
Abstract
We investigated nitric oxide (NO) production and the presence of nitric oxide synthase (NOS) in the mammary gland by use of an organ culture system of rat mammary glands. Mammary glands were excised from the inguinal parts of female Wistar-MS rats primed by implantation with pellets of 17β-estradiol and progesterone and were diced into approximately 3-mm cubes. Three of these cubes were cultured with 2 ml of 10% FCS/ DMEM plus carboxy-PTIO (an NO scavenger, 100μM) in the presence or absence of LPS (0.5μg/ml) for 2 days. The amount of NO produced spontaneously by the cultured mammary glands was relatively minute at the end of the 2-day culture period, and the NO production was significantly enhanced by the presence of LPS. This enhancement of NO production was completely eliminated by addition of hydrocortisone (3μM), an inhibitor of inducible NOS (iNOS), to the incubation medium. Immunoblot analyses with specific antisera against NOS isoforms such as iNOS, endothelial NOS (eNOS), and brain NOS (bNOS) showed immunoreactive bands of iNOS (122 ± 2 kD) and eNOS (152 ± 3 kD) in extracts prepared from the mammary glands in the culture without LPS. The immunoreactive band of iNOS was highly intense after the treatment of mammary glands with LPS, whereas the corresponding eNOS immunoreactive band was faded. The immunohistochemical study of anti-iNOS antiserum on frozen sections of the cultured mammary glands showed that an immunoreactive substance with the antiserum was localized to the basal layer (composed of myoepithelial cells of alveoli and lactiferous ducts) of the mammary epithelia and to the endothelium of blood vessels that penetrated into the interstitium of the mammary glands. Histochemical staining for NADPH-diaphorase activity, which is identical to NOS, showed localization similar to that of iNOS in the mammary glands. Similar observations were noted in the immunohistochemistry of eNOS. In contrast, the immunoreactive signal with the bNOS antiserum was barely detected in the epithelial parts of alveoli and lactiferous ducts of the mammary glands. These observations demonstrate that three isoforms of NOS are present not only in the endothelium of blood vessels but also in the parenchymal cells (the glandular epithelium) of the rat mammary gland, such as epithelial cells and myoepithelial cells, and suggest that NO may have functional roles in the physiology of the mammary glands.
N
NOS has at least three distinct isoforms, including the neuronal (nNOS), brain (bNOS or NOS1), inducible (iNOS or NOS2), and endothelial (eNOS or NOS3) types (Mayer 1995; Wang and Marsden 1995). Although these were originally purified from the cerebellum (Bredt and Snyder 1990), cytokine-induced macrophages (Hevel et al. 1991; Stuehr et al. 1991; Yui et al. 1991), and the vascular endothelium (Förstermann et al. 1991; Pollock et al. 1991) and designated nNOS, iNOS and eNOS, respectively, the isoforms are now known to distribute across a wide spectrum of cell types and tissues. These include the adrenal gland (Afework et al. 1994, 1996; Tanaka and Chiba 1996), the digestive organs (Xue et al. 1994; Burrell et al. 1996; Chen et al. 1996), kidney (Bachmann et al. 1995; Fischer et al. 1995; Schwartz et al. 1997), bladder (Ehrén et al. 1995), liver (Bucher et al. 1997), pancreas (Umehara et al. 1997; Burrell et al. 1996), the female reproductive organs (Myatt et al. 1993; Ben-Shlomo et al. 1994; Huang et al. 1995; Chatterjee et al. 1996; Zackrisson et al. 1996), the male reproductive tract and its accessory organs (Burnett et al. 1995; Stéphan et al. 1995; Zini et al. 1996; Bloch et al. 1997), and many other tissues and organs. In addition to these organs and tissues, NOS isoforms were identified in human breast cancer and cancer cell lines (Thomsen et al. 1995; Zeillinger et al. 1996; Dueñas-Gonzalez et al. 1997). The presence and distribution of NOS isoforms in the normal mammary gland, in contrast, are not yet known. The aim of the present study was to investigate the presence and distribution of the NO-generating enzymes (NOS) in the rat mammary gland by using an organ culture system and immunobiological and immunohistochemical means. The possible functional roles of NO in the mammary gland are also discussed.
Materials and Methods
Materials and Animals
Reagents and chemicals were purchased from Sigma Chemical (St Louis, MO) unless stated otherwise. All animals used in the present study were treated and handled according to the “Recommendations for the Handling of Laboratory Animals for Biomedical Research” complied by the Committee of the Safety and Handling Regulations for Laboratory Animal Experiments in our Institute.
Organ Culture of Rat Mammary Glands
Female Wistar-MS rats (8-week-old) bred at Nippon SLC (Hamamatsu, Japan) were primed by implantation with pellets of 17β-estradiol (0.5 mg/3-week-release type; Innovative Research of America, Toledo, OH) and progesterone (35 mg/3-week-release type, Innovative Research). After 3 weeks of priming the rats were sacrificed by CO2 asphyxiation and the inguinal mammary glands were excised aseptically for organ culture. The isolated mammary glands were diced into approximately 3-mm cubes, and three of these cubes were cultured in a well of 24-multiwell plate containing 2 ml of 10% fetal calf serum (FCS)/Dulbecco's modified Eagle's medium (DMEM) supplemented with antibiotics (100 U/ml penicillin, 100μg/ml streptomycin) and an antimycotic (0.25μg/ml amphotericin B) in a mixture of 5% CO2/95% air at 37C for 2 days. The medium was then replaced with 10% FCS/DMEM plus 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazole-1-oxyl 3-oxide (carboxy-PTIO, 100μM; Dojindo Laboratories, Kumamoto, Japan) in the presence or absence of bacterial lipopolysaccharide (LPS, 0.5μg/ml), and the culture was maintained for another 2 days. At the end of the culture, the conditioned media were collected for determination of the nitrite concentration as described below, and the cultured mammary glands were further processed for the preparation of mammary gland homogenates and frozen thin sections as described below.
Determination of Nitrite
NO produced and secreted by mammary glands into the culture medium was estimated by measuring the nitrite (NO2 -) converted from NO with a Griess reagent mixture. The nitrite concentrations in conditioned media were determined immediately after the termination of the culture by a modification of a previously described colorimetric method (Green et al. 1982; Ben-Shlomo et al. 1994). Briefly, the Griess reagent mixture was freshly prepared by mixing equal volumes of stock Solution A (10% sulfanilamide, 40% phosphoric acid) and stock Solution B [1% N-(1-naphthyl) ethylenediamine dihydrochloride] before the measurement of nitrite. One part of the reagent mixture was transferred into 7 parts of the conditioned media appropriately diluted with PBS, and absorbance at either 545 nm or 570 nm was determined in a spectrophotometer. The nitrite concentration was then estimated from a standard curve that was simultaneously prepared for NaNO2 in PBS. The slope of the standard curve was essentially the same whether DMEM or PBS was used as a diluent.
Preparation of Mammary Gland Homogenates and Immunoblot Analysis of Nitric Oxide Synthase Isoforms
The mammary glands cultured with or without LPS were minced and homogenized in ice-chilled 5 mM Tris-HCl buffer (pH 7.5) containing 0.25 M sucrose, 5 mM EGTA, and inhibitors (1 M phenylmethylsulfonyl fluoride, 2 mM sodium vanadate, 10μg/ml aprotinin, 5μg/ml leupeptin) (van Haren et al. 1992; Onoda and Djakiew 1993a,b). The homogenates were reconstituted in reducing sample buffer and loaded into the minigel system for SDS-PAGE. Subsequently, the separated proteins were electrotransferred to a nitrocellulose membrane (Onoda et al. 1991). The membrane was then soaked in 5% nonfat milk in 20 mM Tris-HCl-buffered saline containing 500 mM sodium chloride (TBS, pH 7.5) for 2 hr to block nonspecific immunoreaction, rinsed twice for 10 min in TBS containing 0.05% Tween 20 (TTBS), and reacted with either anti-iNOS antiserum (Affinity Bioreagents; Golden, CO), anti-eNOS antiserum (Affinity Bioreagents), anti-bNOS antiserum (Affinity Bioreagents), or normal rabbit serum (NRS, 1:1000-2000 dilution) in TTBS containing 1% gelatin (Bio-Rad Labs; Richmond, CA) overnight. The membrane was washed in TTBS twice for 10 min, reacted with alkaline phosphatase-conjugated goat anti-rabbit IgG (1:3000 dilution) in TTBS/gelatin for 1 hr, and rinsed in TTBS twice and TBS once. The immunoreactivity was visualized by the following color development reactions (Onoda and Djakiew 1993a, b). For the alkaline phosphatase reaction, 5% nitroblue tetrazolium (NBT) in 70% dimethylformamide and 5% 5-bromo-4-chloro-3-indolyl phosphate (BCIP) in dimethylformamide were freshly prepared. Both NBT and BCIP reagents were then mixed with 100 mM Tris-HCl (pH 9.5) containing 100 mM sodium chloride and 5 mM magnesium chloride (alkaline phosphatase reaction buffer, APB) to a final concentration of 0.033% and 0.017%, respectively. Before color development, the protein-bound nitrocellulose membrane was washed in APB for 10 min and then the immunoreactive bands were visualized in APB containing NBT and BCIP. Color development was terminated by replacement of the reaction mixture with distilled water. The molecular weight of the immunoreactive bands was estimated from plots of molecular weight vs relative mobility of rainbow marker standard proteins (Amersham; Arlington Heights, IL), which were run simultaneously with the sample proteins. Anti-rat transferrin antiserum (Cappel Research Products; Durham, NC) and anti-rat caseins antiserum (Onoda and Inano 1997) were also used to confirm the presence of typical proteins produced by epithelial cells of the mammary glands.
Immunohistochemistry of Nitric Oxide Synthase Isoforms
The mammary glands incubated as mentioned above were cut into small cubes, fixed in 10% neutral buffered formalin at 4C, usually for about 20 hr, immersed in ascending grades of sucrose-PBS, and brought finally to 20% sucrose-5% glycerol-PBS. The fixed tissues then were embedded in Tissue-Tek OCT compound (Sakura Finetechnical; Tokyo, Japan) and frozen in hexane at −60C. Frozen sections (12μm thick) were prepared in a Jung MC1900 cryostat (Leica; Nussloch, Germany), placed on gelatin-chromium (III)-potassium sulfate-coated slide glasses, and allowed to cool air-dry for a half hour. The sections were rehydrated in PBS before immunostaining. Streptavidin-biotin peroxidase immunostaining was carried out using Histofine SAB-PO kits (Nichirei; Tokyo, Japan) as described previously (Inano et al. 1995; Onoda and Inano 1997). Endogenous peroxidase activity was inactivated by 3% hydrogen peroxide for 15 min at room temperature (RT). The sections were washed twice in PBS for 5 min, then blocked by 10% normal goat serum for 30 min. Next, sections were incubated with either anti- NOS antisera or NRS (1:2000-3000 dilution) at 4C overnight and washed twice in PBS. The sections were treated with biotinylated second antibody (anti-rabbit IgG; goat) for 30 min at RT, washed twice in PBS, then incubated with streptavidin-peroxidase conjugate for 30 min and washed twice in PBS. Antibody localization on the specimens was visualized by the substrate-chromogen mixture [0.61 M Tris-HCl buffer (pH7.4) containing 0.05% 3,3′-diaminobenzidine tetrahydrochloride and 0.01% hydrogen peroxide], and color development was stopped by replacement of the reaction mixture with distilled water. The sections were counterstained with hematoxylin, dehydrated, and mounted with a mounting reagent. Immunostainings with anti-rat transferrin and anti-rat caseins were also carried out to validate the location of the mammary gland epithelium. Photomicrographs were taken under an Olympus BX50 microscope with an automatic camera.
NADPH-diaphorase Staining
The NADPH-diaphorase staining was performed according to the method described by Scherer-Singler (1983) with slight modification. Briefly, fixed frozen thin sections prepared as described above were rehydrated in PBS and incubated with a mixture of 1 mM NADPH (b) and 0.2 mM NBT in 50 mM Tris-HCl (pH 7.4) at 37C for 60 min. In the negative control sections, the NADPH (b) was omitted from the reaction mixture. After the staining, the sections were rinsed in PBS, counterstained in eosin, dehydrated, and mounted with a mounting reagent.
Statistical Analysis
Data are expressed as mean ± SE, and statistical comparisons among control and other groups were performed using ANOVA with StatView-J4.5 software (Abacus Concepts; Berkeley, CA) for Fisher's PLSD of multiple comparisons test. The level of significance was defined as p<0.05.
Results
NO Production by Cultured Mammary Glands
The amount of NO produced spontaneously by mammary glands in culture was relatively minute for the 2-day culture period, whereas the NO concentration in the conditioned media was significantly increased (by almost fivefold of the control) by the addition of LPS (0.5μg/ml) to the culture system (Table 1). The secretion of NO from mammary glands cultured with LPS increased in a dose-dependent manner up to a concentration of 0.5μg/ml, and was slightly inhibited at higher concentrations of LPS up to 2μg/ml (Figure 1). This enhancement of NO secretion by the mammary glands in culture with LPS was completely eliminated by addition of 3μM hydrocortisone to the culture (Table 1). Hydrocortisone and corticosterone inhibited NO production/secretion into the culture medium from mammary glands with LPS stimulation in a dosedependent manner, and the median inhibition doses (ID <50) of hydrocortisone and corticosterone were approximately 40 nM and 150 nM, respectively (Figure 2). These findings suggest that the cultured rat mammary gland contains at least a certain cell population that responds to LPS stimulation and that this cell population produces NO.
Nitric oxide production by cultured rat mammary glands a
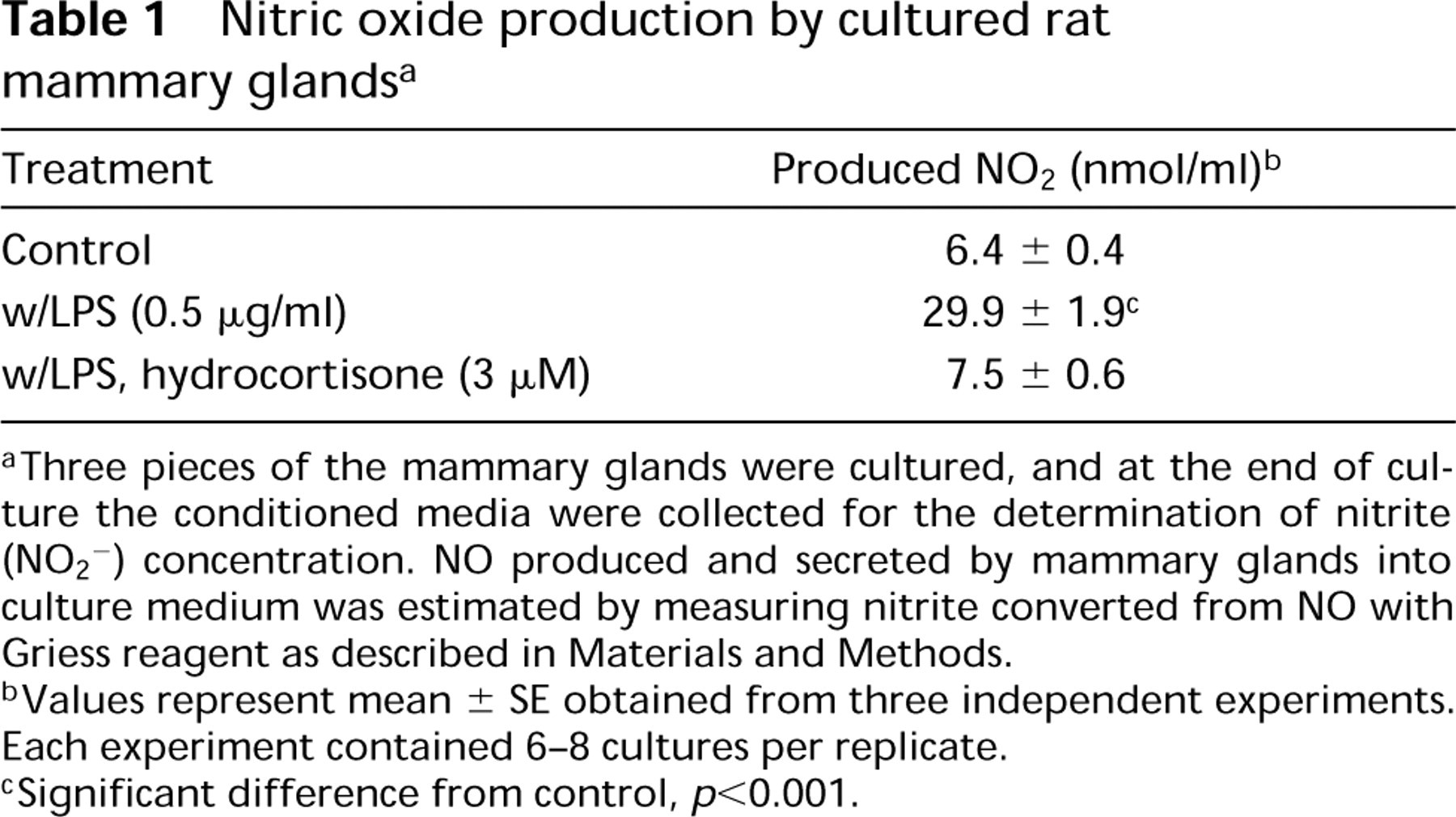
Three pieces of the mammary glands were cultured, and at the end of culture the conditioned media were collected for the determination of nitrite (NO2 2) concentration. NO produced and secreted by mammary glands into culture medium was estimated by measuring nitrite converted from NO with Griess reagent as described in Materials and Methods.
Values represent mean ± SE obtained from three independent experiments. Each experiment contained 6-8 cultures per replicate.
Significant difference from control, p <0.001.
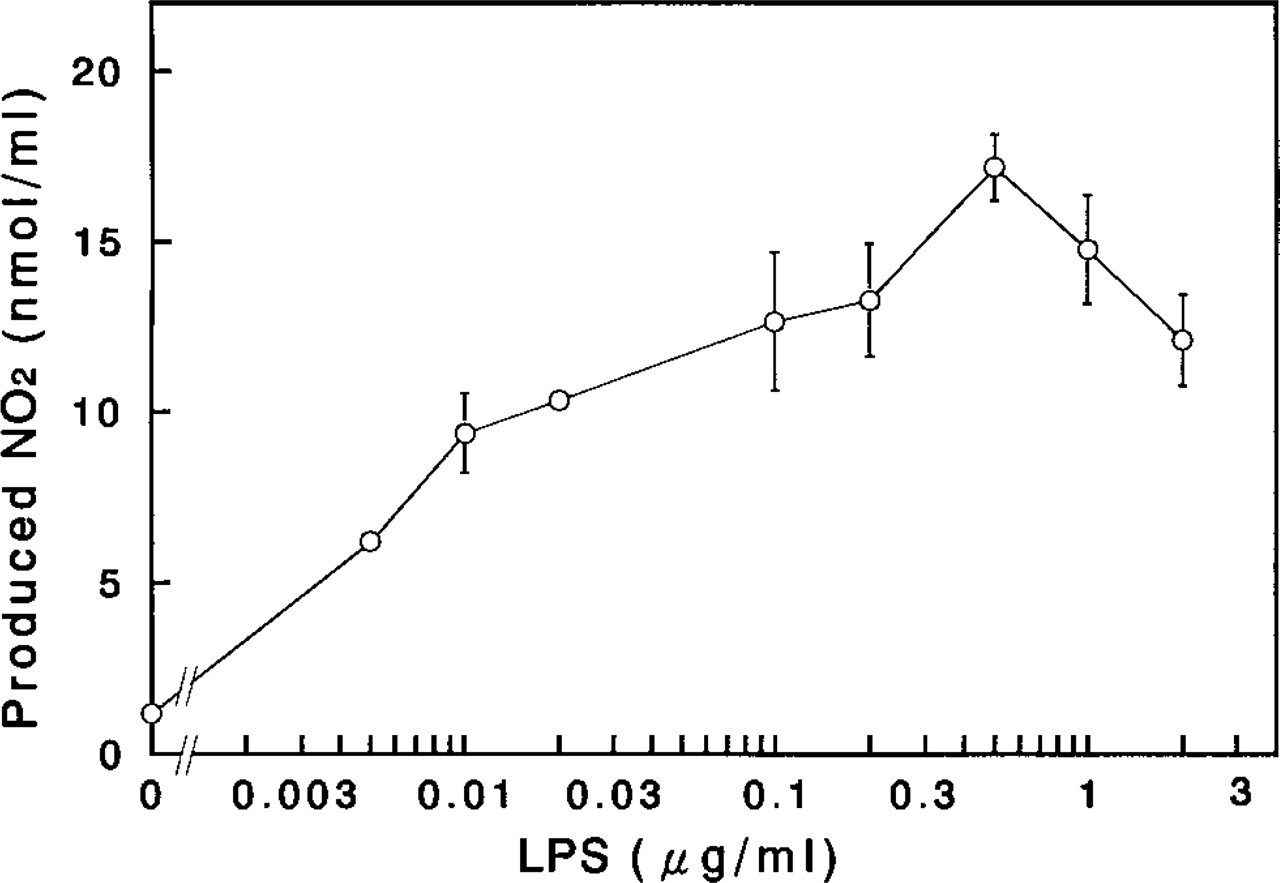
Enhancement of NO production in the cultured rat mammary glands by lipopolysaccharide (LPS). Three pieces of the mammary glands were cultured with the indicated doses of LPS for 2 days and the conditioned media were collected for the determination of the nitrite (NO2-) concentration as described in Materials and Methods. Each point and vertical bar represents the mean ± SE, respectively, obtained from three independent experiments. Each experiment contained three cultures per replicate.
Determination of NOS Isoforms in Mammary Gland Extracts by Immunoblot Analysis
The NOS isoforms that immunoreacted with antiiNOS and anti-eNOS antisera were detected, respectively, in the mammary gland extracts obtained at the terminus of the organ culture of mammary glands (Figures 3 and 4). These immunoreactive bands of iNOS and eNOS had apparent molecular weights of 122 ± 2 kD and 152 ± 3 kD, respectively. In addition to this eNOS immunoreactive band (152 kD), in some cases a few other minor immunoreactive bands with molecular weights of approximately 97 kD, 78 kD, and 46 kD were detected by the anti-eNOS antibody (data not shown). These molecular species may be immunoreactive degradation products of eNOS, which were produced during the preparation of mammary gland extract owing to the presence of certain proteases (Zini et al. 1996). The quantity of immunoreactive band with anti-iNOS antiserum was apparently increased in the mammary gland extract treated with LPS during the culture period (Figure 3), and this increase in iNOS expression was well reflected in the enhancement of NO concentration in the conditioned medium (Table 1). The eNOS expression was clearly diminished in the mammary gland extract after the treatment with LPS (Figure 4), although the gross production of NO was significantly enhanced in the culture with LPS treatment (Table 1). Hydrocortisone (3μM) obviously suppressed not only the iNOS expression but also that of eNOS in the mammary glands cultured with LPS (Figures 3 and 4). Unfortunately, the immunoreaction to the bNOS with anti-bNOS antiserum was inconclusive in the mammary gland extracts obtained in all culture conditions (data not shown).
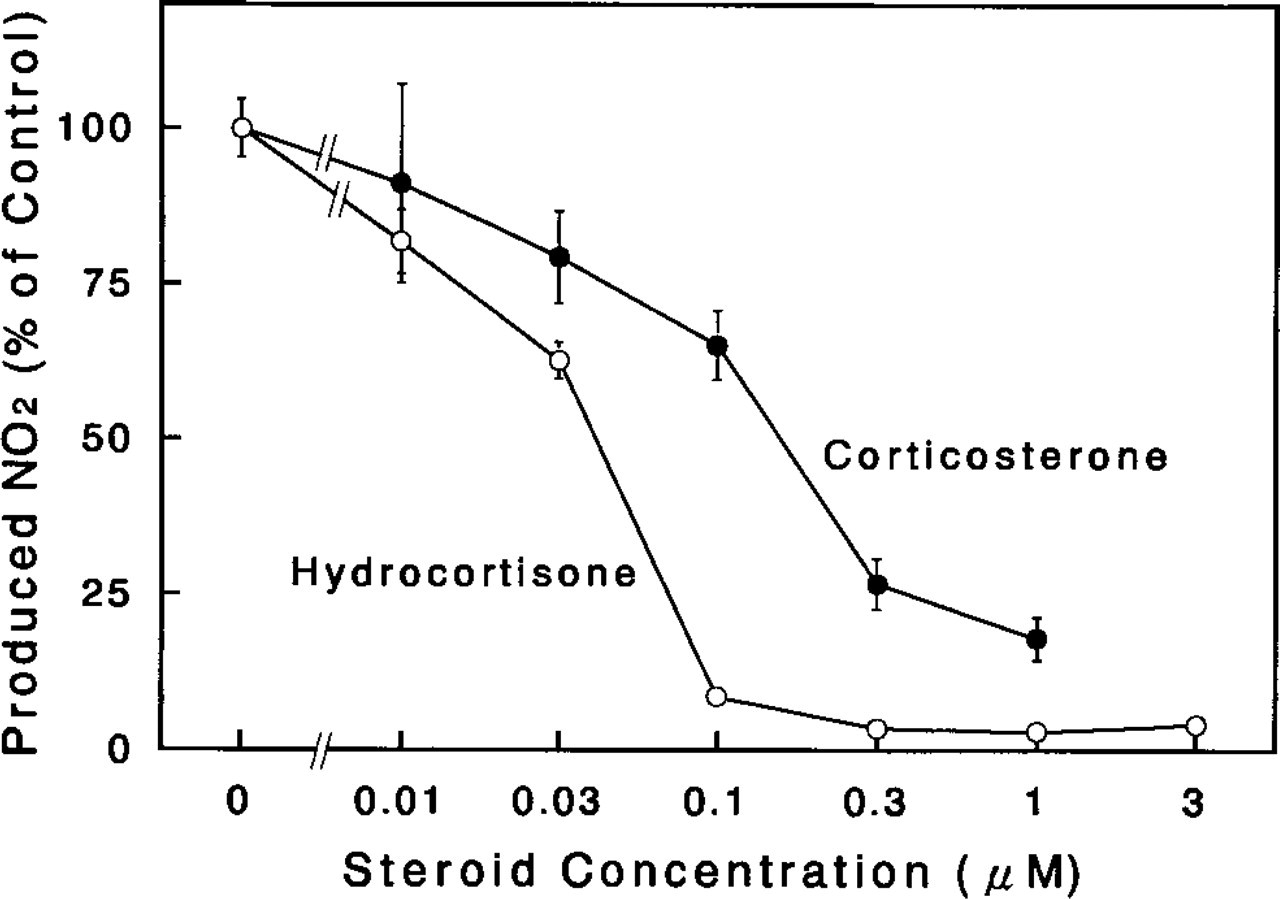
Effect of glucocorticoids on NO production by cultured rat mammary glands. Three pieces of the mammary glands were incubated for 2 days with LPS (0.5 μg/ml) in the presence of the indicated doses of steroids (hydrocortisone, open circles; corticosterone, solid circles), and the conditioned media were collected for the determination of nitrite (NO2-) concentration as described in Materials and Methods. Values presented are the percent of the control culture values (incubated without steroid). Each point and vertical bar represents the mean ± SE, respectively, obtained from three independent experiments. Each experiment contained three cultures per replicate.

Immunoblot analysis of iNOS in extracts obtained from cultured rat mammary glands. The homogenates (10 mg/lane) were loaded onto the SDS-PAGE system and the immunoreactive substances were visualized by Western blot analysis with an alkaline phosphatase-conjugated secondary antibody as described in Materials and Methods. Lane 1, control; Lane 2, control 1 LPS (0.5 μg/ml); Lane 3, control + LPS + hydrocortisone (3 μM).
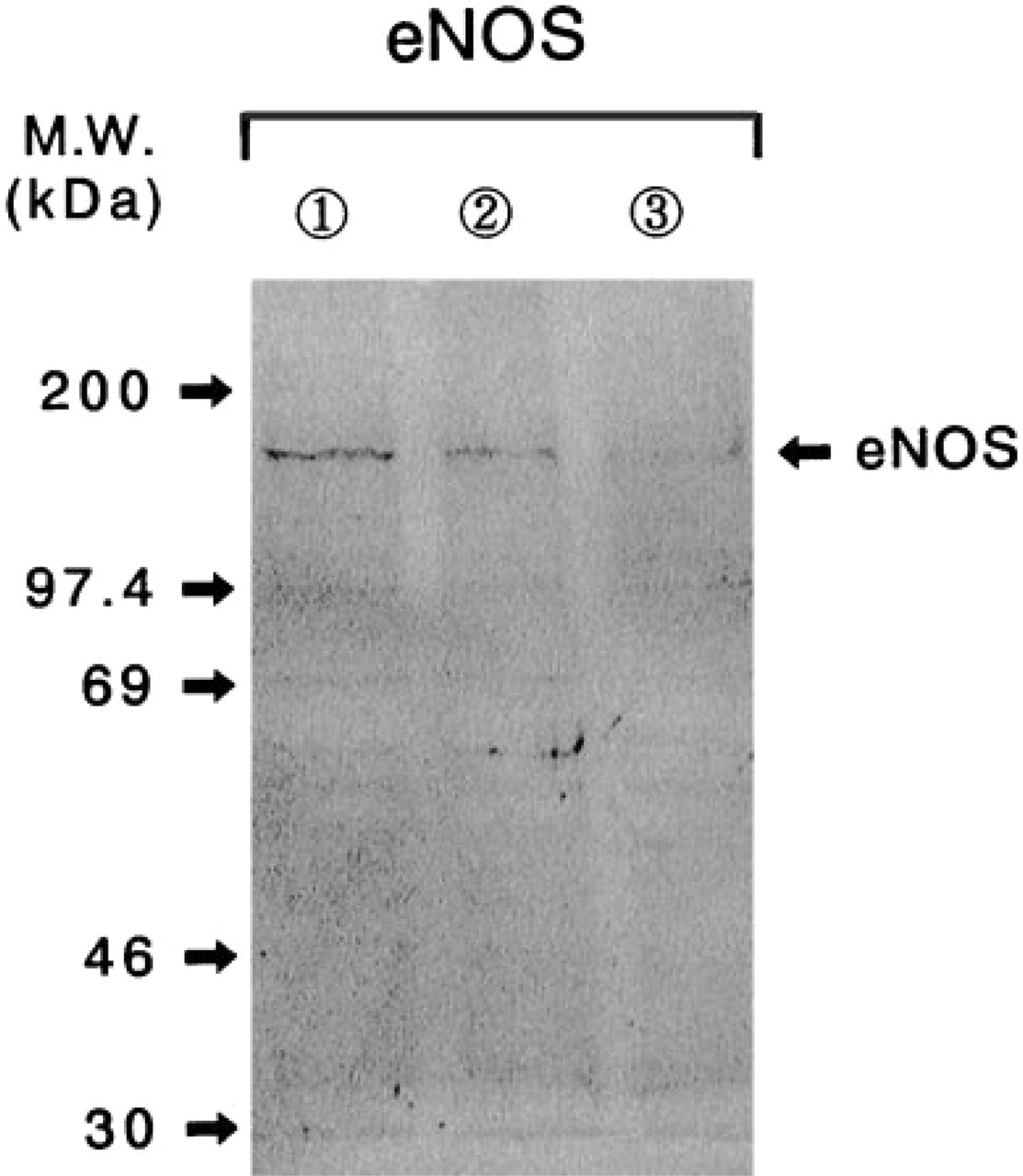
Immunoblot analysis of eNOS in extracts obtained from cultured rat mammary glands. The homogenates (18 μg/lane) were loaded onto the SDS-PAGE system and the immunoreactive substances were visualized as described in Materials and Methods. Lane 1, control; Lane 2, control 1 LPS (0.5 μg/ml); Lane 3, control + LPS + hydrocortisone (3 mM).
Immunohistochemical Observation of NOS Isoforms in Mammary Gland
Because the presence of NOS isoforms in the rat mammary gland was demonstrated by the immunoblot analyses, we carried out immunohistochemical staining of NOS isoforms in the rat mammary glands obtained after the organ culture with or without LPS stimulation to identify the particular location of NOS isoforms with specific antisera.
The immunoreactive signal of iNOS was clearly localized to the basal layers of alveoli and lactiferous ducts of the mammary glands obtained from the organ culture with LPS treatment (Figures 5b and 5b 9). These component cells are probably myoepithelial cells, because the staining pattern of mammary epithelial cells with anti-transferrin and anti-casein antisera was evidently different from the iNOS staining pattern in the alveoli and lactiferous ducts (Figure 6). In addition to the mammary parenchymal cells, the endothelial cell layers of mammary blood vessels were also intensively stained with iNOS antiserum (Figure 5b). Similar immunoreactivity with anti-iNOS antiserum was present in the mammary glands obtained from organ cultures without LPS treatment, although less intense staining with anti-iNOS antiserum was noted (data not shown). The histochemical staining of NADPH-diaphorase activity, which is identical to NOS (Dawson et al. 1991; Hope et al. 1991), showed similarity to the iNOS immunoreactive staining pattern (Figure 7). The immunostaining pattern of eNOS in the non-LPS-treated mammary glands was identical to that observed in the immunostaining of iNOS (Figures 5c and 5c 9). In contrast, the immunohistochemistry with anti-bNOS antiserum showed positive immunoreactivity in the epithelial cell layers of alveoli and lactiferous ducts of the mammary glands, although the staining intensity was relatively weak (Figures 5d and 5d 9). A positive immunoreaction with anti-bNOS antiserum was not detected in the blood vessels of the mammary glands (Figure 5d).
Discussion
Nitric oxide is increasingly appreciated as a major regulator in the nervous, immune, and cardiovascular systems. Apart from being an autocrine and paracrine mediator of homeostasis, NO has been found to inflict damage on important biomolecules, and it was suggested to contribute to the cytocidal action of macrophages and to many pathological events. Recent reports also implicate the overproduction of NO in inflammation, arthritis, myositis, and other diseases, and a role of NO in the carcinogenic process and tumor progression has been described (Esumi et al. 1995; Doi et al. 1996; Tamir and Tannenbaum 1996). In fact, NOS isoforms are expressed in breast cancer cell lines (Zeillinger et al. 1996) and are correlated with breast cancer metastasis (Thomsen et al. 1995; Dueñas-Gonzalez et al. 1997).
The tentative presence of NOS activity in the mammary gland was first reported by Thomsen et al. (1995) in human breast cancer and normal tissue and by Lacasse et al. (1996) in goat and cow mammary glands. The former report described that NOS activity was detectable only in invasive tumors compared with benign or normal tissue, that iNOS immunoreactivity was predominantly within tumor-associated macrophages, and that bNOS was observed in vascular endothelial and myoepithelial cells in breast tumors. In the latter study, eNOS activity was detected only in the vascular endothelium and secretory epithelium of the two species. In our present study, however, it was demonstrated by immunoblot analysis and immunohistochemical criteria that three isoforms of NOS are present in the rat mammary gland. To our knowledge, this is the first observation of three NOS isoforms in the normal rat mammary glands. We found that iNOS and eNOS isoforms localize to myoepithelial cells of the glandular epithelium and the endothelial cells of blood vessels, whereas the bNOS isoform is found in the epithelial cells of mammary parenchymal glands. Although we do not have a clear explanation regarding the discrepancy in the localization of eNOS and bNOS isoforms between the reports mentioned above and our present findings, species and cell type differences may account for the inconsistencies.
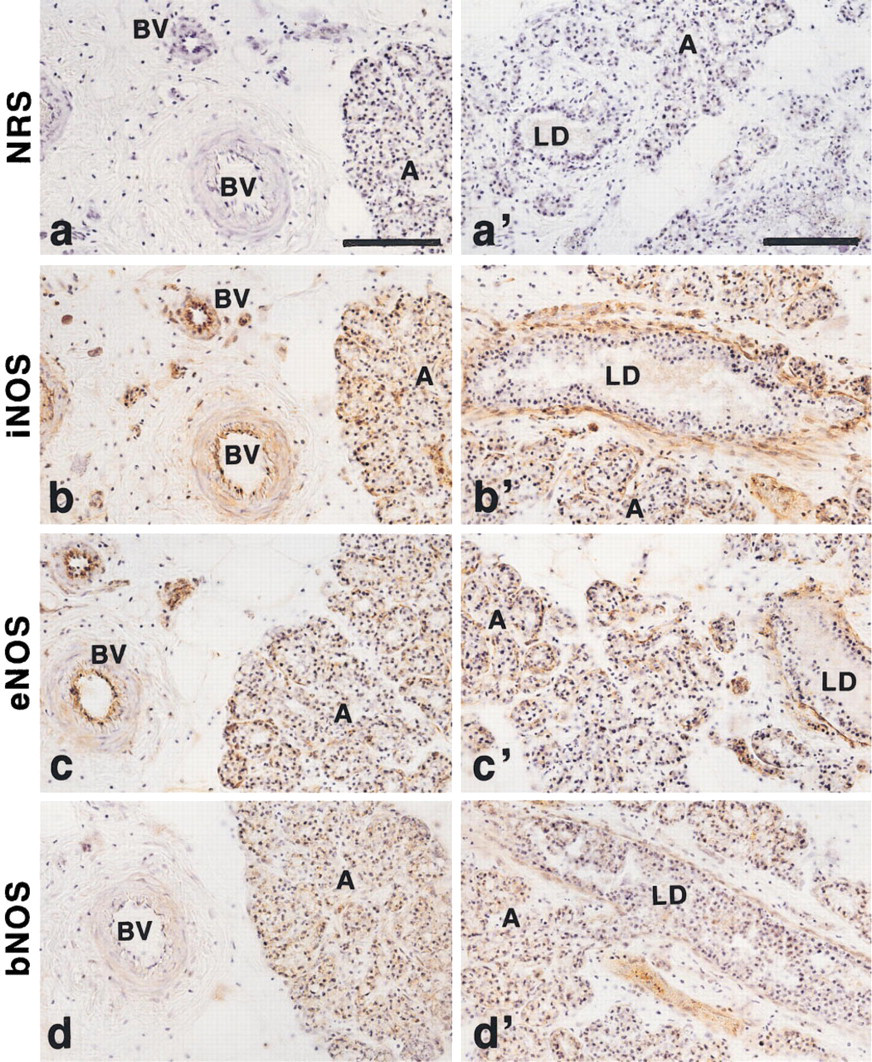
Immunohistochemistry of NOS isoforms in rat mammary glands. The mammary glands were isolated from hormone-primed Wistar-MS rats, cultured, and subjected to immunohistochemistry with specific antisera against NOS isoforms as described in Materials and Methods. For iNOS immunostaining (
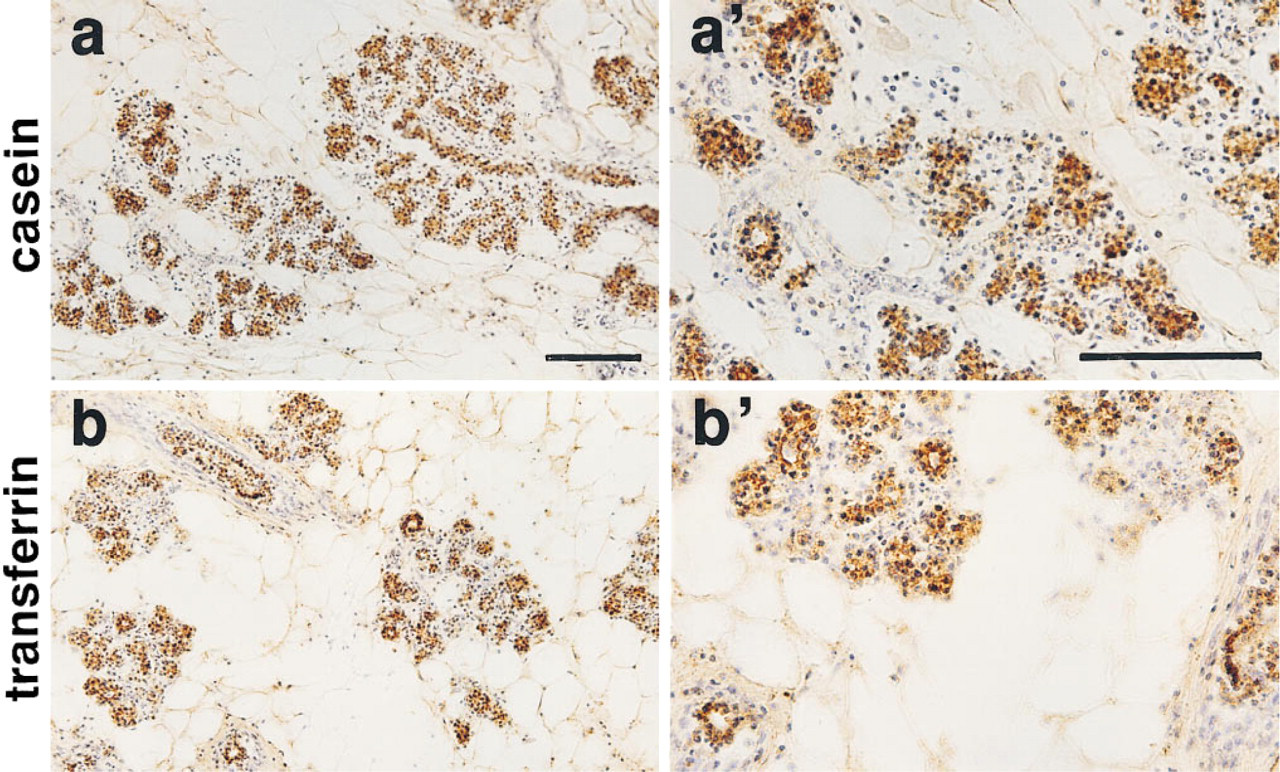
Immunohistochemistry of casein and transferrin in rat mammary glands. The mammary glands were obtained from control cultures and processed for casein (
Interestingly, iNOS was present not only in LPStreated mammary glands but also in the mammary glands that were not stimulated with LPS, although the amount of iNOS in the normal unstimulated mammary glands was relatively minute in comparison with the LPS-stimulated mammary glands. This was also reflected as spontaneous NO production in the NO concentration of culture medium in the mammary gland organ cultures without LPS stimulation. This is suggestive of a basal expression of iNOS and of other constitutive NOS isoforms in the unstimulated rat mammary glands. Because there are precedents for the expression of iNOS isoform in normal rat kidney (Morrissey et al. 1994) and guinea pig skeletal muscle (Gath et al. 1996,1997), it is not unlikely that the iNOS isoform is constitutively expressed to some extent in the normal rat mammary gland. However, it should be noted that the inevitable stress given to the isolated tissues during the dissection process and the subsequent culture conditions may have contributed to the induction of the iNOS isoform in the normal mammary glands.
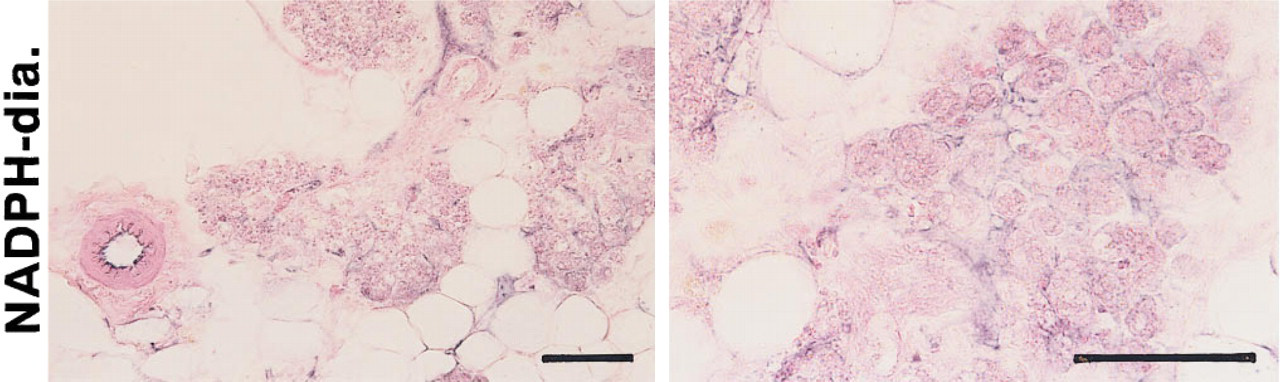
Histochemistry of NADPH-diaphorase activity in rat mammary glands. The mammary glands shown here were obtained from LPDtreated cultures and processed for NADPH-diaphorase activity staining as described in Materials and Methods. Bars = 100 μm.
Another surprising finding of our study is that the expression of the eNOS isoform in the mammary gland declined after the exposure of the mammary glands to LPS in the culture, although the iNOSderived NO production masked a reduced NO production by eNOS in the cultures with LPS treatment. The downregulation of constitutive NOS expression by iNOS was recently reported not only for eNOS in cultured bovine coronary venular endothelial cells (Lu et al. 1996) and rat glomerular cells (Schwartz et al. 1997) but also for bNOS in the skeletal muscle of guinea pig (Gath et al. 1997), although these isoforms are usually considered to be constitutively expressed genes. It was suggested by Schwartz et al. (1997) that NO autoinhibition via complex interactions among NOS isoforms after LPS treatment may contribute to renal function. In addition, Lu et al. (1996) showed that LPS decreased the eNOS expression in cultured bovine coronary venular endothelial cells at protein and mRNA levels in a dose- and time-dependent manner, and concluded that the reduction of eNOS by LPS treatment resulted from an increased degradation rate of its transcripts. Therefore, it is likely that similar downregulation mechanisms occur in a reciprocal modulation of the eNOS and iNOS expressions of the mammary gland after the LPS exposure.
It is known that the growth and differentiation of the mammary gland are essentially controlled by multiple interactions of several peptide and steroid hormones from endocrine organs, such as the pituitary, ovary, and adrenal gland. Subsequently, development occurs in several phases, characterized by distinct morphological features (Topper and Freeman 1980). However, it was reported recently that release of these hormones from the endocrine organs is influenced by an NO-generating system. Higher levels of NO produced by iNOS in anterior pituitary cells may attenuate prolactin (PRL) and growth hormone (GH) release during emergency conditions of immune and inflammatory reactions, whereas low amounts of NO produced constitutively may take charge of subtle physiological adaptation (Vankelecom et al. 1997). Steroidogenesis in cultured ovarian cells is also suppressed by an inhibitory action of NO on P-450 aromatase activity (Masuda et al. 1997). Therefore, it is likely that development of the mammary gland is indirectly regulated through the NO-generating system by which hormone release is controlled in the endocrine organs. Conversely, steroids have been shown to regulate the NO synthesis in diverse types of cells and organs. Various glucocorticoids, mineralocorticoids, and sex steroids inhibited the spontaneous formation of nitrite in the medium of cultured rat hepatocytes (Pittner and Spitzer 1993). Yallampalli et al. (1994) showed that steroid hormones modulate the production of NO in the rat uterus, contributing to uterine contractility during pregnancy and the initiation of term or preterm labor. Taken together, these observations indicate that it is plausible that an NO-generating system may closely participate in the regulation of morphological and functional features of the mammary gland, because the growth and differentiation of the mammary gland are dependent on the hormonal conditions, which change with the developmental stages of this complicated gland.
Involution of the mammary gland is a tissue remodeling process, of which an integral component is the apoptosis of mammary epithelial cells (Furth et al. 1997). After lactation the mammary gland enters into an involutional process, which culminates in the return of the tissue to a quiescent stage within a short period. The beginning of this irreversible involution stage is characterized by decreased systemic hormone levels and proteinase action. Perplexingly, there a considerable number of studies have shown contrary effects of NO (i.e., apoptosis induction and cell protection by NO). The expression of cell-cycle control proteins, such as c-Myc and P53, increased after incubation with NO donors in vascular smooth muscle cells, whereas Bcl-2 protein, a survival protein, decreased (Nishio and Watanabe 1998). Wang and co-workers (1997) detected iNOS expression in the glomerular and tubulointerstitial cells of Class IV lupus nephritis in which p53 overexpression was correlated. These studies suggested that NO production plays a role in the occurrence of apoptosis through proto-oncoprotein expression. However, the mechanism of the inhibition of hepatocyte apoptosis by NO has been shown by Kim and colleagues (1997); who postulated that NO prevents apoptosis in hepatocytes by inhibiting caspase-3-like protease activity via a cGMP-dependent mechanism and by inhibition of caspase-3-like activity through protein S-nitrosylation. Although the roles of NO in apoptosis are still controversial, perhaps because NO can be either beneficial or detrimental depending on the cell type, it might help our understanding of the precise roles of the NO-generating system in mammary gland physiology to clarify the stage-specificity of the expression of NOS isoforms, which may be influenced by pituitary, ovarian, and adrenal hormones.
In any event, our present data demonstrate that the presence of three NOS isoforms in the rat mammary gland is certain and that these NOS isoforms may correlate with mammary gland development and regulatory functions. Additional studies are needed to elucidate the precise roles of NO in the physiology and pathophysiology of the mammary gland.
Footnotes
Acknowledgments
Supported by grants for Special Project Research for Experimental Studies on Radiation Health, Detriment and Its Modifying Factors, and for Research Programme on Bioregulatory Mechanisms of the National Institute of Radiological Sciences.
We are grateful to Dr Yuji Ishikawa, Senior Researcher in the Division of Biology and Oncology, for helpful advice on the preparation of the frozen sections from cultured mammary glands.