
Abstract
CIAPIN1, a newly identified antiapoptotic molecule that plays an essential role in mouse definitive hematopoiesis, is considered a downstream effector of the receptor tyrosine kinase-Ras signaling pathway. Our previous studies have indicated that CIAPIN1 is involved in the development of multidrug resistance (MDR) in gastric cancer cells. However, the mechanism of CIAPIN1-mediated antiapoptosis and MDR has not been fully elucidated. To reveal the possible physiological role of CIAPIN1, we examined the expression and distribution of CIAPIN1 in fetal and adult human tissues using immunohistochemistry. We found that CIAPIN1 was ubiquitously distributed in fetal and adult tissues, and was localized in both the cytoplasm and the nucleus. The expression patterns of CIAPIN1 were similar in fetal and adult tissues, and was correlated with the previously described expression pattern of p21ras. These observations suggest that CIAPIN1 expression appears to be involved in cell differentiation, and that it might exert universal and possibly important physiological functions under the regulation of Ras in humans.
C
Identification of the tissue or cell types that express CIAPIN1 in vivo will provide a clue to the elucidation of its physiological function. CIAPIN1 mRNA expression in a number of human adult tissues was characterized by Northern blotting in a previous study (Shibayama et al. 2004). However, mRNA levels only indirectly represent the contents of the translated protein product. Immunohistochemistry and Western blotting provide more accurate measurements than does Northern blotting on the temporal expression and quantitative levels of the protein. Furthermore, the approach using a whole-tissue extract may provide misleading information when cells within a given tissue express CIAPIN1 distinctly at different stages of differentiation or when there exist different subsets of specialized cells that express CIAPIN1 distinctly. The aim of this study was to detect expression of CIAPIN1 in normal fetal and adult human tissues using immu-nohistochemistry, to establish CIAPIN1 expression profiles in humans.
Materials and Methods
Tissue Specimens
Commercially available fetal and adult human tissue arrays were obtained from Cybrdi (Xi'an, China). The fetal tissue microarray (ec01-001) contained 96 cores of normal, four-to five-month gestational age fetal tissues representing most major organ systems. Each section was a 1-mm-diameter disk of formalin-fixed, paraffin-embedded tissue. The adult tissue array (nc01-001) contained 53 points of a 1.5-mm-diameter disk of formalin-fixed, paraffin-embedded tissues representing histologically normal organs from individuals aged 25-69 years.
Mouse tissues were obtained from an 8-week-old healthy BALB/c mouse (Animal Center of the Fourth Military Medical University). The stomach, intestines, liver, pancreas, spleen, and kidney were taken after perfusion in 4% paraformaldehyde (dissolved in 0.1 M phosphate buffer, pH 7.4). All tissues were postfixed with 4% paraformaldehyde at room temperature for at least 3 hr. The fixed tissues were then dehydrated in ethanol and embedded in paraffin. Sections (5-μm thick) were cut and mounted for immunostaining. In addition, fresh tissue samples of the mouse stomach, intestine, liver, pancreas, spleen, and kidney taken prior to perfusion in paraformaldehyde were homogenized in radio-immunoprecipitation assay (RIPA) lysis buffer (50 mM Tris-Cl, pH 8.0, 150 mM NaCl, 0.1% SDS, 0.02% sodium azide, 100 μg/ml PMSF, 1 μg/ml aprotinin, 1% nonidet P-40, and 0.5% sodium deoxycholate) in an ice bath followed by centrifugation at 12,000 rpm for 10 min at 4C. The supernatants were collected for Western blotting analysis.
Cell Lines
Cells of human gastric adenocarcinoma cell line SGC7901 and of mouse fibroblast cell line NIH3T3 were cultured in DMEM supplemented with 10% FCS (Sigma Chemical Co.; St. Louis, Mo). The cells were maintained in a humidified chamber with 5% CO2 at 37C. At approximately 80% confluence, the cell layer was washed with ice-cold PBS (pH 7.4) and scraped off. Cells were harvested by centrifugation and lysed in RIPA lysis buffer in an ice bath for 30 min. After centrifugation at 12,000 rpm for 10 min at 4C, the supernatant was collected for Western blotting analysis.
Antibodies and Reagents
Mouse monoclonal antibody (MAb) against CIAPIN1 (clone 3C6) was raised by immunizing with full-length recombinant hCIAPIN1 expressed in Escherichia coli and standard cell fusion techniques, as described elsewhere (Hao et al. 2005). This MAb recognizes hCIAPIN1 and cross-reacts with mCIAPIN1. Mouse Streptavidin-Peroxidase immunostaining kit (SP-9000 Histostain-Plus Kit) was from ZYMED (Zymed Laboratories; San Francisco, CA).
Immunohistochemistry
Immunohistochemistry was performed using the Histostain-Plus SP kit, which offers superior sensitivity. Briefly, the sections were deparaffinized with xylene and rehydrated through gradient ethanol immersion. Endogenous peroxidase activity was quenched by 0.3% (v/v) hydrogen peroxide in methanol for 20 min, followed by three 5-min washes with PBS. The sections were then blocked with 10% (v/v) normal goat serum in PBS for 1 hr, followed by overnight incubation at 4C with the anti-CIAPIN1 antibody diluted (1:200, initial concentration 1.8 mg/ml) in PBS containing 3% (wt/vol) BSA. Negative control was performed by replacing the primary antibody with preimmune mouse serum. After three 5-min washes with PBS containing 0.02% (v/v) Tween-20 (PBST), the sections were treated with biotinized goat anti-mouse antibody for 20 min at room temperature, followed by three additional 5-min washes with PBST. Then the specimens were incubated with streptavidin-horseradish peroxidase for 20 min at room temperature, followed by repeated washes, as described above. Reaction product was visualized with DAB at room temperature for 5 min. Sections were counterstained with hematoxylin for 30 sec and rinsed with tap water, immediately dehydrated by sequential immersion in gradient ethanol and xylene, then mounted with Permount on coverslips. Images were obtained under a light microscope (Olympus BX51; olympus, Japan) equipped with a DP70 digital camera.
The IHC staining for hCIAPIN1 protein was evaluated by scanning the entire tissue specimen under low-power magnification (X40) and then confirmed under high-power magnification (X200 and Xl000). The staining intensity was stratified as - (absent), + (weak), ++ (moderate), or +++ (strong).
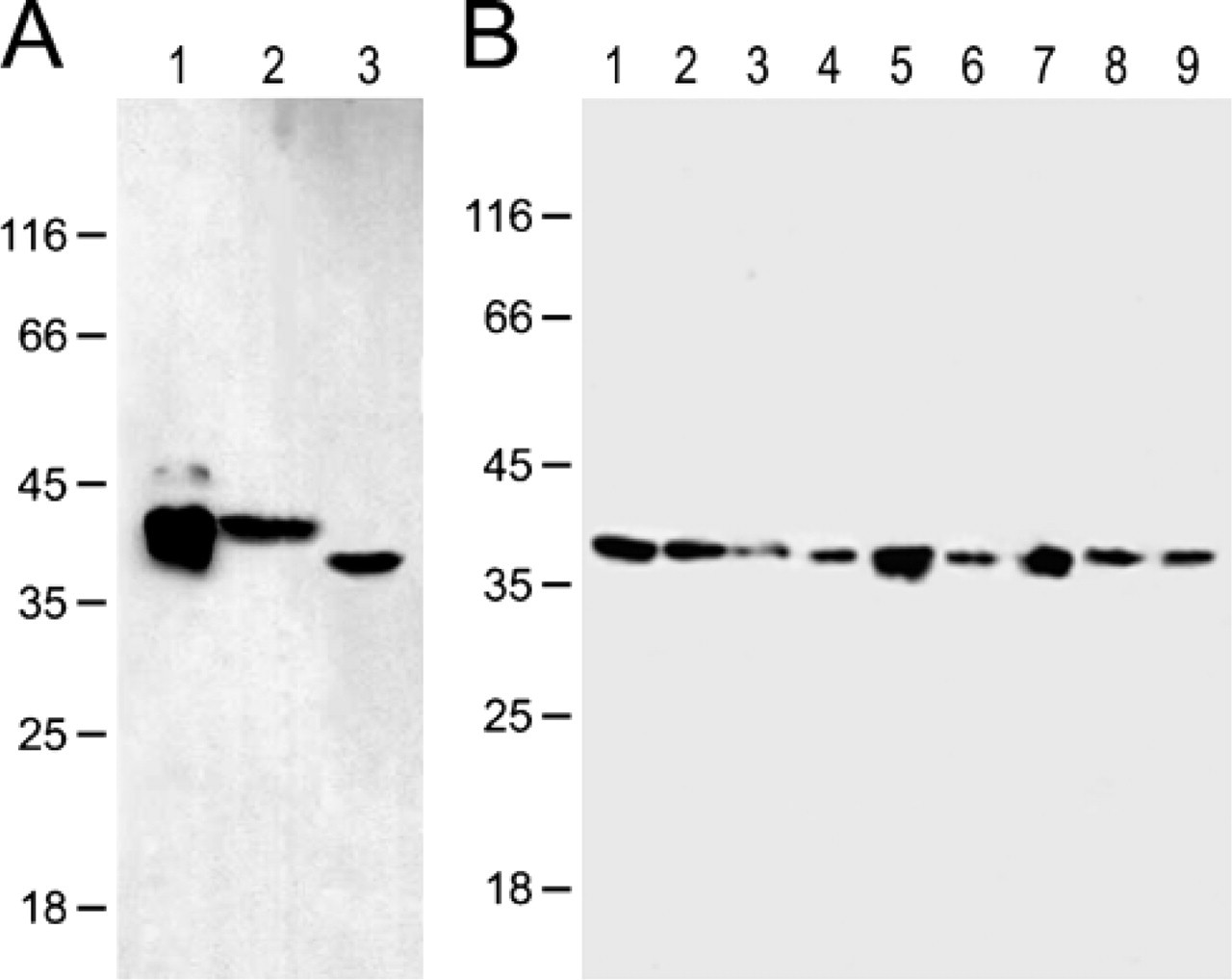
Identification of the specificity of the anti-CIAPIN1 monoclonal antibody by Western blotting. (
Western Blotting
Equal amounts of total proteins were loaded on 12% SDS-PAGE and electroblotted to a nitrocellulose membrane. Nonspecific binding was blocked with 5% non-fat milk in PBS for 1 hr at room temperature. Then the membrane was incubated with anti-CIAPIN1 MAb (1:400) overnight at 4C, rinsed with TBST three times, then incubated with HRP-labeled goat anti-mouse IgG for 1 hr. After three washes with TBST, the bands were developed with the enhanced chemiluminescence reagent for 5 min.
Results
Specificity of the In-House-Generated Anti-CIAPIN1 MAb
To confirm the specificity of the in-house-generated anti-CIAPIN1 MAb, we performed Western blotting to test whether the antibody reacts with any other antigens in human and mouse tissues and cells. As shown in Figure 1, a single band was detected on each lane of the tissue and cell protein samples, confirming the specificity of the anti-CIAPIN1 MAb.
To help define the tissue and cell type expression and distribution, we examined the expression of CIAPIN1 in the major organs of the mouse, including the stomach, small intestine, pancreas, liver, kidney, and spleen. The immunostaining of mouse tissues with anti-CIAPIN1 MAb demonstrated distinct specific immunoreactive cells (Figures 2A-2G). Replacement of the primary antibody with preimmune mouse serum abolished the immunostaining in the tissues, confirming the specificity of the immunoreaction (Figures 2B, 2B', 2C, and 2C').
We also compared the immunostaining of mouse tissues with and without antigen retrieval. Antigen retrieval was performed by microwaving (500 W) the sections for 15 min in citric acid buffer (2 mM citric acid and 9 mM trisodium citrate dehydrate, pH 6.0). No obvious improvement was found in immunostaining after antigen retrieval (data not shown). Thus, antigen retrieval was omitted in the process of immunohisto-chemical staining.
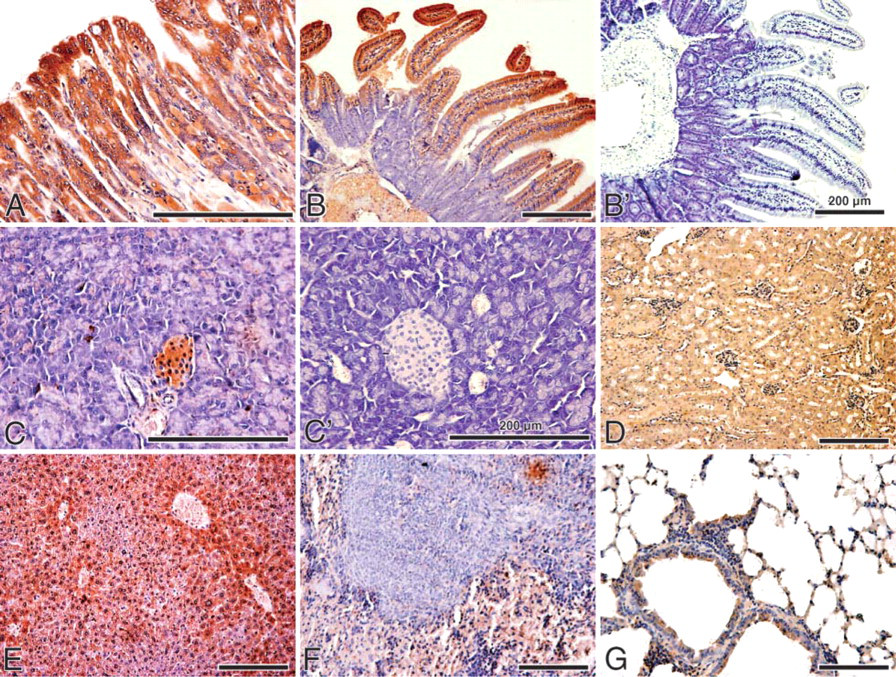
Expression of CIAPIN1 in normal mouse tissues. (
Distribution of CIAPIN1 in Human Cells and Tissues
CIAPIN1 immunoreactivity with the specific MAb was generally localized in both the cytoplasm and the nucleus. Under high magnification, positive staining of intra-nuclear structure characteristic of nucleoli was clearly observed in both adult and fetal tissues (Figures 3A and 3F and Figures 4A, 4E, and 4G). This was especially evident in the cells where CIAPIN1 immunostaining was strong.
CIAPIN1 protein was found in virtually all of the normal fetal and adult tissues examined, although the expression levels differed among tissues. The distribution of CIAPIN1 in human tissues was consistent with that in mouse tissues, and the distribution pattern of CIAPIN1 in fetal tissues was similar to that in adult tissues. The distribution and the quantitative expression of CIAPIN1 in fetal and adult human tissues is described in detail below and further summarized in Table 1.
Muscles and Bones. Both fetal and adult skeletal muscle and myocardial cells showed strong CIAPIN1 staining (Figures 3A and 3B and Figure 4A). Distinct cytoplasmic and nuclear staining, as well as accumulation in the nucleolus, was observed in myocardial cells (Figure 3A and Figure 4A). Smooth muscle cells of the gastrointestinal tract showed moderate immunostaining, whereas smooth muscle cells in the uterus and oviduct showed only weak immunostainig. In fetal bones, osteobasts and osteoclasts were strongly stained (Figure 3C).
Skin and Mammary Glands. Strong immunostaining was observed in the epidermis (stratified squamous epithelium), sweat glands, and folliculus. The dermis was weakly stained (Figure 3D). In endothelial cells of dermal blood vessels, moderate immunostaining was observed. In adults, CIAPIN1 immunostaining was stronger in the stratum corneum and intermediate layers than in the basal layers (data not shown).
In quiescent mammary glands, CIAPIN1 immunostaining was moderate in duct epithelia and weak in alveolar epithelia, but it was absent in cells in the underlying tissues, e.g., stromal fibroblasts and adipocytes.
Digestive System. Similar to what was observed in the epidermis, squamous, non-keratinizing epithelia of the esophagus were strongly stained. Both intermediate and luminal layers of the squamous esophageal strata showed stronger reactivity than did the basal layers (Figure 4B).
The developing gastric epithelial cells showed moderate staining (data not shown). In adults, expression patterns of CIAPIN1 in the fundus differed from those in the antrum. Immunoreactivity was strong in the parietal cells of the fundus and body gastric glands, and moderate to strong in the middle portion (neck) and deep gastric glands in the antrum. The surface epithelial cells were weakly stained (Figure 4C).
In the developing small intestine and colon, CIAPIN1 immunostaining diffused throughout the mucosal layer, and a gradient of CIAPIN1 immunoreactivity was observed along the crypt-villous axis. Relatively moderate reactivity was seen in crypt cells, and progressively stronger reactivity appeared toward the villous tips (Figures 3E and 3F). Strong, diffused CIAPIN1 expression was also detected in the fetal rectum (data not shown). In adult colon, although moderate immunopositive signal was localized in the apical mucosa, the glands were negatively or weakly immunostained (Figure 4D). (Adult small intestine and rectum were unavailable).
Hepatocytes were strongly stained in both fetal and adult livers. In fetal livers, immunostaining of hemotapoietic cells was comparable to that of hepatocytes (Figure 3D and Figure 4E). In fetal pancreas, moderate CIAPIN1 staining was observed in ductal epithelia and acinar secretory cells (Figure 3H). The most prominent CIAPIN1 staining in adult pancreas was found in cells of the islets of Langerhans, which showed strong specific immunostaining, whereas ductal epithelia and acinar secretory cells were only weakly stained (Figure 4F).
Brain. Moderate immunopositive reactions for hCIA-PIN1 were found in the cytoplasm and the nucleus of the nerve cells and the glial cells of both fetal and adult brains (Figure 4G and data not shown).
Lungs. Luminal surfaces of bronchiolar and bronchial epithelial cells, including ciliated and non-ciliated epithelial cells, showed specific moderate staining. Moderate immunoreactivity was also observed in other alveolar lining cells and alveolar macrophages (Figure 3I and Figure 4H).
Urinary System. Positive immunoreactions were observed in the epithelium of the proximal and distal convoluted tubules and collecting tubules. The proximal tubule cells showed the strongest staining, and the collecting tubules showed the weakest staining. The renal glomerulus and blood vessel cells were weakly stained. The surrounding fibrous stroma was negative. Fetal and adult kidney showed similar immunostaining patterns (Figure 3J and Figure 4I). In adult prostate, only the glandular epithelia showed weak specific immunostaining (Figure 4J).
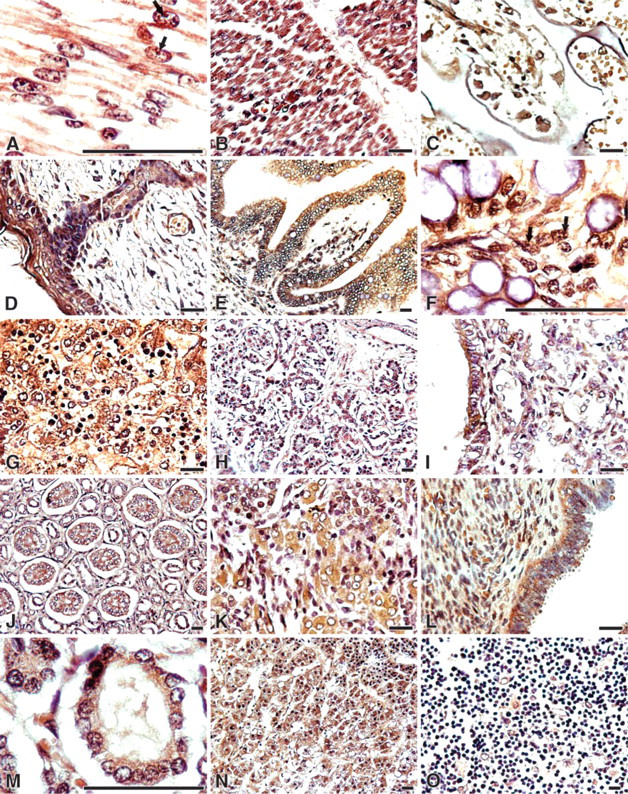
Expression of CIAPIN1 in normal fetal human tissues. (
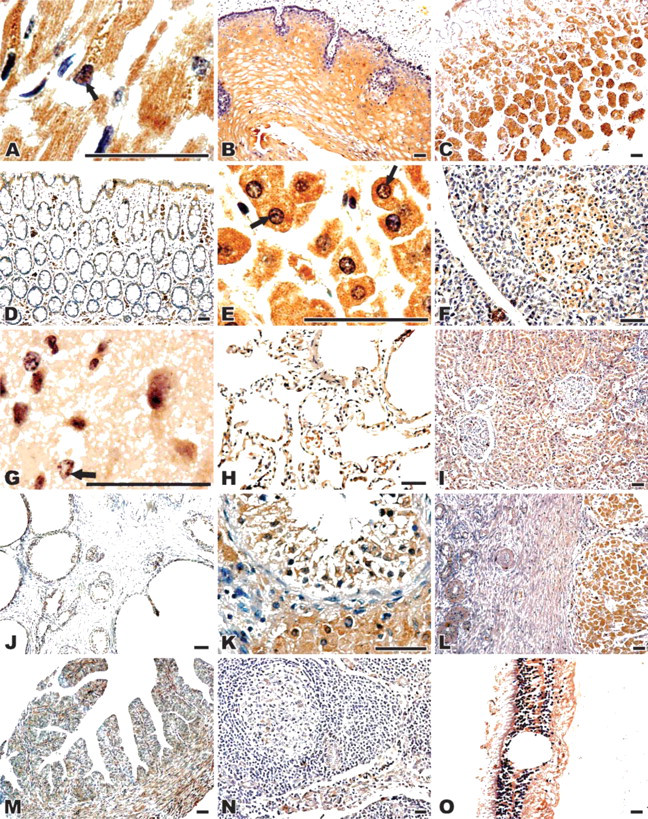
Expression of CIAPIN1 in normal adult human tissues. (
Male Germinal System. All the parenchymal cell types in the testis were positively stained, including Sertoli and Leydig cells, fibromyocytes, and spermatogonia, with the strongest staining in spermatogonia and Leydig cells (Figure 3K and Figure 4K).
Female Germinal System. Both fetal and adult endometrium showed moderate staining (Figure 3L and data not shown). In the ovary, the primary and secondary oocytes and follicular cells showed moderate immunostaining (Figure 4L and data not shown). In adult ovary, luteal cells showed strong staining, but the stromal components were weakly stained (Figure 4L). Epithelial cells in the oviduct mucosa were weakly stained (Figure 4M). In the placenta, Hofbauer cells showed strong immunoreaction, but other cells were weakly to moderately stained (data not shown).
Endocrine Organs. CIAPIN1 expression in fetal thyroid and adrenal gland was detected in this study. Thyroid follicular epithelial cells showed moderate immunostaining (Figure 3M). The adrenal glands displayed a distinct pattern of CIAPIN immunostaining: all secretory epithelial cells of the zona fasciculata and zona reticularis in the adrenal cortex displayed strong staining, whereas cells of the zona granulosa showed moderate staining (Figure 3N).
Lymphatic Tissues. Mature lymphatic cells showed negative immunostaining. But in the lymph nodes, cells in the germinal center, lymphoblasts, trabecula, and postcapillary venule endothelium were weakly stained (Figure 4N). In fetal thymus, epithelial reticular cells demonstrated moderate immunostaining whereas thymocytes demonstrated negative to weak immunostaining (Figure 3O). In fetal spleen, hematopoietic cells expressed a high level of CIAPIN1 (data not shown).
Eyeballs. Retinal cells and ciliary cells were strongly stained (Figure 4O), whereas sclera epithelia were moderately stained (data not shown).
Discussion
This is the firststudy to establishthe expression pattern of hCIAPIN1 in normal fetal and adult human tissues. Commercially available multi-tissue slides enabled us to conduct a large survey of tissues simultaneously, and thus the comparison should be more precise and authentic, because the tissue disks were stained under the same conditions (Oberst et al. 2003; Watanabe et al. 2005).
In this study, we found that CIAPIN1 was ubiquitously expressed in human tissues, although the expression levels differed among tissues. This indicates that CIAPIN1 cannot be considered a specific tissue marker. Our results on CIAPIN1 protein expression are consistent with the reported data on CIAPIN1 mRNA quantification in major human organ extracts obtained by Shibayama et al. (2004), who demonstrated that human CIAPIN1 was expressed in a wide range of organs, with highest expression levels in heart, liver, and pancreas. Our results are also supported by SAGE anatomic analysis (http://cgap.nci.nih.gov/microarray/GeneList) and the fact that cDNA of human CIAPIN1 is cloned from a wide variety of tissues (Loftus et al. 1999; Wiemann et al. 2001; Strausberg et al. 2002) (GenBank accession numbers are available at http://www.ncbi.nlm.nih.gov/Unigene/clust.cgi? ORG = Hs and CID = 4900)
On the basis of the widespread distribution of CIAPIN1, we postulate that CIAPIN1 functions universally in all human tissues. The ubiquitous distribution of CIAPIN1 as an effector of the Ras signal transduction pathway is reasonable, because proto-oncogene Ras is widely expressed in developing and adult human tissues (Chesa et al. 1987; Furth et al. 1987; Radosevich et al. 1989) and the Ras signaling pathway plays a crucial role in cell survival, growth, and differentiation (Peyssonnaux and Eychene 2001). Interestingly, the distribution pattern of CIAPIN1 expression in adult human tissues observed in this study is closely correlated with the previously reported distribution pattern of p21ras expression in normal human tissues (Chesa et al. 1987). The correlation in distribution between p21ras and CIAPIN1 expression suggests that the expression of CIAPIN1 is strictly regulated by Ras, and that CIAPIN1 is an important effector of the Ras signaling pathway. In normal tissues, the activation of Ras is largely dependent on a diverse array of extracellular signals, such as growth factors, cytokines, and hormones (Campbell et al. 1998). Virtually all human tissues and cells are under consistent or intermittent stimulation by growth factors, cytokines, or hormones. For example, hepatocyte growth factor (HGF) acts as a crucial regulator of cell growth and survival for hepatocytes (Uehara et al. 1995) such as IL-6-type cytokines (i.e., cardiotrophin-1, leukemia inhibitory factor, and oncostatin M) for cardiac myocytes (Yamauchi and Kishimoto 2000), and insulin-like growth factor 1, leukemia inhibitory factor, and HGF for skeletal-muscle cells (Husmann et al. 1996). Growth factors and cytokines are of vital importance in embryonic and fetal development (Takeda and Iwashita 1993; Poulsen et al. 1996). Often a specific growth factor or cytokine influences the development of a series of organs and vice versa. For example, in addition to its involvement in hematopoiesis, EPO is essential for the development of the neural system, myocardium, and skeletal muscles (Lacombe and Mayeux 1998; Yu et al. 2002); as is epidermal growth factor for the development of kidney (Jung et al. 2005), thymocytes (Freitas et al. 1998), stomach (Tremblay et al. 1997), and myocardium (Goldman et al. 1996); HGF for liver (Kolatsi-Joannou et al. 1997), stomach (Kong et al. 1998), lung (Sato and Takahashi 1997), and pancreatic islets (Otonkoski et al. 1994); and colony-stimulating factor and insulin-like growth factor-1 for the development of the fetus (Lammie et al. 1994; Roth and Stanley 1995; Benito et al. 1996; Calhoun et al. 1999). All of these cytokines and growth factors activate Ras by binding their receptors, and their functions are dependent, at least in part, on the Ras signaling pathway (Margolis and Skolnik 1994). The widespread distribution of CIAPIN1 as a mediator of the Ras signaling pathway might reflect the universal function of Ras in human tissues.
CIAPIN1 expression in histologically normal adult and fetal human tissues

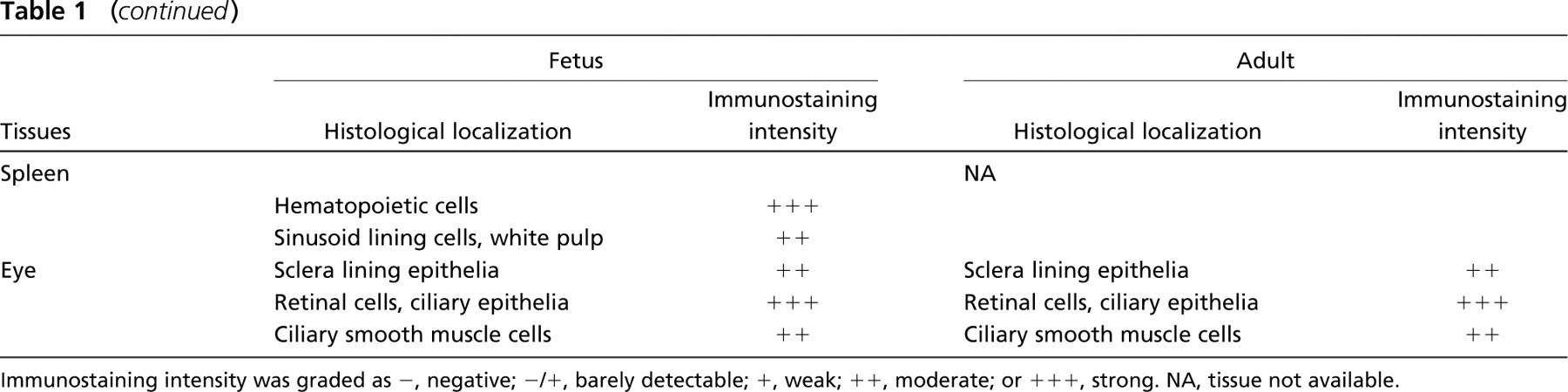
Immunostaining intensity was graded as -, negative; -/+, barely detectable; +, weak; ++, moderate; or +++, strong. NA, tissue not available.
The results on the functions of proto-oncogene Ras in normal tissues are conflicting. Recently, more and more evidence has suggested that wild-type Ras functions as a tumor suppressor instead of an oncogene. Wild-type Ras suppresses cell proliferation and promotes cell differentiation in normal or tumorous tissues (Furth et al. 1987; Rebollo and Martinez 1999; Diaz et al. 2002; James et al. 2003; Passioura et al. 2005; Singh et al. 2005). In the present study, it was observed that levels of CIAPIN1 expression were higher in the differentiated mature cells than in the proliferative basal layer cells in the skin and esophagus, at the tips of the villi than in the crypts in the small intestine, and in the apical epithelia than in the glands in colon. Moreover, CIAPIN1 expression was upregulated in the terminal differentiated neural cells and muscle cells, but was downregulated in gastric and liver cancers, compared with their noncancerous counterparts (Hao Z, et al., unpublished data). These observations suggest that CIAPIN1 expression is associated with cell proliferation, and that the differentiation-promoting effect of Ras might be mediated by CIAPIN1. CIAPIN1 serves as an antiapoptotic molecule in Ba/F3 cells (Shibayama et al. 2004), but its functions in other tissues or cell types remain undetermined. Our observations demonstrate that actively metabolic tissues and cells such as hepatocytes, parietal cells of gastric glands, proximal tubules, endocrine cells, and myocardium express relatively high levels of CIAPIN1. However, it is yet to be determined whether a high apoptotic stress causes these cells to express higher levels of CIAPIN1 to prevent apoptosis.
Unlike the previous report on exclusive cytoplasmic localization of CIAPIN1 (Shibayama et al. 2004), our immunohistochemical results demonstrated that CIA-PIN1 occurs in both cytoplasm and nucleus, and further accumulates in the nucleoli. This is one of the prominent features of CIAPIN1 distribution revealed in this study. In addition, we found that CIAPIN1 contains a non-classical nuclear localization signal (KKRK, amino acid residues 236-239), and bioinformatic analysis predicts that CIAPIN1 is preferentially localized in the nucleus (data not shown). This observation necessitates further studies to confirm the subcellular localization of CIAPIN1. Because the aim of this study is to establish the expression pattern of CIAPIN1 in different tissues, further observations and analyses on the subcellular localization of CIAPIN1 are not described here.
In conclusion, CIAPIN1 is ubiquitously distributed in fetal and adult tissues, and localized in both the cytoplasm and the nucleus. CIAPIN1 expression appears to be involved in cell differentiation, and might exert universal and possibly important physiological functions through the regulation of Ras in humans, although further investigation is required.
Footnotes
Acknowledgments
This work was supported by a grant from the Chinese National Foundation of Natural Sciences (no. 30471989).
We thank Dr. Mei Zhang, Department of Pathology, 2nd Hospital of Xi'an Jiaotong University, for histological assessment.