissues at all time points examined. The COX-2 enzyme was mainly localized in the cervical smooth muscle (Figure 5B and Figure 6G). Expression of COX-2 was also localized in the vascular smooth muscle (Figure 5B and Figure 6I). Epithelial labeling occurred in the laboring cervix (Figure 5B5 and Figure 6H). The estrous cervix had label for the COX-2 enzyme in the cervical smooth muscle, the vascular smooth muscle, and within the epithelium (Figure 5B1).
Cervical iNOS and COX-2 mRNA Expression during Gestation
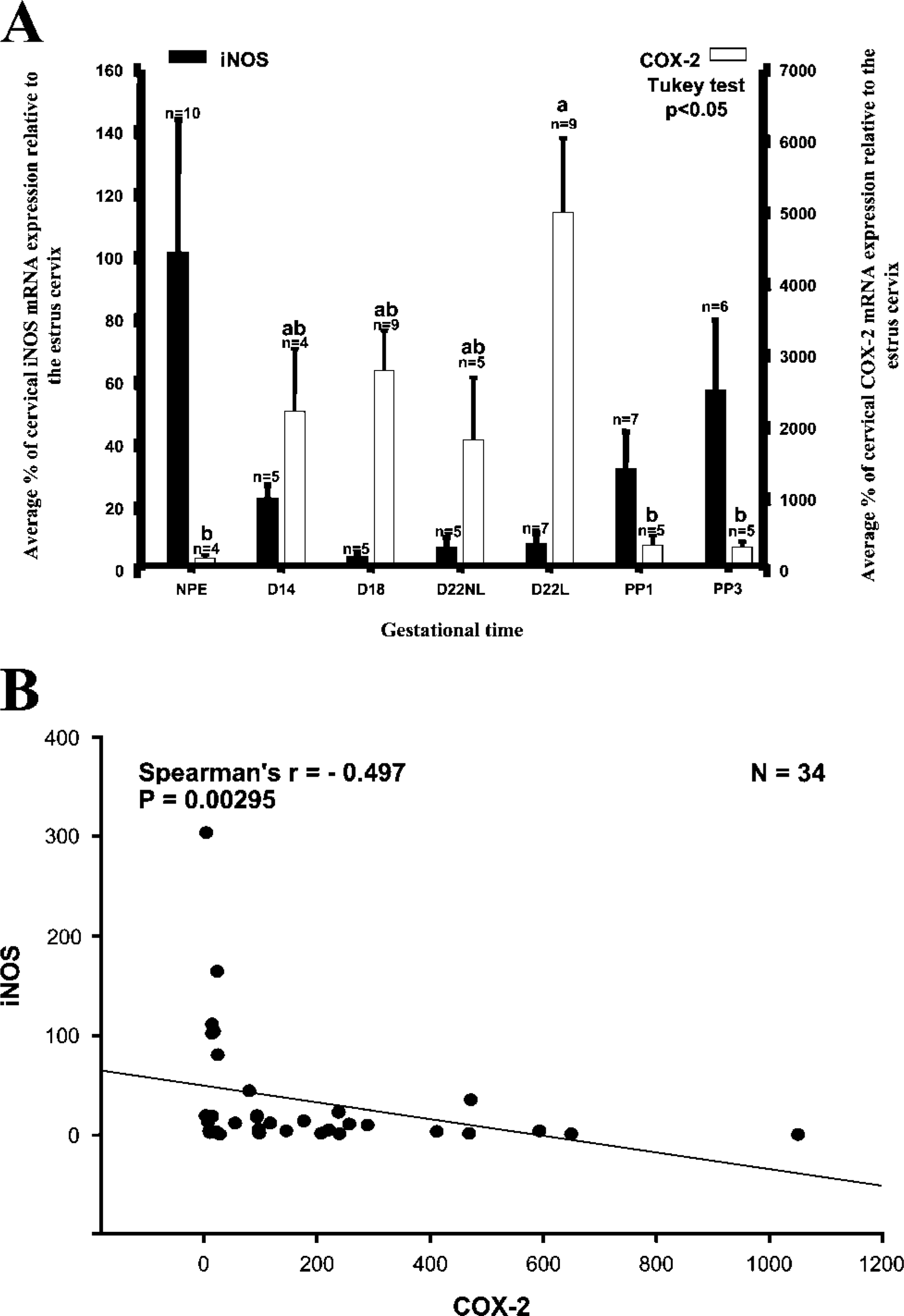
iNOS. The iNOS mRNA expression decreased as pregnancy progressed to term with the highest levels occurring in the nonpregnant and postpartum cervix and the lowest levels occurring at day 18 (Figure 7A), although the differences were not statistically significant.
COX-2. There was about a 50-fold increase in COX-2 mRNA expression in the day 22 laboring cervix compared with estrus, followed by a more than 19-fold decrease in the postpartum day 3 cervix (p<0.05; Figure 7A).
(
iNOS and COX-2. There is a significant inverse relationship between iNOS and COX-2 mRNA expression (Spearman r=-0.497; p=0.00295) (Figure 7B).
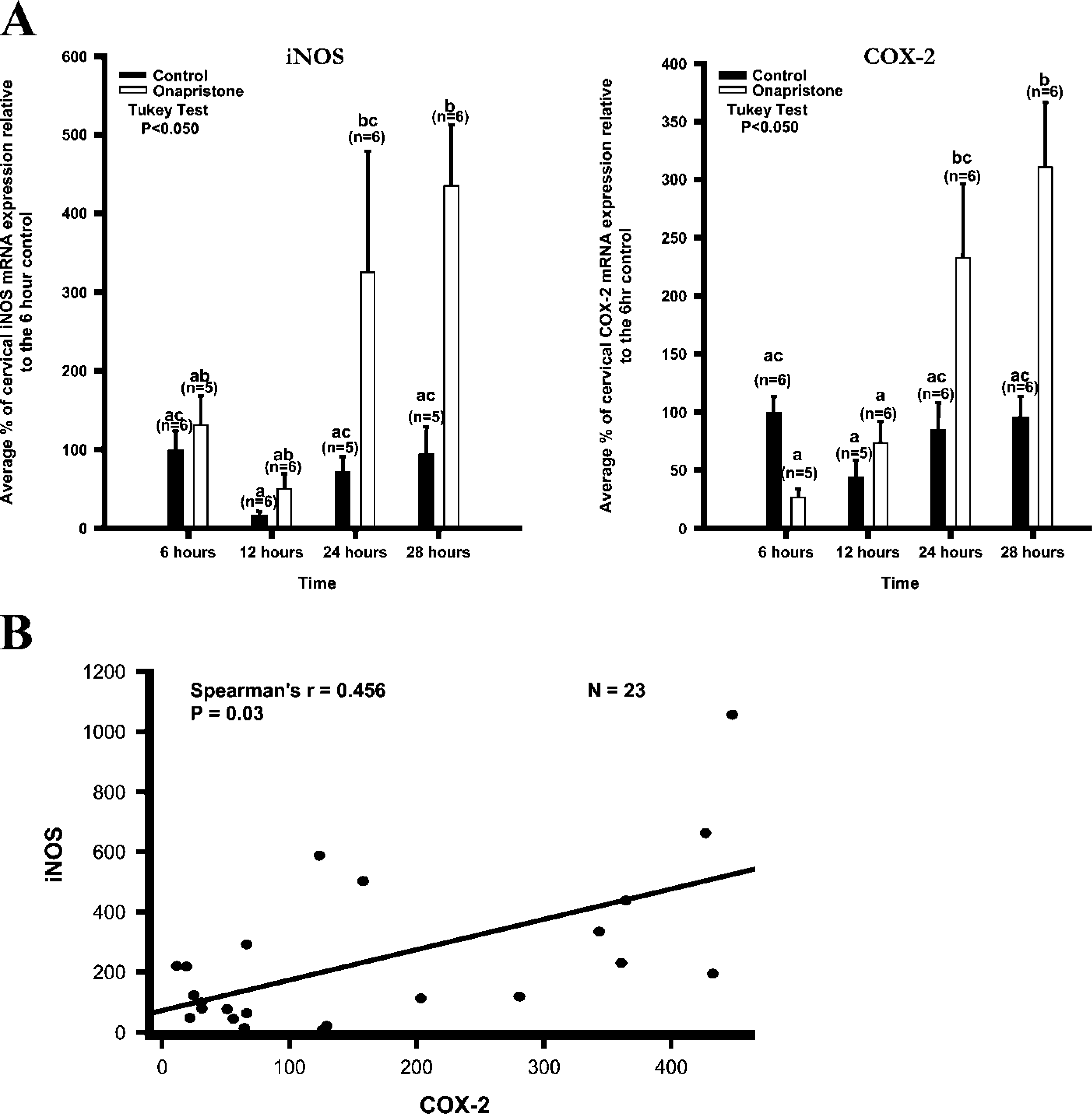
Cervical iNOS and COX-2 mRNA Expression After Induction of Preterm Labor with the Antiprogestin Onapristone (ZK98299)
Onapristone. Treatment with Onapristone stimulated preterm labor, reducing the duration of the pregnancy. The vehicle control rats did not deliver early, whereas the Onapristone treated rats were in labor or delivering preterm 28 hr after the initial treatment on the 18th day of gestation.
iNOS. Expression of the iNOS mRNA gradually increased 6, 12, 24, and 28 hr after Onapristone treatment in the day 17 rat cervix. iNOS mRNA levels peaked 28 hr after treatment with Onapristone (Figure 8A). This peak was significantly different from all the control groups (p<0.05).
(
COX-2. Expression of the COX-2 mRNA gradually increased 6, 12, 24, and 28 hr after Onapristone treatment in the day 17 rat cervix. COX-2 mRNA levels peaked 28 hr after treatment with Onapristone (Figure 8A). This peak was significantly different from all groups except for the 24 hr-treated group (p<0.05).
iNOS and COX-2. A significant correlation between iNOS and COX-2 was demonstrated (Spearman r = 0.456; p=0.03; n=23) (Figure 8B), indicating that as iNOS increased, so did COX-2 after treating with Onapristone; and the increase in iNOS and COX-2 was associated with preterm labor.
Cervical iNOS and COX-2 mRNA Expression After the Prolongation of Pregnancy with Progesterone
Progesterone. Treatment with progesterone inhibited term labor, prolonging the pregnancy. The vehicle control rats delivered at term (day 22), whereas the progesterone treated rats remained pregnant even after 6 days of treatment.
Representative pictures of iNOS (
iNOS. In animals treated with progesterone, iNOS mRNA expression appeared to be reduced and maintained at a low and consistent level (Figure 9). However, the decrease was not significant.
COX-2. In animals treated with progesterone, COX-2 mRNA expression was maintained at a low and consistent level, demonstrating a significant (p=0.036) decrease in the treated vs the nontreated groups. COX-2 mRNA expression was significantly (p<0.001) decreased at day 20 in the treated vs the nontreated group (Figure 9). COX-2 significantly increased in the day 20 nontreated group compared with the day 21 (p<0.001), 22 nonlabor (p=0.001), and postpartum day 2 (p=0.005) nontreated rat cervix. The increase in COX-2 on day 20 of gestation in the cervix may be due to an increase in leukocyte infiltration or epithelial expression.
Cervical iNOS and COX-2 mRNA Expression during Both Term and Preterm Laboring Conditions
At term in the day 22 laboring rat cervix, iNOS mRNA significantly decreased (p=0.022), whereas COX-2 mRNA significantly increased (p= 0.011) (Figure 10A). The expression of iNOS (p= 0.005) and COX-2 (p= 0.004) mRNA was significantly increased during labor under preterm laboring conditions, 28 hr after treatment (Onapristone) compared with the 28-hr control cervix (Figure 10B). During preterm vs term labor, iNOS mRNA was significantly increased (p=0.004); however, there was no difference in the COX-2 mRNA levels (Figure 10C).
(
Discussion
Preterm labor and birth is the leading cause of infant mortality. It is responsible for 75% of all infant mortality and 50% of long-term neurologic disorders (Berkowitz and Papiernik 1993; Creasy 1993; Simpson 1997; Goldenberg and Rouse 1998). Given these statistics, a better understanding of the mechanisms that regulate cervical ripening and parturition under normal (normal term pregnancy) and abnormal (preterm labor and the prolongation of pregnancy) conditions is critically important for our understanding of the pathogenesis of these conditions and possible development of new therapeutic interventions. Treatments that could prolong pregnancy, preventing premature birth, could have a profound effect on fetal mortality and the family unit (Iams 1995).
Our studies examine the relationship between iNOS and COX-2 in the control of cervical ripening and parturition under both normal (normal term pregnancy) and abnormal (preterm labor and the prolongation of pregnancy) conditions. This is accomplished by first characterizing, both visually and quantitatively, the changes taking place in the collagen. Second, is through the localization and characterization of iNOS and COX-2 expression under normal conditions during the different stages of the ripening process and parturition; and third, is by characterizing the changes in the iNOS and COX-2 mRNA expression in the rat cervical tissues during pregnancy under abnormal conditions. Our data suggest that as pregnancy progresses to term, there is a steady decrease in the birefringence and organization of the collagen. Quantitatively, the decrease in the birefringence and the organization of the collagen is associated with a decrease in the luminosity. This indicates that as pregnancy progresses to term there is a progressive decrease in the crosslinked collagen and the long collagen fiber strands. Our data suggest that the mechanisms regulating cervical ripening and parturition may differ under term vs preterm laboring conditions. Specifically, under normal term pregnant conditions, there may not be a relationship between the NO and PG pathways. They may act independently of each other in regulating the iNOS and COX-2 mRNA expression in the control of cervical ripening and parturition. Under normal conditions, the iNOS and COX-2 enzymes were colocalized in the cervical smooth muscle, the vascular smooth muscle, and epithelium at similar times during gestation. The iNOS mRNA levels had a tendency to decrease, whereas COX-2 mRNA levels increased as pregnancy progressed to term, demonstrating an inverse relationship. The data under abnormal conditions suggest that under preterm laboring conditions, there is a relationship between iNOS and COX-2 expression in the control of cervical ripening and parturition. Also, the data suggest that progesterone may play a role early on in the relationship between iNOS and COX-2 in regulating the mRNA expression and parturition. Under abnormal conditions, progesterone prolonged pregnancy and appeared to cause a decrease in iNOS and COX-2 mRNA expression. Onapristone stimulated preterm parturition and produced a parallel increase in iNOS and COX-2 mRNA expression, demonstrating a positive correlation. The data on term vs preterm parturition indicate that PGs are important for parturition. However, NO may play a more important role under preterm labor conditions possibly working through PGs. The data also indicate there may need to be a peak in PGs levels for parturition to occur. This suggests that if the peak occurs, early preterm labor is initiated; if the peak does not occur, the pregnancy is prolonged. At term, under normal laboring conditions (at parturition), iNOS mRNA decreased, whereas COX-2 increased. In contrast, under preterm laboring conditions (at parturition), both iNOS and COX-2 mRNA increased. When comparing term with preterm parturition, there was no change in the COX-2 mRNA expression, but there was a significant increase in iNOS mRNA expression under preterm laboring conditions.
(
Our studies indicate that, under polarized light, the picrosirius red F3BA dye can be used to visualize and measure changes in the collagen during the ripening process and parturition. These changes can be correlated with various treatments or changes in the expression of various enzymes that occur during pregnancy. In our studies, there was a decrease in the organization and birefringence of the cervix as pregnancy progressed to term (Figure 2 and Figure 3). This is comparable to previous studies showing a decrease in cervical resistance and light-induced fluorescence as pregnancy progresses to term and the cervix ripens (Shi et al. 1999). Tissue taken from the fibrous core of the human cervix demonstrated an increase in the distensibility during pregnancy compared with the nonpregnant cervix (Conrad and Ueland 1983). This suggests that as pregnancy progresses to term, the cervix becomes soft and pliable, indicating the importance of collagen and changes in collagen in the ripening of the cervix and parturition (Danforth 1980; Conrad and Ueland 1983; Shi et al. 1999).
Our studies, and other studies, indicate that the ripening of the cervix is a continual process that occurs throughout gestation, preparing the cervix for delivery and a successful end to pregnancy (Conrad and Ueland, 1983; Shi et al., 1999). The human cervix is clinically soft several weeks before parturition, but remains closed with a high tensile strength, possibly because of the length or orientation of the collagen fibers (Yu et al. 1995). Our studies show that in the rigid cervix (Figure 2; estrus, day 14, postpartum 3), collagen is oriented as bundles of long fibers, suggesting a high tensile strength (Vidal et al. 1982; Leppert and Yu 1994; Yu et al. 1995; Shi et al. 1999). This is lost late in gestation when the cervix is already soft (Figure 2; day 18, 22, and postpartum 1 cervix). The long strands of collagen are gone, replaced by aggregates of much shorter and thinner fibers and a decreased birefringence. With a change in the orientation of the collagen, the cervix is now soft, and effacement and dilation can occur (Vidal et al. 1982; Leppert and Yu 1994; Yu et al. 1995). This suggests that the orientation and the length of the collagen fibers are important in maintaining the rigidity of the cervix and play an important role in the ripening process during pregnancy.
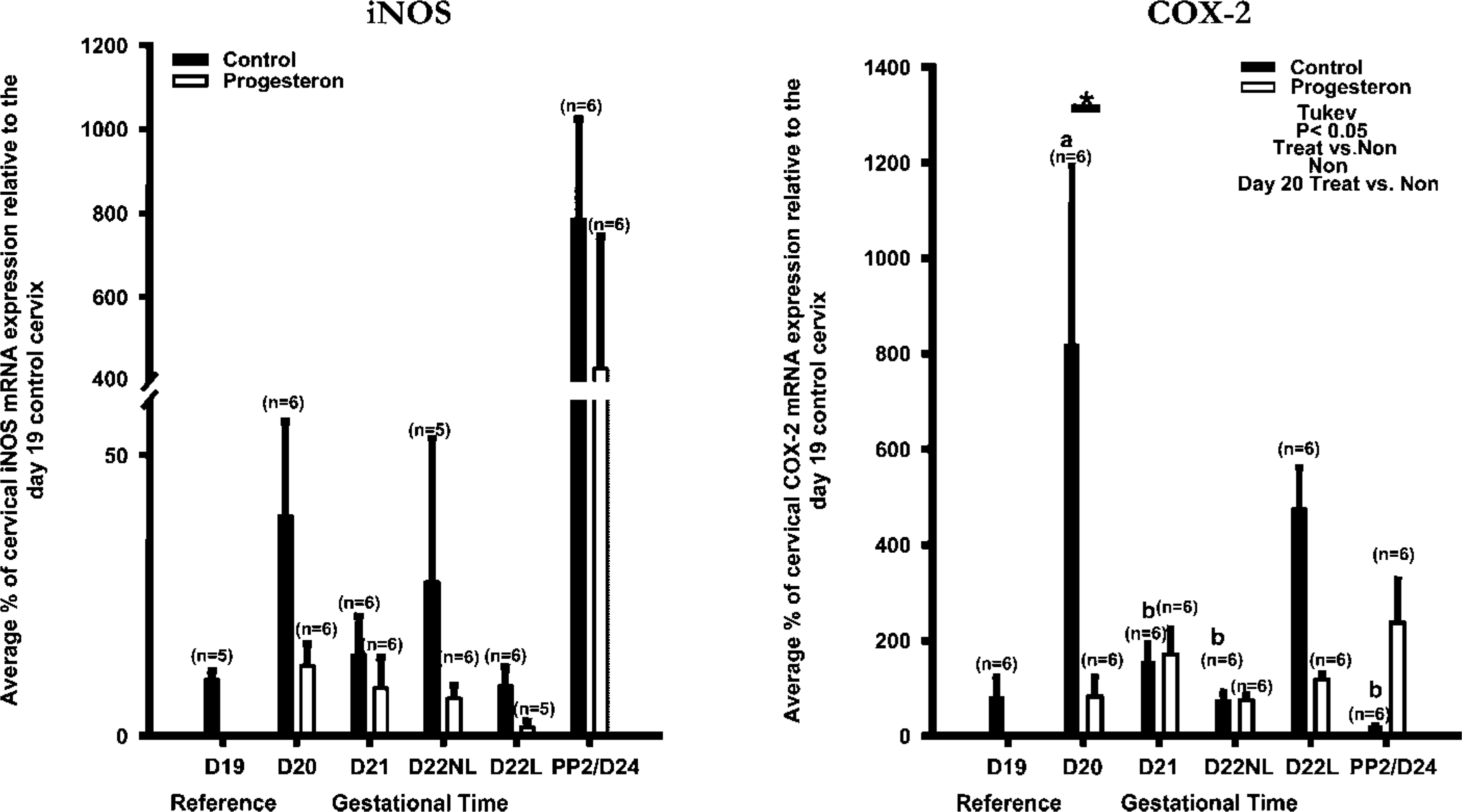
The expression of iNOS and COX-2 mRNA in rat cervical tissues after prolongation of pregnancy with progesterone. The data are expressed as the average % of cervical iNOS or COX-2 mRNA relative to the day 19 control cervix compared with gestational time. Statistical comparison between the groups was performed using a two-way ANOVA followed by a Tukey test for multiple comparisons (p<0.05 is significant). Different letters are an indication of statistical significance (five to six animals per group).
There may be a possible interaction between the NO and PG pathways at the protein or product level in the control of enzyme activity of iNOS and COX-2 during cervical ripening and parturition (Corbett et al. 1993; Salvemini et al. 1993; Tetsuka et al. 1994; Swaisgood et al. 1997; Ledingham et al. 1999; Ekerhovd et al. 2002). Our data show the expression of the iNOS and COX-2 enzymes in similar tissues at similar times during gestation, suggesting a possible interaction between the pathways (Figures 4-6). Buhimschi et al. (1996) and Ledingham et al. (2000) showed the iNOS enzyme to be present in the cervix during pregnancy. Dong et al. (1996) showed the COX-2 enzyme to be present in the cervix during pregnancy. Salvemini et al. (1993) and Ekerhovd et al. (2002) showed a potential interaction between iNOS and COX-2 at the protein and product level. Our data support these studies, indicating a possible interaction between the NO and PG pathways at the protein or product level.
At the mRNA level, iNOS had a tendency to decrease, whereas COX-2 increased, demonstrating an inverse relationship (Figures 7A and 7B) (Puri and Garfield 1982; Dong et al. 1996; Fittkow et al. 2001; Tornblom et al. 2005). Fittkow et al. (2001) showed iNOS levels to be low in guinea pigs during pregnancy. Puri and Garfield (1982) showed an increase in the PG levels. Dong et al. (1996) and Fittkow et al. (2001) showed an increase in the COX-2 level. Our data confirm this.
The decrease in iNOS in our study was not significant. This may be due to variability within the physiologic samples (Figure 7A). Other studies report that iNOS or NO increases rather than decreases in the cervix during labor (Buhimschi et al. 1996; Ali et al. 1997; Ledingham et al. 2000). The difference between our results and previous studies may be explained by the time points examined, because most of these studies examined the later stages of gestation. However, compared with the estrous and postpartum tissues, iNOS expression was low during pregnancy (Figure 7A) (Fittkow et al. 2001). Another explanation may be the differences in the techniques, because our lab now uses a much more sensitive real-time RT-PCR system. In addition, previous studies in rat cervical tissues used Sprague-Dawley rats from Harlan, whereas our studies use rats from Charles River. There may be subtle genetic variability in two outbred rat colonies of the same species (Shi et al. 2000a).
The decrease in iNOS and the increase in COX-2 are thought to play a role in regulating matrix metallopeptidase (MMP)-8 and MMP-9 expression in the continued breakdown of the collagen during the ripening process (Winkler et al. 1999). NO is thought to downregulate or inhibit MMP-8 and MMP-9 (Upchurch et al. 2001). The decrease in iNOS expression or NO production may be required to allow an increase in MMP-8 and MMP-9 in the later stages of gestation. The increase in COX-2 expression or PG production may be required for parturition and to allow an increase in IL-8, a chemotactic agent that attracts neutrophils that are known to express MMP-8 and MMP-9 (Chwalisz et al. 1994; El Maradny et al. 1994; Winkler et al. 1999; Elliott et al. 2001; Watari et al. 2003).
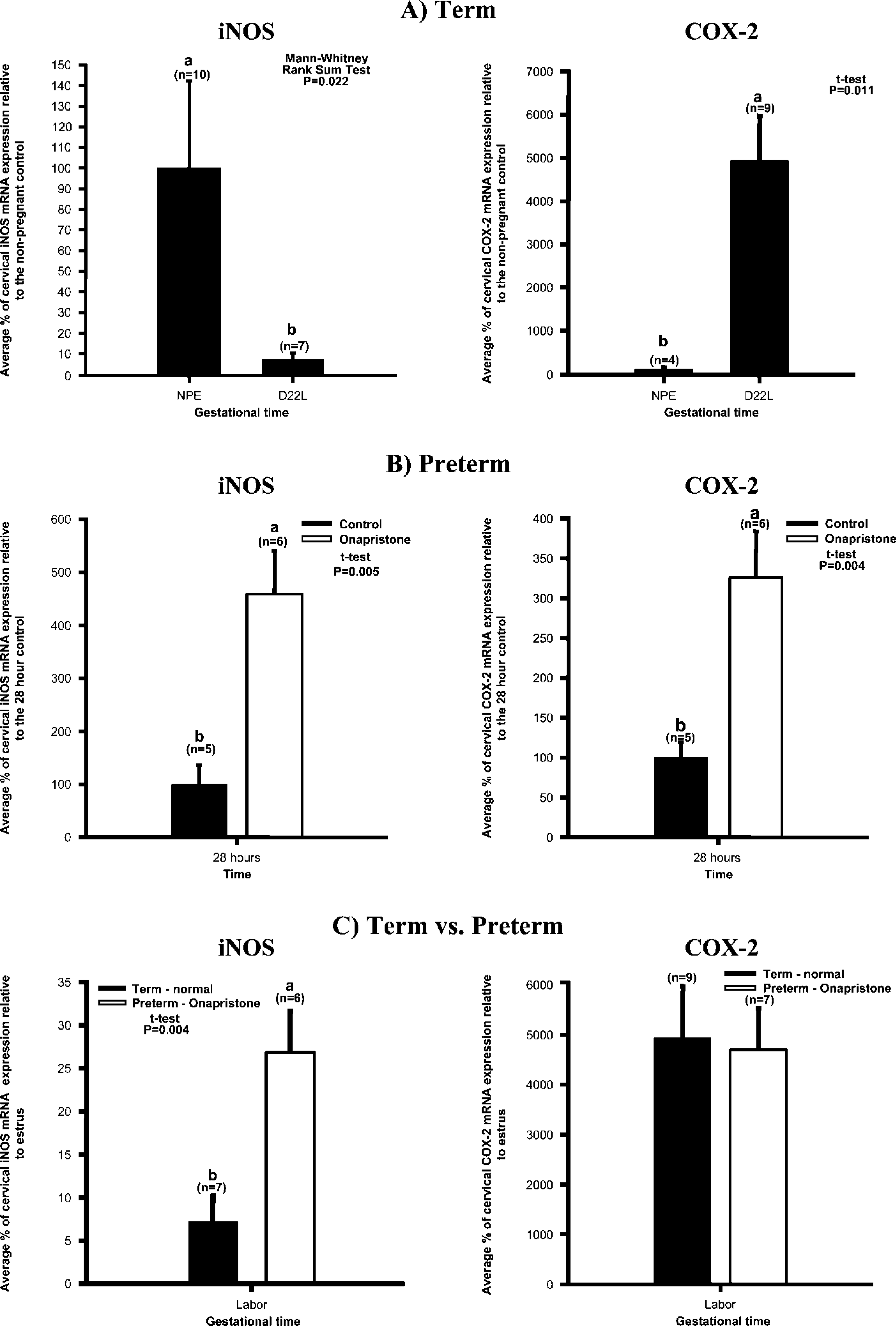
A comparison between iNOS and COX-2 mRNA expression during term and preterm laboring conditions. (
The iNOS and COX-2 mRNA expression appears to increase (Figure 7A; iNOS: at estrus and in the postpartum tissues; COX-2: at labor) primarily at times during pregnancy when epithelial staining for the iNOS and COX-2 enzyme (Figures 4-6) occurs, suggesting that an increase in iNOS and COX-2 may be due to increased epithelial expression (Tschugguel et al. 1999; Tornblom et al. 2005).
Onapristone is shown to ripen the cervix in many animals (Elger et al. 1986; Frydman et al. 1988; Hegele-Hartung et al. 1989; Chwalisz et al. 1991; Chwalisz 1994). In our studies, Onapristone stimulates preterm parturition and produces a parallel increase in the iNOS and COX-2 mRNA expression, demonstrating a positive statistical correlation and suggesting a potential relationship/interaction between the NO and PG pathways under preterm laboring conditions (Figures 8A and 8B) (Nathanielsz 1978a,b; Thorburn and Challis 1979; Puri and Garfield 1982; Buhimschi et al. 1996; Dong et al. 1996; Yallampalli et al. 1996; Ali et al. 1997; Selles et al. 2002; Tornblom et al. 2005). Hegele-Hartung et al. (1989) show that after being treated with an antiprogestin, the collagen fibers become scattered and dissociate from one another. This is associated with stromal edema and an increase in leukocyte infiltration into the cervix. Chwalisz et al. (1997) show a decrease in the cervical resistance after being treated with antiprogestin, NO, or PGs. Ali et al. (1997) show an increase in iNOS expression after treating with an antiprogestin. These studies, and ours, suggest that progesterone plays a role in regulating the ripening of the cervix and parturition, possibly acting through iNOS and COX-2 expression. It also suggests that cervical incompetence or preterm labor may be due to either deficiencies or abnormalities in the PR or a decrease in the progesterone.
In our study, progesterone prolongs pregnancy and decreases the iNOS and COX-2 mRNA expression, maintaining them at a relatively low and consistent level (Figure 9). The decrease in iNOS is not significant, possibly because of the iNOS levels already being low (Figure 7A). Buhimschi et al. (1996) showed a decrease in NO production after being treated with progesterone. This would suggest that progesterone plays a role in the ripening of the cervix and parturition, possibly acting through an increase in NO and PG production. Also, failure of the cervix to ripen or prolongation of the pregnancy may be due to an increase or overexpression of progesterone, its receptors, or a decrease in the NO and PG production (Puri and Garfield 1982; Buhimschi et al. 1996; Dong and Yallampalli 1996; Ali et al. 1997).
There is a peak in the COX-2 mRNA expression that occurs under both term and preterm laboring conditions. The iNOS mRNA expression is only increased under preterm laboring conditions. This suggests that PGs are important for parturition, but that NO may play a more important role under preterm labor conditions—possibly acting through PGs. It also suggests that PGs may need to increase or reach a certain level for parturition to occur (Figure 10) (Chwalisz et al. 1991; Radestad and Bygdeman 1993). Our data suggest that a shift in this peak to an early time point may stimulate preterm parturition (Figure 8A and Figure 10), or if the peak does not occur, pregnancy may be prolonged (Figure 9).
Fibronectin (a determination of the fibronectin protein in the vaginal fluid) and cervical changes are used in predicting preterm labor. However, they have been found to work mainly in high-risk situations. This increase or peak in PGs has the potential to be used as a method, along with fibronectin and changes in the cervix, for predicting preterm labor in both high- and low-risk situations early in pregnancy.
In summary, the goals of our studies were to better understand the relationship/interaction between iNOS and COX-2 in the control of cervical ripening and parturition. A better understanding of how these pathways are regulated may have a significant impact on the development of the diagnostic and therapeutic modalities in treating medical conditions involving pregnancy and birth. The main observations of our studies are that the picrosirius polarization staining method can be used to visualize and measure changes in the organization of the collagen during the ripening process and parturition. The method can also be used to correlate changes in the enzymes with the breakdown of the collagen in the cervix. There is a decrease in the organization and birefringence of the collagen as pregnancy progresses to term in association with the iNOS and COX-2 enzymes being localized within the cervix throughout gestation. This suggests a decrease in the crosslinked collagen and the long collagen fibers. It also suggests that the orientation of the collagen changes, becoming dispersed and disorganized as the cervix softens and that NO and PGs may play an important role in this process. Our data show that there may be an interaction between the NO and PG pathways in the control of cervical ripening and parturition. Progesterone may play a role in regulating the relationship or interaction between the NO and PG pathways early in the ripening process and parturition. Early in the ripening process, when progesterone levels are high, it is thought to decrease and maintain the iNOS and COX-2 levels, the NO and PGs levels at relatively low and consistent level preventing and interaction between the two pathways. However, when progesterone levels decrease in the later stages of gestation, it is thought to cause an increase in the PG levels and, possibly, the NO levels, allowing for an interaction between the two pathways in the control of the ripening process and parturition. This would suggest that under normal conditions, this interaction may not occur, at least in the regulation of the mRNA expression, because an increase in iNOS is not observed. The two pathways may act independently of each other in the ripening of the cervix and parturition under normal conditions, at least at the mRNA level. In contrast, under preterm laboring conditions, there may be an interaction, because an increase in both iNOS and COX-2 is observed in the regulation of the mRNA expression. The NO and PG pathways may interact in regulating the ripening of the cervix and parturition under preterm laboring conditions. Another important novel finding from these studies is that PGs may be involved in regulating parturition under both normal and abnormal conditions. NO may only be involved in regulating parturition under abnormal conditions, such as with preterm labor, possible acting through PGs. We are further defining the role of PGs in the ripening process and parturition, suggesting that PGs are increased under both term and preterm laboring conditions, stimulating parturition; and, when downregulated, the pregnancy is prolonged. These are new and unique findings that represent a significant advance in the literature in understanding the relationship between iNOS and COX-2 in the control of cervical ripening and parturition. In addition, it was previously thought that the mechanisms regulating the ripening processes and parturition, under both normal term and preterm laboring conditions, were the same, but that under preterm laboring conditions, the process was sped up. We demonstrate an inverse relationship between iNOS and COX-2 under normal term pregnant conditions and a direct relationship under preterm laboring conditions. This suggests that the mechanisms regulating the ripening process and parturition under normal term and preterm laboring conditions may differ, but the outcome is the same (an increase in PGs and the fetus being delivered). Finally, the failure of the cervix to ripen or the prolongation of pregnancy or cervical incompetence or preterm labor may be due to problems associated with progesterone or the regulation of iNOS and COX-2 expression (NO and PG production).
Footnotes
Acknowledgements
This study was supported by National Institutes of Health Grant #5 F31 HD-008739.
We express our gratitude to Yakov Pesah, PhD, of Baylor College of Medicine for his help with this work.