
Abstract
SPARC (secreted protein, acidic, and rich in cysteine) is a matricellular protein that is present in the intervertebral disc; in man, levels of SPARC decrease with aging and degeneration. In this study, we asked whether targeted deletion of SPARC in the mouse influenced disc morphology. SPARC-null and wild-type (WT) mice were studied at 0.3–21 months of age. Radiologic examination of spines from 2-month-old SPARC-null mice revealed wedging, endplate calcification, and sclerosis, features absent in age-matched WT spines. Discs from 3-month-old SPARC-null mice had a greater number of annulus cells than those of WT animals (1884.6 ± 397.9 [mean ± SD] vs 1500.2 ± 188.2, p=0.031). By 19 months discs from SPARC-null mice contained fewer cells than WT counterparts (1383.6 ± 363.3 vs 1466.8 ± 148.0, p=0.033). Histology of midsagittal spines showed herniations of lower lumbar discs of SPARC-null mice ages 14–19 months; in contrast, no herniations were seen in WT age-matched animals. Ultrastructural studies showed uniform collagen fibril diameters in the WT annulus, whereas in SPARC-null disc fibrils were of variable size with irregular margins. Consistent with the connective tissue deficits observed in other tissues of SPARC-null mice, our findings support a fundamental role for SPARC in the production, assembly, or maintenance of the disc extracellular matrix.
T
In other tissues, SPARC is known to play dynamic roles in modulating interactions between cells and their ECM; in collagen fibrillogenesis, deposition, and remodeling; and in growth factor efficacy (Bradshaw and Sage 2001). Because SPARC has been shown to bind to protein constituents of the ECM (collagen types I–V and VIII, vitronectin, and thrombospondin-1) (Brekken and Sage 2001), it has been proposed that SPARC modulates cell-ECM interactions and influences cell behavior during remodeling of the ECM (Sweetwyne et al. 2004). Cleavage of SPARC by certain matrix metalloproteinases increases the affinity of SPARC for collagens I, IV, and V (Sasaki et al. 1997), and such activity may serve to direct SPARC to sites of ECM remodeling.
Examination of SPARC-null mice has revealed that these animals have osteopenia and decreased bone formation (Delany et al. 2000), as well as defects in collagen fibril formation that influence responses to insults such as dermal wounding (Bradshaw et al. 2001,2002), implants (Puolakkainen et al. 2003), and tumor cells (Brekken et al. 2003). Such studies support the hypothesis that SPARC has specific functions in specific sites and that regulated expression of SPARC is important during development and in the response to alterations in tissue homeostasis.
To explore the function and activity of SPARC in the homeostasis of intervertebral disc ECM, we have studied discs in SPARC-null mice. Specifically, we have asked whether SPARC affects the morphology of the disc and whether the absence of SPARC predisposes the disc to accelerated aging.
Materials and Methods
Animal studies were performed following approval by the Institutional Animal Care and Use Committees at Carolinas Medical Center and the Hope Heart Program. C57Bl/6 × 129/SvJ wild-type (WT) and SPARC-null mice were used in this study. The SPARC-null colony has been backcrossed onto a C57Bl/6 background, such that animals used in these studies were essentially a C57Bl6 strain. Mice were raised in a specific pathogen-free facility. Confirmation of complete ablation of SPARC expression has been shown by Northern blotting and immunoblotting (Norose et al. 1998). WT and SPARC-null mice, 1.5–21 months of age, were weighed and euthanized, and their spinal segments were removed for the radiological and morphological experiments described in the following sections.
Lateral radiographs of the lumbar spine were obtained with a MicroFocus Imaging system with high-resolution Kodak X-OMAT TL mammography film (Kodak; Rochester, NY). Specimens were fixed in either 10% neutral buffered formalin or 70% ethanol and were decalcified in a solution of 22.5% formic acid (Allegiance; McGaw Park, IL) and 10% sodium citrate (Sigma; St Louis, MO). Complete decalcification was determined by radiography. The spine was cut in sagittal section, embedded in paraffin, and sectioned at 4 μm. Sections were stained with Masson-trichrome for evaluation of general disc features and with picrosirius red-Alcian blue for evaluation of proteoglycan content (Gruber et al. 2002).
Adjacent sections were processed for immunohistochemical localization of collagens I, II, III, and VI. Paraffin sections were cut at 4 μm, collected on PLUS slides (Allegiance), dried at 60C, and deparaffinized in xylene (Allegiance). Sections were rehydrated through graded alcohol changes (AAPER; Shelbyville, KY) to distilled water. The immunohistochemistry procedure was performed with the Dako Autostainer Plus (DakoCytomation; Carpinteria, CA) automated stainer. Endogenous enzyme was blocked using 3% H2O2 (Sigma) in methanol (Allegiance) for 5 min. Slides were rinsed with Tris-buffered saline containing 0.05% Tween 20 (TBST) (DakoCytomation) and were treated with 10% normal goat serum (Sigma). Normal sera were diluted in a solution of 5% BSA (Sigma) and 4% nonfat dried milk (Carnation; Young America, MN) in PBS, pH 7.4 (Roche, Indianapolis, IN). The blocking solution was blown off the slides without rinsing and the primary antibody was applied. All antibodies were purchased from Biodesign International (Saco, ME). Anticollagen I and II IgG were used at a 1:100 dilution, anticollagen III was used at a 1:200 dilution, and anticollagen VI IgG was used at a 1:400 dilution. All antibodies were diluted in 10% normal goat serum. Ten percent normal goat serum was used as a negative control. Slides were incubated in primary antibody for 90 min, rinsed in TBST, and treated with biotinylated goat anti-rabbit IgG (Vector Laboratories; Burlingame, CA) diluted 1:50 in 10% normal goat serum for 1 hr. Slides were rinsed in TBST and were treated with streptavidin conjugated horseradish peroxidase (DakoCytomation) for 20 min. Slides were rinsed in TBST and treated with the Chromagen Vector NovaRed (Vector Laboratories) for 5 min. Slides were rinsed in water, removed from the Autostainer, counterstained with Light Green (Polysciences; Warrington, PA), dehydrated, cleared, and mounted with Cytoseal XYL (Allegiance).
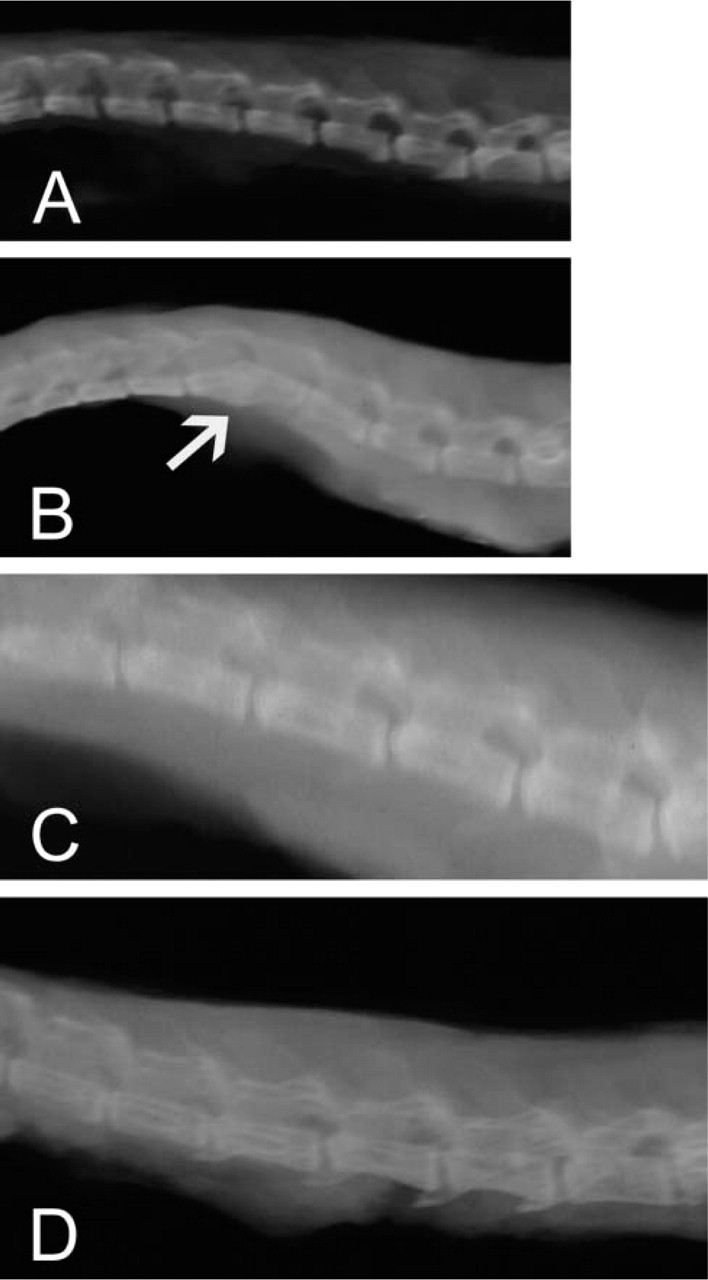
Radiographic characteristics of the lumbar spine of SPARC-null and wild-type (WT) mice. Lateral radiographs of the lumbar spine of WT mice at ages 2 (
Light microscopy was performed on the lumbar spines of the following WT animals: 11 mice aged 0.3–2.9 months, 15 3-month-old mice, 8 mice aged 4.5–5 months, and 6 mice aged 6–20 months. The following SPARC-null mice were studied: 16 mice aged 0.3–2.9 months, 12 3-month-old mice, and 15 mice aged 6–21 months.
Histomorphometry was performed on the annulus to determine cell densities in WT and SPARC-null mice. Quantitative histomorphometry was performed on tissue sections stained with Masson-trichrome dye using the OsteoMeasure system (OsteoMetrics; Atlanta, GA).
For ultrastructural studies, discs from two WT and two SPARC-null mice were minced, immersed in Karnovsky's fixative, postfixed with osmium tetroxide, embedded in Spurr resin, thin-sectioned, and grid-stained with uranyl acetate and lead citrate. Sections were viewed on a Philips CM10 transmission electron microscope. Analysis of collagen fibril diameters was performed with the OsteoMeasure system (OsteoMetrics).
Standard statistical methods were performed with SAS version 8.2 (SAS; Carey, NC). Data are presented as mean ± SD (n).
Results
Spinal Radiology and Morphology
Spines from SPARC-null and WT mice at 1.5–21 months of age were examined in lateral radiographs. Wedging, disc space narrowing, endplate calcification, endplate sclerosis, and irregular vertebral endplate margins (all features seen in human lumbar discs during degeneration) were present in the lower lumbar discs of the older SPARC-null mice.
As shown in Figure 1A, in a 2-month-old WT animal, disc spaces were regular and showed no wedging or endplate calcification. In contrast, radiologic images of 2-month-old SPARC-null mice showed wedging, endplate calcification, and sclerosis. Figure 1B shows loss of disc space and spontaneous fusion (arrow). As the SPARC-null mouse ages, radiological signs of degeneration appeared earlier compared with age-matched WT spines. Note that the lumbar spine of the 6-month-old SPARC-null mouse in Figure 1D showed substantially more prominent wedging and endplate calcification than observed in the spine of an 11-month-old WT mouse (Figure 1C).
Figure 2 shows histological images of the midsagittal spine. At 2 months of age, the mouse's WT disc spaces have relatively uniform widths at the dorsal and ventral margins (Figure 2A). In the 2-month-old SPARC-null mouse, however, there is marked disc wedging (the ventral portion of the disc is much wider than the dorsal portion) (Figure 2C).
Immunostaining for type I collagen resulted in only faint localization in the ventral annulus in SPARC-null discs (Figure 3B) in comparison to the normal, abundant immunoreactive product present in WT discs (Figure 3A). No immunohistochemical differences between WT and SPARC-null discs were found with respect to types II, III, and VI collagen (data not shown).
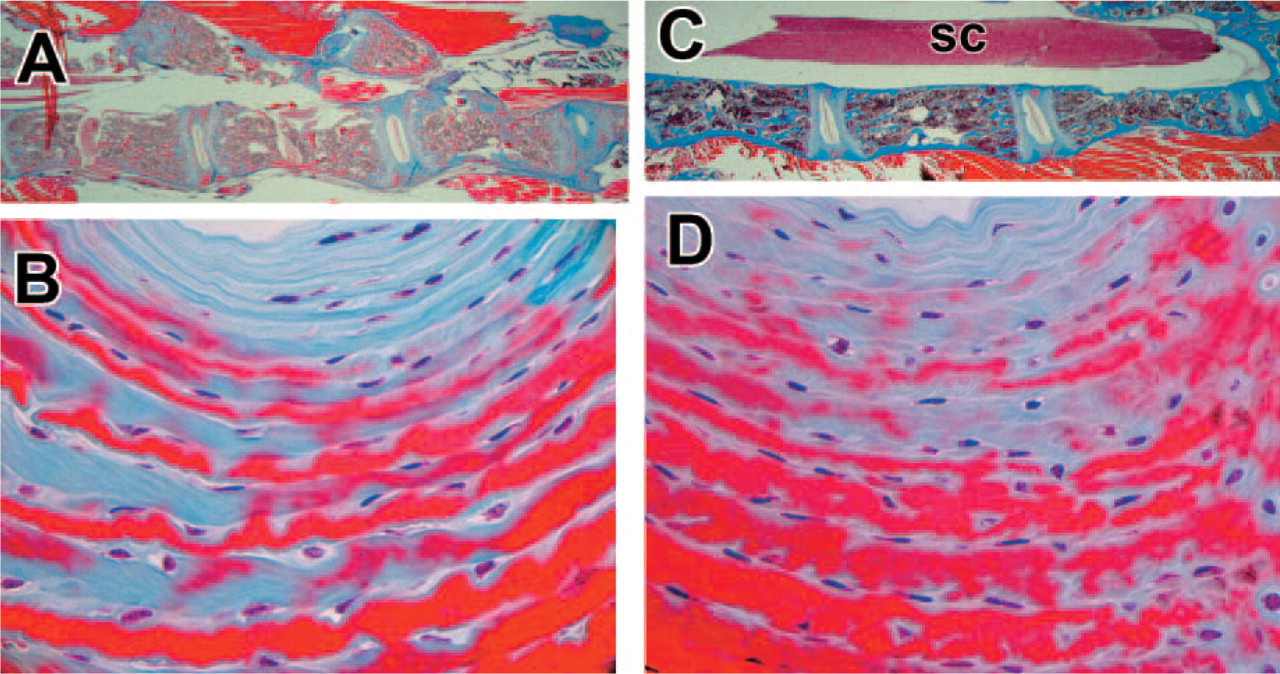
Histologic characteristics of the lumbar spine of SPARC-null and wild-type (WT) mice. Orientation of sections: The top of each figure is dorsal; the bottom is ventral. (
Immunohistochemical reactivity of anti-type I collagen antibody on discs from SPARC-null and wild-type (WT) mice. A 3-month-old WT mouse (
Cell densities in the annulus of WT and SPARC-null annuli from animals aged 3 and 19 months were measured by histomorphometry (Table 1). In WT discs, cell densities did not change from 3 to 19 months. SPARC-null discs, in sharp contrast, contained greater number of cells at 3 months, and subsequently exhibited a substantial loss of cells by 19 months. Statistical analysis using ANOVA revealed that there were both significant effects of time (i.e., aging) (p = 0.017) and a statistically significant interaction between the SPARC-null state (“treatment” in the ANOVA analysis) and time (p = 0.034). These findings are important because cell loss in the SPARC-null disc is expected to contribute to impaired ECM maintenance and remodeling and could explain the disc herniations described in the following section in the SPARC-null aging spine.
Characterization of Intervertebral Discs
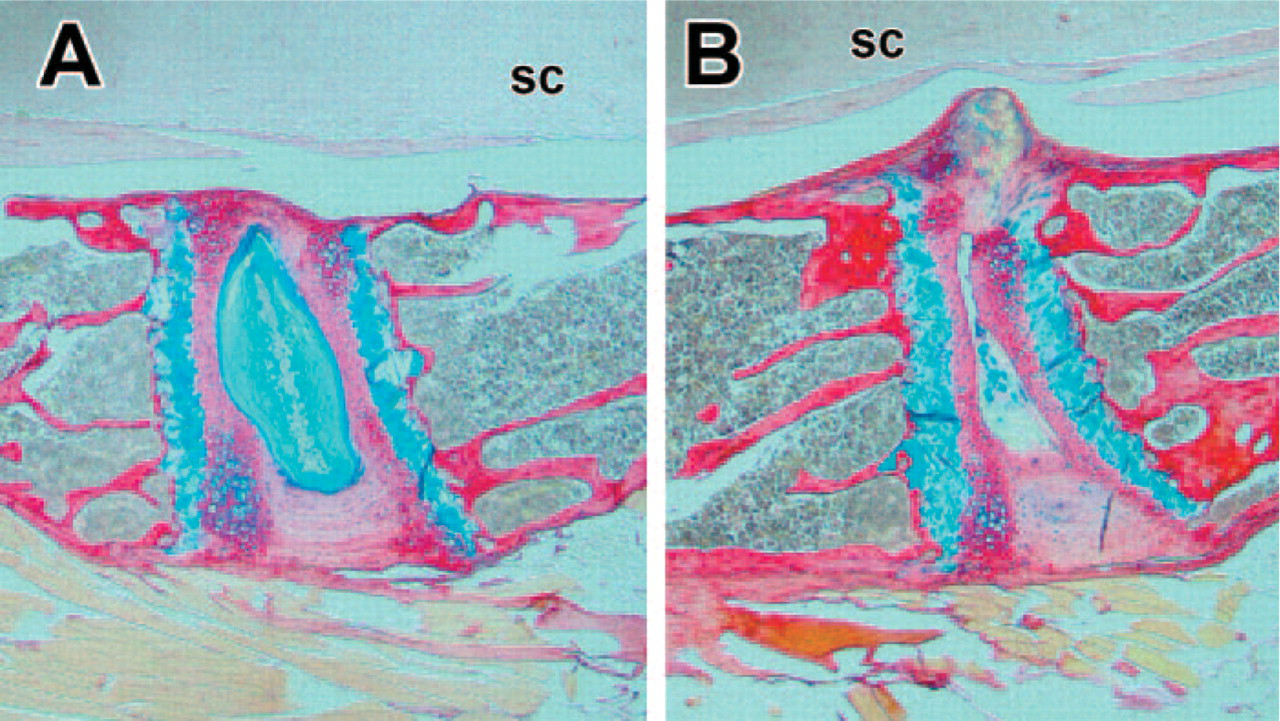
No herniations have been identified in any of the WT discs. In SPARC-null specimens, however, histological evidence of herniated discs has been found in mice aged 14, 19, and 20 months (Figure 4). These herniations projected dorsally from the disc and often appeared to impinge on the spinal cord. Multiple herniated discs have been observed in the same spine (as shown in Figures 4B and 4C, which depict adjacent discs).
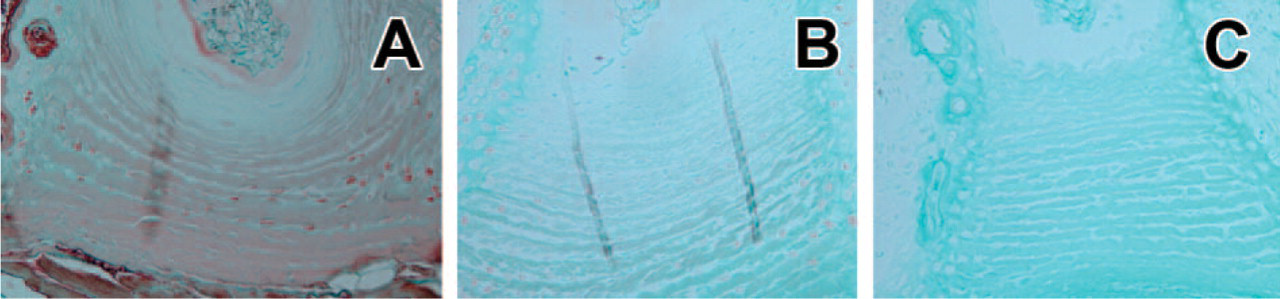
The herniated discs displayed an apparently decreased content of proteoglycan, in comparison to that of WT counterparts (Figure 5B vs 5A).
Ultrastructural Characterization of Fibril Morphology
In the WT annulus, collagen fibrils examined in cross-section (Figure 6A) and longitudinal section (Figure 6B) were uniform in diameter and exhibited a regular fibril margin. In marked contrast, fibrils from SPARC-null discs showed a wide variety of sizes and diameters (Figures 6C and 6D) and often had irregular margins reminiscent of irregular packing or assembly (e.g., “spiraled collagen” or “helical collagen” [Ghadially 1997]).
Although there was no difference in mean fibril diameter in the annulus of WT vs null discs (WT 57.67 ± 11.898 [103] vs null, 56.07 ± 20.35 [151]), chi-square analysis showed that there was a significant difference in the pattern of size distributions of the fibrils when SPARC-null collagen diameters were compared with WT diameters (p<0.0001) (Figure 7). Both smaller and larger diameter fibrils were present in null discs (null range: 13.72–115.16 nm; WT range: 56.99–91.92 nm). The majority (21%) of fibrils from discs of null animals were 41–50 nm in diameter. Although 40.8% of the fibrils in the WT annulus were 51–60 nm in diameter, only 21% of the null annulus fibrils were of that size.
Discussion
SPARC is a calcium-binding protein secreted by a number of cells, including fibroblasts, endothelial cells, and osteoblasts (Stenner et al. 1986; Sage et al. 1989a; Kasugai et al. 1991; Vuorio et al. 2003). SPARC is expressed in many tissues during development, whereas in the adult, it is more prominent in renewing populations of cells (Sage et al. 1989b), in tissues undergoing remodeling as a result of injury or disease, and during bone formation, adipogenesis, and angiogenesis (Chiquet-Ehrismann 1991; Sage and Bornstein 1991; Reed et al. 1993; Puolakkainen et al. 1999; Bradshaw and Sage 2001; Bornstein and Sage 2002). SPARC has been shown to induce the expression of matrix metalloproteinases (collagenase) in fibroblasts (Tremble et al. 1993), and in vertebrates it binds a number of ECM components (such as the fibrillar collagens [types I, II, III, and V] and type IV [basement membrane] collagen), and is believed to regulate cytokine expression, as exemplified by its effect on transforming growth factor (TGF-β1) in mesangial cells (Francki et al. 1999; Bradshaw and Sage 2001).
Analysis of the number of cells/mm2 at 3 and 19 monthsa

Data are expressed as mean ± SD (n) where n = number of discs evaluated. Data were analyzed with two-way ANOVA. WT, wild-type; SPARC, secreted protein, acidic, and rich in cysteine; NS, not significant.
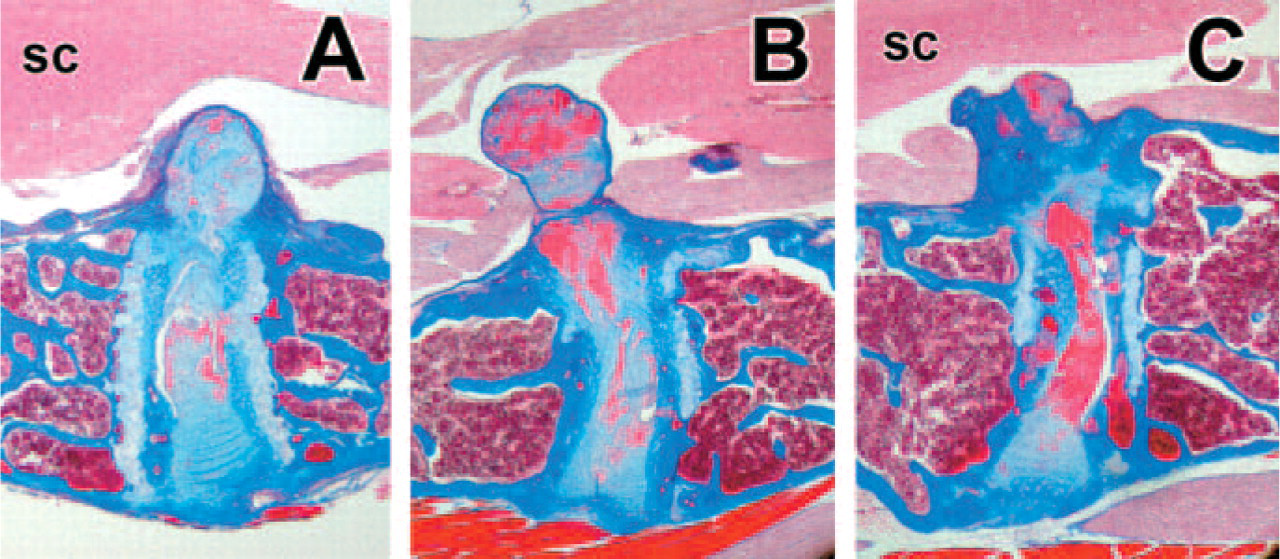
Morphological evidence of herniations in the lumbar discs of SPARC-null mice. No herniations have been observed in spines of wild-type mice. Discs in SPARC-null mice showed prominent histologic evidence of herniations in SPARC-null spines (as shown here, often in adjacent discs). sc, spinal cord; Masson-trichrome stain. (x20)
Mice with targeted deletions of the matricellular proteins are valuable research models. Deletions of most of the matricellular proteins studied to date result in either grossly normal or subtle phenotypes that are exacerbated during injury. Framson and Sage (2004) have recently reviewed the major phenotypic characteristics of SPARC-null mice. These mice exhibit defects in the lens capsule associated with early cataract formation (Yan et al. 2002), osteopenia (Delany et al. 2000), reduction of the dermis, with small collagen fibrils and decreased tensile strength of the skin (Bradshaw et al. 2003b), and increased adipose tissue (Bradshaw et al. 2003a). Our data contribute to the number and type of tissues adversely affected by the absence of SPARC to include alterations of the ECM of the intervertebral disc associated with accelerated disc degeneration during aging.
Distribution of proteoglycan in intervertebral discs revealed by picrosirius red-alcian blue (Gruber et al. 2002). Proteoglycan is stained blue, and collagen is stained red. Abundant proteoglycan is present in the nucleus of the disc from a 19-month-old SPARC-null mouse (
The SPARC-null mouse also exhibits an altered response to injury, reflected in accelerated closure of cutaneous wounds (Bradshaw et al. 2002), diminished encapsulation of implanted materials (Puolakkainen et al. 2003), reduced tumor growth of mammary carcinoma (Sangaletti et al. 2003), and enhanced growth and metastasis of Lewis lung carcinoma and B-cell lymphoma (Brekken et al. 2003). Such changes in the SPARC-null phenotype reflect the significant role played by SPARC in the design, maintenance, and repair of a diversity of tissues types. As pointed out by Framson and Sage (2004), the common theme revealed by these defects is that SPARC plays a critical role in the production or assembly of the ECM. The results presented here indicate that discs from young SPARC-null mice can meet the structural and remodeling demands made on them, but with aging and loss of disc cells, the ECM fails, with resultant herniations.
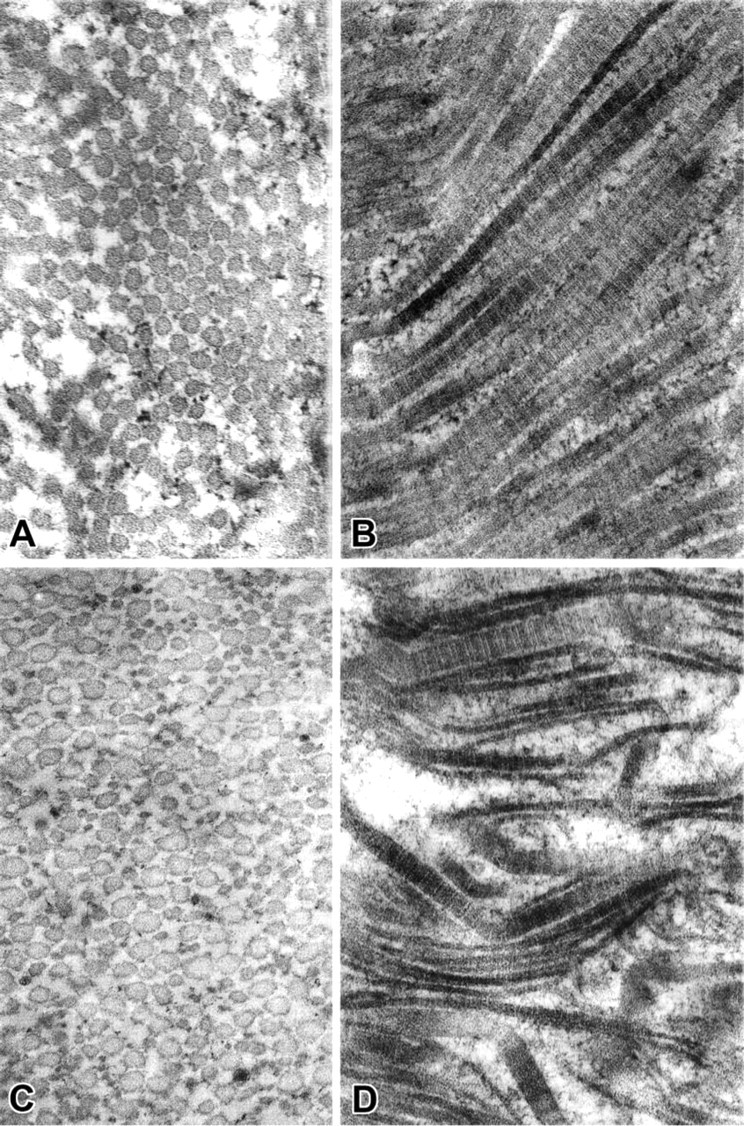
Ultrastructural characteristics of collagen fibrils in the disc annulus In contrast to uniform diameters of fibrils in cross-section (
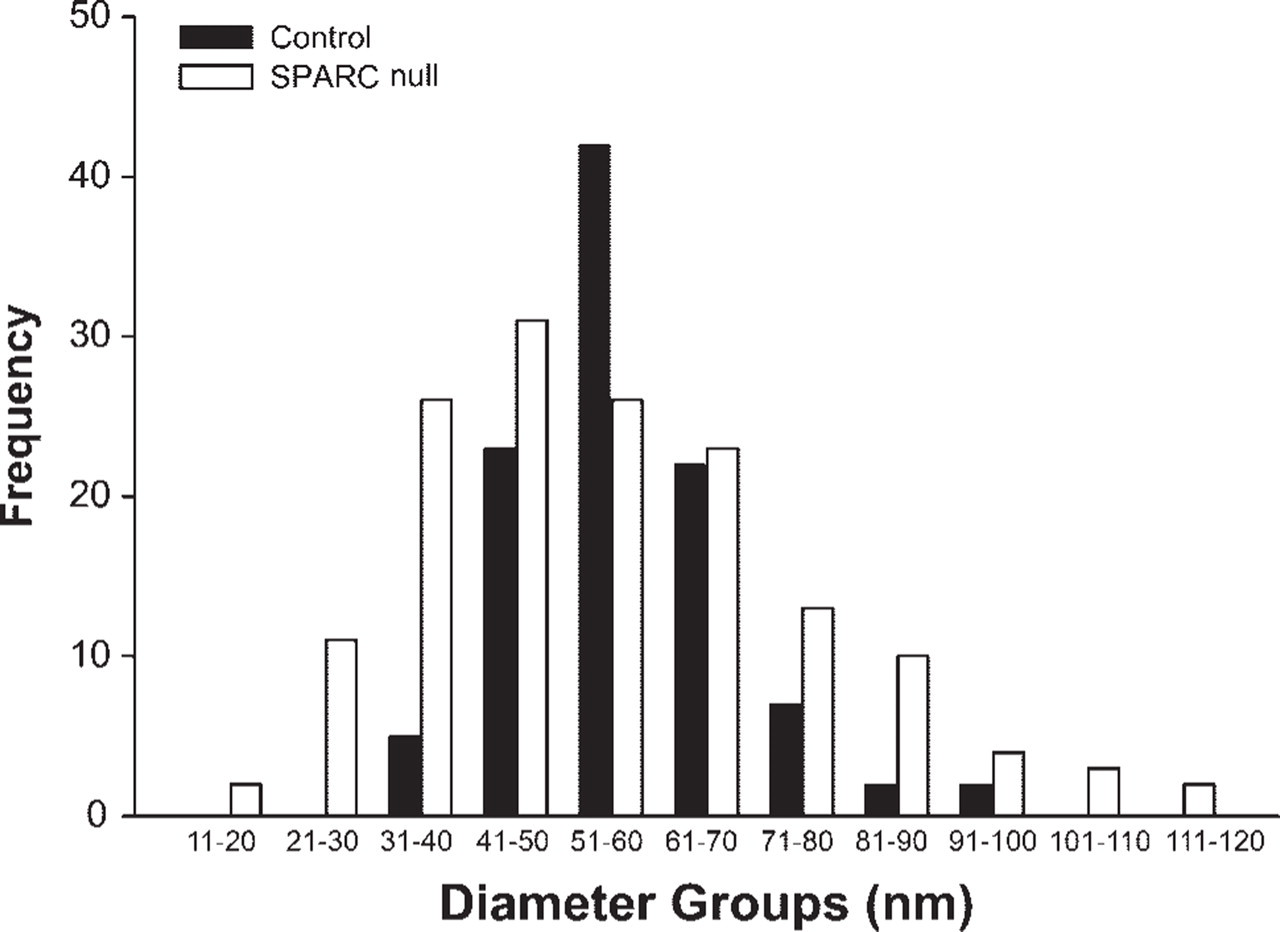
Distribution profiles of fibril diameters. Fibril diameters were measured, and distribution profiles of widths were subsequent calculated. Fibril diameters in discs of SPARC-null animals (gray bars) showed a distribution profile that differed significantly from that of fibril diameters in wild-type discs (black bars). (See text for statistical analysis.)
From what is known of the role of SPARC in other tissues, our data are consistent with several possible mechanisms that could account for the disc ECM failure in SPARC-null mice. One candidate is the substantial downregulation of type I collagen that is seen in vitro and is reflected in alterations such as the thin dermis of mice lacking SPARC (Bradshaw et al. 2003b). TGF-β is a recognized mitogen for disc cells (Gruber et al. 1997), and it is known that SPARC affects cellular levels of TGF-β, its receptor activation, and components of its signal transduction (Francki et al. 1999; Schiemann and Schiemann 2003).
The decreased proteoglycan content seen in herniated discs was an important finding, consistent with observations in human disc aging and degeneration wherein glycosaminoglycan content decreases with age (Antoniou et al. 1996). Because proteoglycans bind water and contribute to the “shock absorber” function of the disc, the changes in biochemical disc composition observed here in herniated discs of SPARC-null mice probably have significant functional consequences, and further work is needed to investigate proteoglycan loss in discs of the SPARC-null mouse.
In summary, the accelerated degeneration in discs from SPARC-null mice indicates that SPARC has an essential role in normal disc ECM remodeling and matrix homeostasis. The absence of SPARC adversely affects: (a) the number of disc cells in older animals, (b) the ultrastructure of collagen fibrils (evidenced by alterations in fibril diameter and shape in both young and old mice), (c) type I collagen content (apparent reduction in the annulus of both young and old mice), (d) spinal structure (apparent from radiologic wedging and endplate changes in young and old mice), and (e) the structural integrity of the disc (seen in the development of disc herniations in older mice).