
Abstract
Galectin is an animal lectin that recognizes β-galactosides of glycoconjugates and is abundant in the gut. This study revealed the cellular expression of galectin subtypes throughout the mouse digestive tract by in situ hybridization. Signals for five subtypes (galectin-2, -3, -4/6, and -7) were detected exclusively in the epithelia. In the glandular stomach, galectin-2 and -4/6 were predominantly expressed from gastric pits to neck of gastric glands, where mucous cells were the main cellular sources. The small intestine exhibited intense, maturation-associated expressions of galectin-2, -3, and -4/6 mRNAs. Galectin-2 was intensely expressed from crypts to the base of villi, whereas transcripts of galectin-3 gathered at villous tips. Signals for galectin-4/6 were most intense at the lower half of villi. Galectin-2 was also expressed in goblet cells of the small intestine but not in those of the large intestine. In the large intestine, galectin-4/6 predominated, and the upper half of crypts simultaneously contained transcripts of galectin-3. Stratified epithelium from the lip to forestomach and anus intensely expressed galectin-7 with weak expressions of galectin-3. Because galectins in the digestive tract may be multi-functional, information on their cell/stage-specific expression contributes to a better understanding of the functions and pathological involvements of galectins.
G
It is generally noted that the galectin subtypes display cell- and tissue-specific distributions except for galectin-1, which is ubiquitously present as a stromal galectin in mammalian tissues (Poirier et al. 1992) and interacts with laminin (Cooper et al. 1991; van den Brule et al. 1995) and fibronectin (Ozeki et al. 1995). The mammalian digestive tract is one of the organs rich in galectin, and six subtypes of galectin (galectin-2, -3, 4, -6, -7, and -9) have been detected by several assay systems. Oka et al. (1999) showed an intense expression of rat galectin-2 mRNA in the stomach and small intestine and a less intense expression in the large intestine by Northern blot and reverse transcriptase polymerase chain reaction (RT-PCR) analyses. By immunostaining, they documented the presence of galectin-2 immunoreactivity only in the glandular epithelial cells of the rat stomach, possibly some population of parietal cells (Oka et al. 1999). Galectin-3, formerly described as the Mac-2 antigen, carbohydrate-binding protein (CBP)-30/35, and IgE-binding protein, was identified in cell lineages of hematopoietic origin such as macrophages (Cherayil et al. 1989; Woo et al. 1990) and mast cells (Craig et al. 1995). In the human and bovine intestine, immunostaining for galectin-3 detected the immunoreactivity in epithelia of the duodenum and colon (Sanjuan et al. 1997; Kaltner et al. 2002). Galectin-4 is a subtype unique to the gastrointestinal tract (Oda et al. 1993; Gitt et al. 1998). Northern blot analyses showed that the galectin-4 mRNA expression was intense in the small and large intestine but much less so in the stomach of the rat (Oda et al. 1993) and mouse (Gitt et al. 1998). Immunohistochemical studies for galectin-4 have provided fragmental information on cellular localization such as the stratified epithelium of the oral cavity and esophagus in pigs (Chiu et al. 1992), the esophageal epithelium in rats (Wasano and Hirakawa 1995), small intestinal epithelium in pigs (Danielsen and van Deurs 1997), and the colonic epithelium in humans (Huflejt and Leffler 2004). Galectin-6 is highly homologous to galectin-4 (83% in amino acid sequence, 93% in nucleotide sequence) but so far has only been found in mice and probably arose from a recent duplication during evolution. Because of this similarity, it is difficult to distinguish galectin-6 from galectin-4, and they are consequently detected together in most antibody-based and nucleotide hybridization assays (Leffler 1997). Accordingly, their combined expression was briefly noted in the intestinal epithelium of mice by in situ hybridization analysis (Gitt et al. 1998). Galectin-7 was originally cloned from the human epidermis as a molecular marker for keratinocyte differentiation (Madsen et al. 1995; Magnaldo et al. 1995). Immunohistochemically, human and mouse galectin-7 is distributed in stratified epithelial cells of the esophagus, tongue, and lip, as well as the epidermis (Magnaldo et al. 1998; Sato et al. 2002a). Galectin-9, previously described as ecalectin, possesses C-terminal CRD highly homologous (94% in cDNA) to that of galectin-5, which was found in rat erythrocytes (Gitt et al. 1995) but is not present in the mouse (Wada and Kanwar 1997). Wada and Kanwar (1997) reported the expression of mouse galectin-9 mRNA in the small intestine, liver, thymus, and other tissues by Northern blot analysis, although the types of expressing cells have not been investigated.
The question therefore arises as to whether these family members show an overlapping or differential expression in the gastrointestinal tract or follow any rules in their combined expression. To answer this question, a systematic analysis of galectin expression is needed at cellular levels. In the present study, we identify the cell types of galectin-expressing cells in the mouse digestive tract by in situ hybridization using oligonucleotide probes specific for each of the predominant galectin subtypes (galectin-1 to galectin-7 except for galectin-5). The method used in this study has an advantage in its specific identification of subtypes with a higher homology and in its visual comparison of signal intensity. Furthermore, it is superior to immunohistochemistry when the proteins in question are highly water soluble (Huflejt et al. 1997) and tend to condense onto extracellular components after secretion (Wasano and Hirakawa 1995, 1999).
Materials and Methods
Animals and Tissue Sampling
Eight-week-old, adult male ddY mice (Japan SLC; Shizuoka, Japan) were used in the present study. The mice were killed by bloodletting from the heart under deep anesthesia with pentobarbital, and fresh tissues were obtained from various regions of the gastrointestinal tract from the lip to the anus. The whole stomach and the duodenum, 0.5 cm in length from the pyloric orifice, were removed together and divided into two pieces: the fore/glandular stomach and the pyloric antrum to duodenum. The rest of the small intestine was double folded, and the middle regions of both the upper and lower parts were obtained as the jejunum and ileum, the latter of which was selected to include Peyer's patches. Three different parts of the large intestine were dissected from the middle parts of the cecum and proximal colon and from the rectum with anus. These tissues were washed sufficiently with physiological saline and embedded into a freezing medium (OCT compound; Sakura Finetechnical Co. Ltd., Tokyo, Japan). The salivary glands, pancreas, and liver were also collected and embedded in OCT compound. These embedded tissues were quickly frozen in liquid nitrogen and stored until use. All experiments were performed under protocols following the Guidelines for Animal Experimentation, Graduate School of Medicine, Hokkaido University, Japan.
In Situ Hybridization
Two non-overlapping 45-mer antisense oligonucleotide probes specific for each of the mouse galectin mRNAs and for a proton pump (H+, K+-ATPase) and pepsinogen C were synthesized. The targeted nucleotide residues of probes used in this study are shown in Table 1. These oligonucleotides were labeled with 35S- or 33P-dATP using terminal deoxynucleotidyl transferase (TOYOBO; Osaka, Japan). Ten-μm-thick fresh frozen sections were prepared and mounted on glass slides precoated with 3-aminopropyltriethoxysilane. They were fixed with 4% paraformaldehyde in 0.1 M phosphate buffer for 15 min and then acetylated with 0.25% acetic anhydride in 0.1 M triethanolamine-HCl (pH 8.0) for 10 min. Hybridization was performed at 42C for 10 hr by adding 10, 000 cpm/ μl of 35S- or 33P-labeled oligonucleotide probes. Slides were rinsed at room temperature for 30 min in 2× SSC (1× SSC: 150 mM sodium chloride, 15 mM sodium citrate) containing 0.1% sarkosyl, twice at 55C for 40 min in 0.1× SSC containing 0.1% sarkosyl, dehydrated through a graded series of ethanol, and air dried. Sections were either exposed to BioMax MR films (Kodak; Rochester, NY) for a week or dipped into autoradiographic emulsion (NTB-2; Kodak) and exposed at 4C for 4–8 weeks. The hybridized sections were counterstained with hematoxylin after development. Some hybridized or non-hybridized sections adjacent to hybridized ones were stained with periodic acid/Schiff reaction (PAS) to identify mucous cells expressing galectin.
In situ hybridization using two non-overlapping antisense probes exhibited consistent labeling in all tissues examined. Specificity of the hybridization was also confirmed by the disappearance of the signals with an excess dose of unlabeled antisense probes.
Results
The intense and consistent expressions of galectin-2, -3, -4/6, and -7 mRNAs were recognized in the epithelium of the murine digestive tract, whereas mRNA expressions for galectin-1 were negligible in the epithelia. Weak signals for galectin-1 were detected diffusely in the lamina propria mucosae and the muscle layer. Because of the extremely high sequence homology between galectin-4 and galectin-6, we failed to design antisense probes that could specifically detect each of them. Thus, their combined expression (galectin-4/6) was investigated using antisense probes designed for galectin-6. No significant signals for any galectin subtypes examined were detectable in the salivary glands, pancreas, or liver.
Observation of X-ray films revealed region-dependent expressions of galectin mRNAs in the mucosal layer from the lip to anus (Figure 1). Galectin-2 mRNA was expressed intensely in the glandular stomach and weakly in the pyloric antrum (Figure 1A). The small intestine from the duodenum to ileum showed an intense expression of galectin-2 mRNA, but the large intestine lacked the transcripts of this subtype (Figure 1A). In contrast, galectin-3 mRNA expression was intense in the large intestine including the cecum, colon, and rectum, whereas the stomach and small intestine had weak signals for galectin-3 that slightly increased in intensity toward the ileum (Figure 1B). Signals for galectin-3 mRNA were also observed in the stratified epithelium of the lip, tongue, esophagus, forestomach, and of the anus (Figure 1B), though much weaker than those of galectin-7. An intense expression of galectin-4/6 mRNAs was widely distributed from the glandular stomach to rectum, being especially intense in the large intestine (Figure 1C). Faint signals for galectin-4/6 were also found in the forestomach (Figure 1C). Galectin-7 mRNA expression was characterized by a restricted distribution in the stratified epithelia from the lip to forestomach, and of the anus (Figure 1D).
Light microscopic observations of hybridized sections revealed the detailed expression of each galectin subtype mRNA; the expression of all subtypes except for galectin-1 was confined to the epithelium in all regions of the gastrointestinal tract. The predominant subtype expressed in the glandular stomach was galectin-2 and -4/6, followed by galectin-3. An intense expression of galectin-2 mRNA was found in the upper half of the gastric mucosa, namely, from gastric pits to the neck of gastric glands (fundic glands) excluding the surface region of gastric pits, which lacked significant signals (Figures 2A and 2C). Signals for galectin-4/6 showed a similar distribution to galectin-2 but extended to the surface region of gastric pits (Figures 2B and 2D). Weak but significant signals for galectin-3 were found only at the surface region of gastric pits (Figure 2E). The galectin-2 and -4/6 mRNA-expressing cells in the parietal cell area displayed a mosaic pattern (Figures 2C and 2D) and were complementary to H+, K+-ATPase mRNA-expressing parietal cells as shown by alternative staining of adjacent sections (Figures 2F and 2G). Mature chief cells, which expressed pepsinogen C mRNA, also failed to contain any transcripts for galectin subtypes (Figure 2H). These findings indicate that galectins in the stomach are produced by surface mucous cells, mucous neck cells, and undifferentiated cells at the isthmus of gastric glands. The pyloric antrum also contained signals for galectin-2, -3, and -4/6, all of which were localized in gastric pits containing PAS-positive mucous substance, but these signals were absent in pyloric glands with a weak PAS reaction (Figure 3). There were some differences in the detailed localization of these three galectin subtypes in gastric pits of the antrum. The expression of galectin-2 mRNA was free from the surface region of gastric pits (Figure 3B). On the other hand, the signals for galectin-3 were distributed mainly at the surface region of gastric pits (Figure 3D), and the signals for galectin-4/6 were found throughout gastric pits (Figure 3C). This expression pattern is essentially identical to that in the acid-secreting area of the glandular stomach.
Targeted nucleotide residues of probes used in this study

Exhibiting an extremely high sequence homology to the nucleotide residues 275–328 and 785–828 of galectin-4 (BC011236).
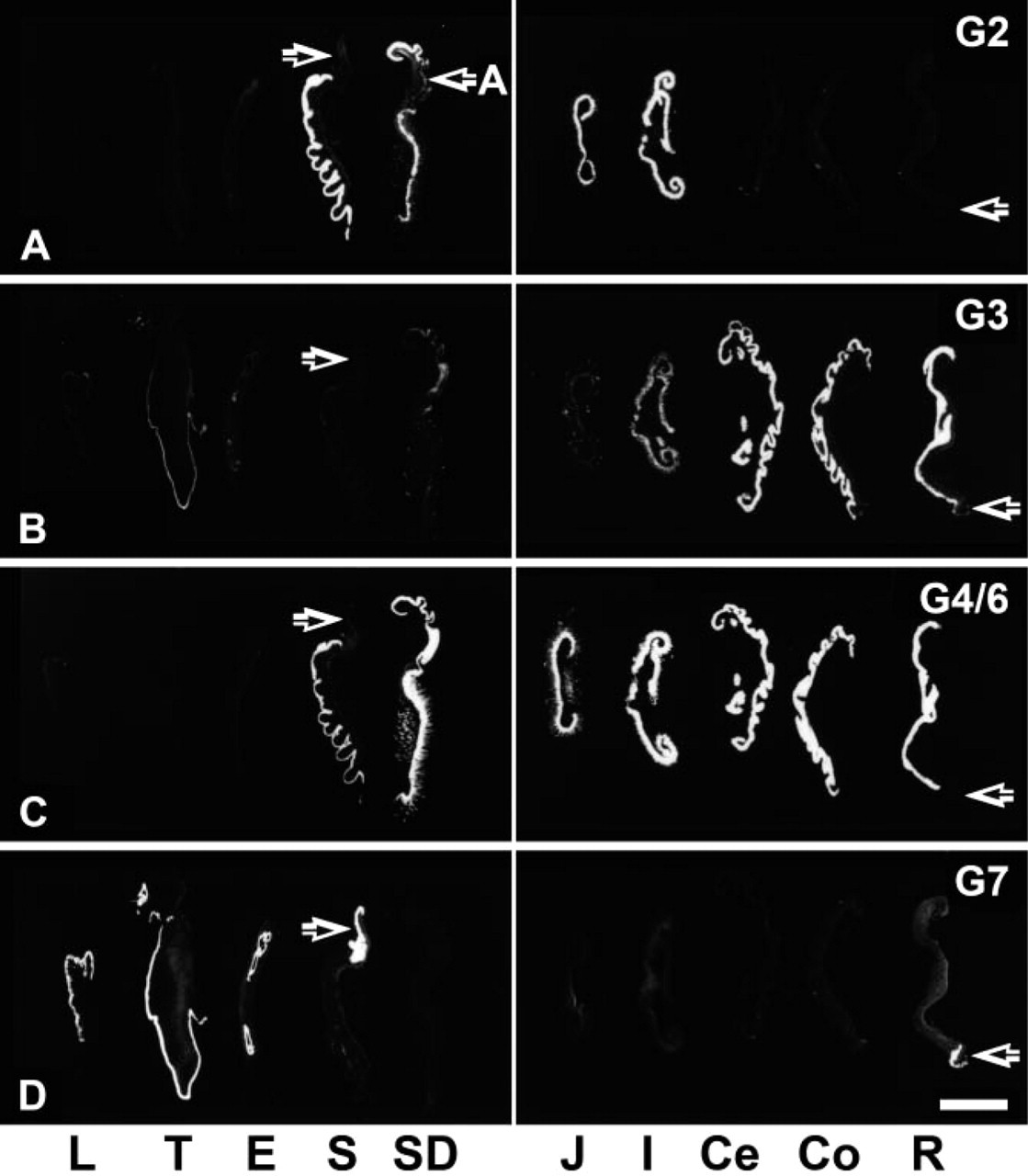
Observation of X-ray films for galectin mRNA expression. (
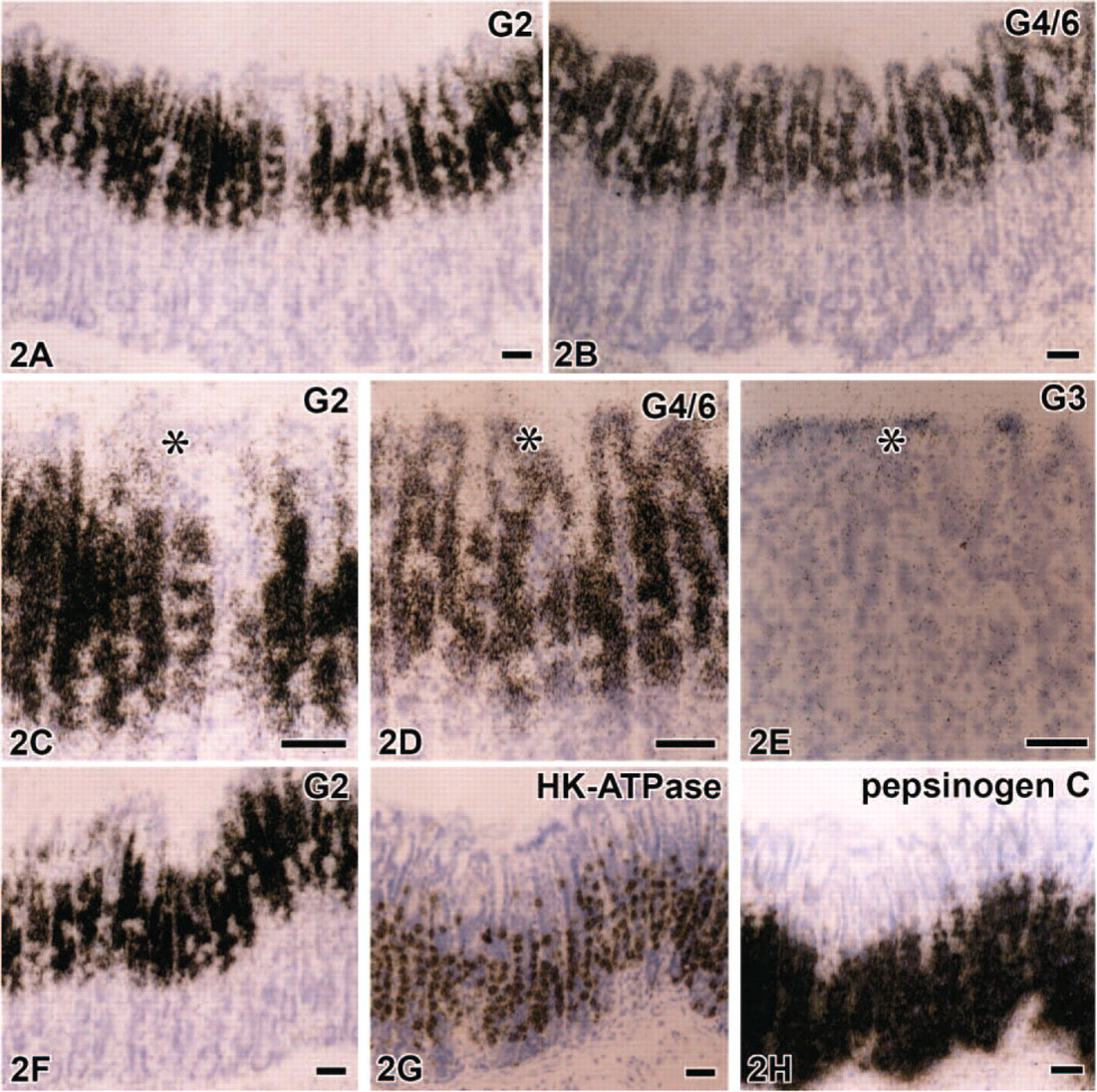
Predominant subtypes expressed in the glandular stomach are galectin-2 in
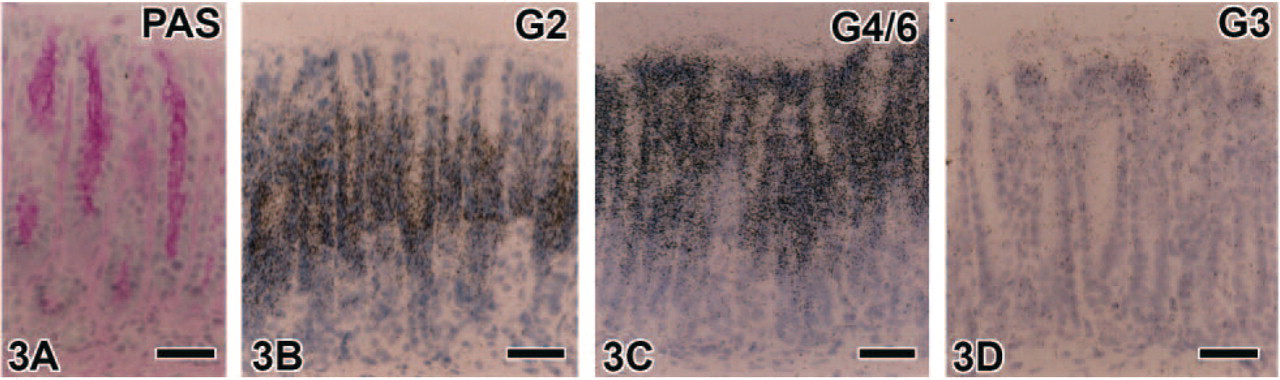
(
The small intestine intensely expressed mRNAs of galectin-2, -3, and -4/6 with the same combination of subtypes as the stomach, and their expression patterns were consistent from the duodenum to ileum. The signals for these subtypes were distributed from the upper half of crypts to villous tips with distinct cellular localization, whereas the bottoms of crypts lacked any of the signals examined (Figures 4A–4C). Intense signals for galectin-2 were recognized in the upper half of crypts and the base of villi (Figure 4A), and spotted signals were dispersed along the entire length of the villous epithelium (Figures 4A and 4D). When PAS staining was performed in the same sections used for in situ hybridization, the dispersed expression sites of galectin-2 mRNA were identified as PAS-positive goblet cells (Figures 4G and 4H). On the other hand, signals for galectin-3 in the small intestine gathered in the upper half of villi, being more intense at villous tips (Figures 4C and 4F). Galectin-4/6 mRNA expression was broadly distributed in the epithelium from the upper regions of crypts to villous tips (Figure 4B). The transcripts of galectin-4/6 in villi were most abundant in the basal one third and gradually decreased in intensity toward villous tips (Figure 4E). Small rounded cells basally located apart from the epithelial lining cells, possibly some population of leukocytes, showed condensed signals for galectin-4/6 (Figure 4I). The duodenal glands contained scattered signals only for galectin-4/6 (Figure 4J). Peyer's patches exhibited a moderately intense expression for galectin-4/6 and -3 at the covering (dome) epithelium (Figures 4K and 4L). Some galectin-2 mRNA-expressing cells were scattered at the epithelium covering Peyer's patches and identified as goblet cells by PAS staining (data not shown).
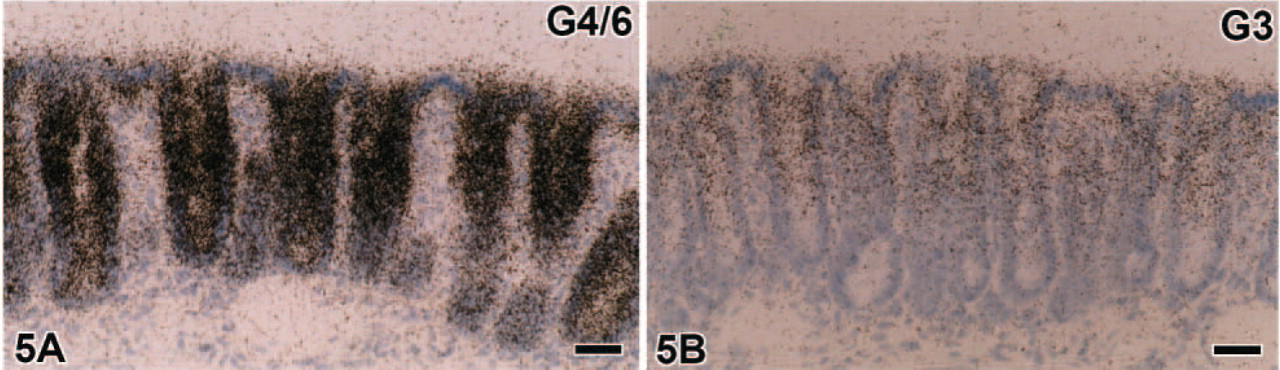
In the large intestine, galectin-3 and -4/6 were the predominant types (Figures 5A and 5B). Similar to the small intestine, galectin-3 mRNA expression in the large intestine occurred on the luminal side of mucosa, being intense in the upper half of crypts (Figure 5B), whereas signals for galectin-4/6 were broadly distributed in crypts but were less intense at their bottom (Figure 5A). The signals of galectin-3 and -4/6 in the large intestine tended to increase in intensity from the cecum toward the rectum. In contrast to goblet cells in the small intestine, those in the large intestine lacked any signals for galectin-2.
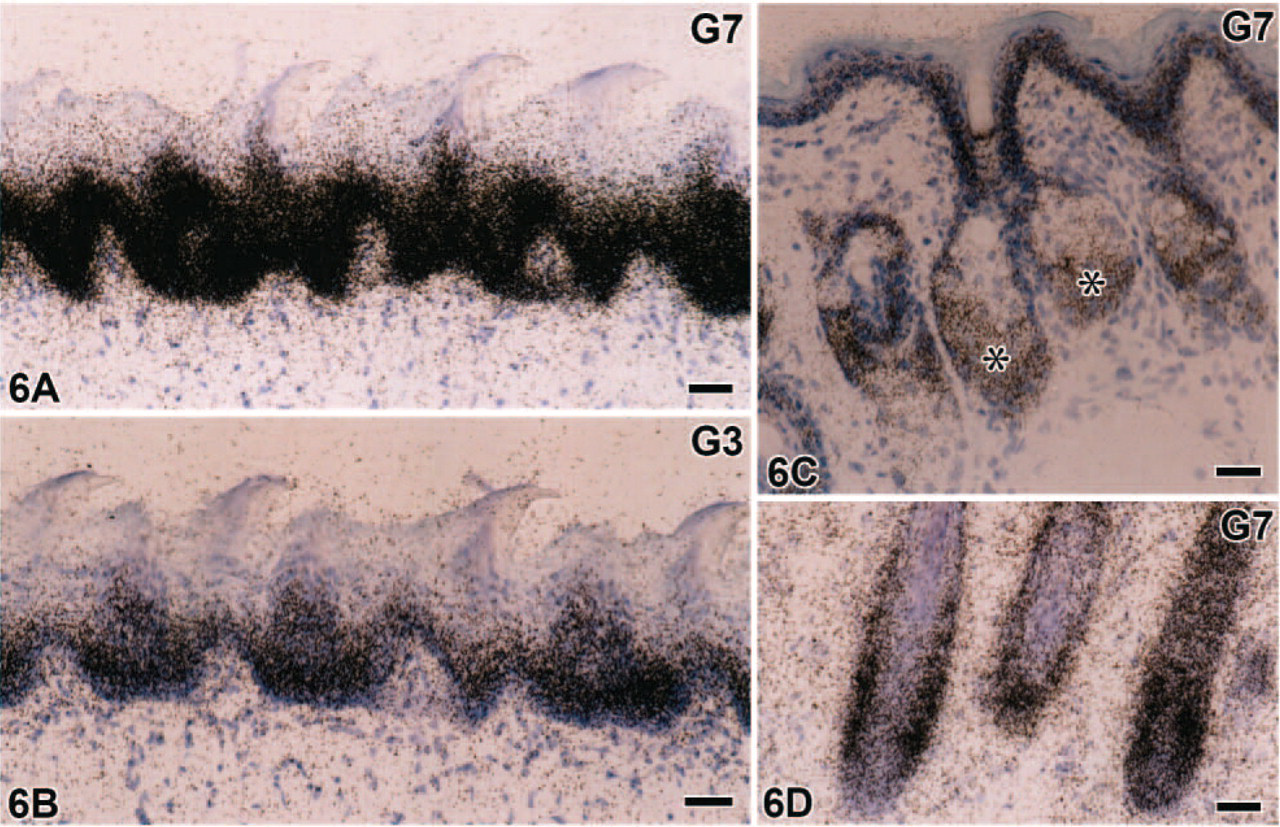
Stratified squamous epithelial cells from the lip (both mucosal and cutaneous sides) to the forestomach and those of the anus consistently expressed mRNAs of galectin-3 and -7, the latter of which predominated (Figures 6A and 6B). Weak signals for galectin-4/6 were found in the stratified epithelium of the forestomach (data not shown) but not in the other stratified epithelium examined. Signals for galectin-7 and -3 were preferentially recognized in keratinocytes at the lower half of the spinous layer, and galectin-7 expression extended to the basal layer (Figures 6A and 6B). Sebaceous glands and hair follicles in the lip and anus also contained abundant signals for galectin-7 (Figures 6C and 6D) and weak signals for galectin-3 and -4/6 (data not shown).
Discussion
Galectins comprise one family of animal lectins, are characterized by their ability to bind a lactose unit of glycoconjugates through preserved CRDs, and are extremely rich in the gastrointestinal tract. The present in situ hybridization analysis using X-ray films clearly demonstrated the region-dependent expression of galectin subtypes, namely, galectin-2, -3, -4/6, and -7, in the murine digestive tract with reference to the intensity of expression. Our findings concerning the regional distribution of these galectin subtypes largely correspond to previous data obtained by Northern and Western blottings or RT-PCR analyses (Gitt et al. 1998; Magnaldo et al. 1998; Oka et al. 1999; Sato et al. 2002a). In addition to the overlapping expression of galectin subtypes, we were able to show that every region of the gastrointestinal tract expresses any galectin subtype following set rules. Furthermore, the distinct cellular localization of galectins assayed at the microscopic level may allow the discovery of subtype-specific functions.
Tissue distribution of galectin-2 in the digestive tract has been roughly analyzed by the RT-PCR method in the rat (Oka et al. 1999). Namely, the galectin-2 mRNA expression at tissue levels was intense in the stomach and small intestine but much less so in the large intestine. The present in situ hybridization study confirmed the intense expression of mouse galectin-2 mRNA in the stomach and small intestine but could not detect any significant signals in the large intestine. Although Oka et al. (1999) immunohistochemically identified galectin-2-expressing cells in the rat stomach as parietal cells, our galectin-2 mRNA-expressing cells were not identical to parietal cells characterized by expression of H+, K+-ATPase mRNA and instead corresponded to mucous cells distributed from gastric pits to the neck of gastric glands. The relation of galectin-2 to mucous secretion was also recognized in goblet cells of the small intestine. The lack of galectin-2 transcripts in goblet cells of the large intestine may be explained by differences in type or at least differences in the chemical component of mucus (More et al. 1987). On the other hand, the expression sites of galectin-2 mRNA contained the proliferating zones, as clearly shown in the glandular stomach and small intestine where mitotic and undifferentiated epithelial cells reside. Although this unique expression pattern suggests the involvement of galectin-2 in the mitosis and differentiation of gut epithelial cells, no information is available for the relation between galectin-2 and cell proliferation. Other galectin subtypes, rather than galectin-2, are reported to be potent growth regulators, including galectin-1 and galectin-3, which play a role in cell proliferation as positive or negative regulators (Liu 2000).
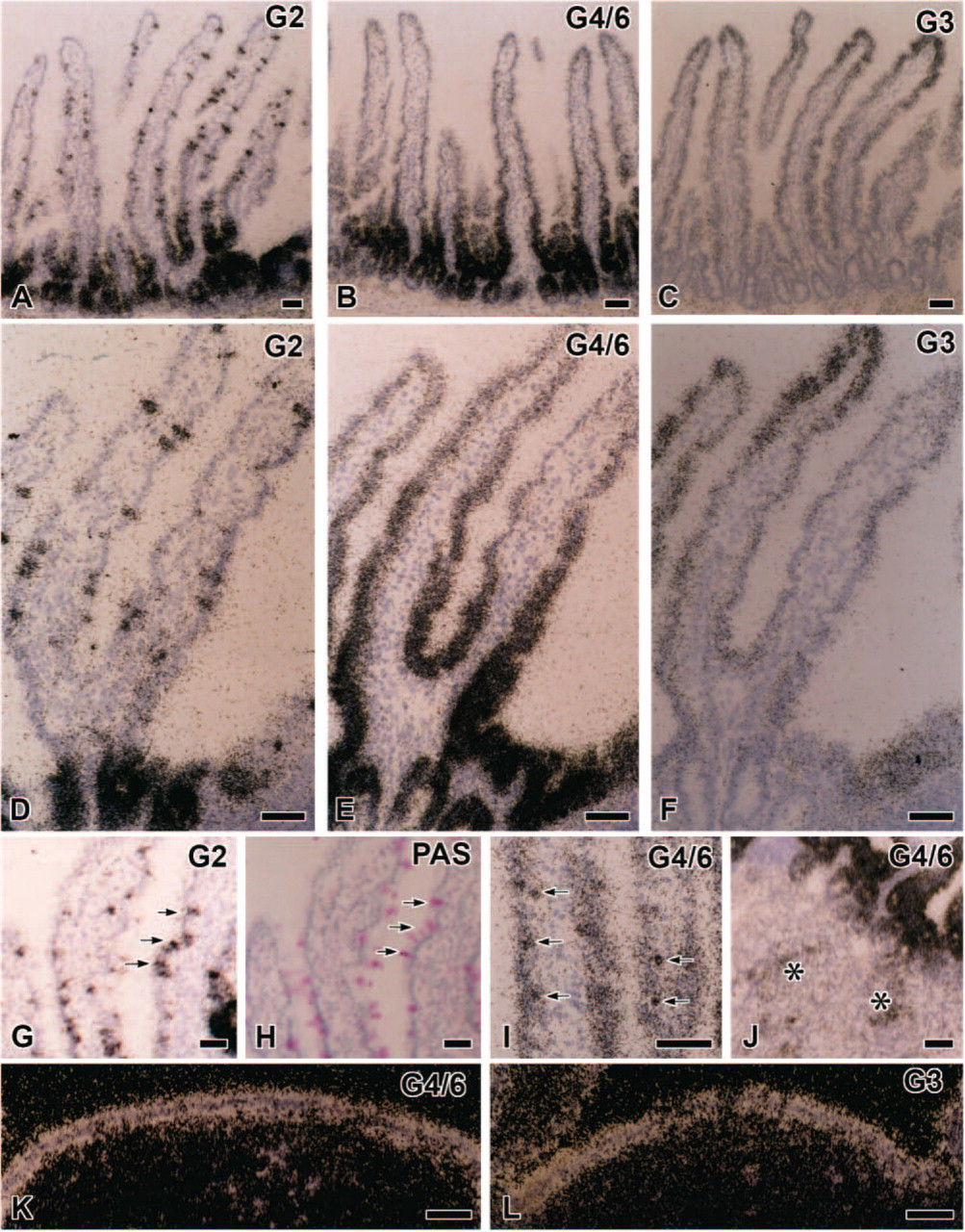
Galectin mRNA expression in the small intestine, ileum (
Galectin mRNA expression in the colon. (
Galectin mRNA expression in the tongue and lips. The stratified squamous epithelium, especially the basal half of the tongue, intensely expresses galectin-7 with a weak to moderately intense expression of galectin-3 (
In contrast to the active production of galectin-3 by leukocytes and its crucial roles in infections and inflammation (Almkvist and Karlsson 2004; Sato and Nieminen 2004), we could not detect any signals for galectin-3 in lamina propria cells including gastric mast cells and intestinal macrophages or in hepatic sinusoidal macrophages. Our failure to detect galectin-3 mRNA in leukocytes under normal conditions may be explained by the fact that infection and inflammation readily induce the galectin-3 expression in leukocytes (Sato et al. 2002b; Almkvist and Karlsson 2004). Instead, the analysis in the present study of normal mice demonstrated the broadest distribution of galectin-3 transcripts in epithelia from the lip to the anus. Previous immunohistochemical studies for galectin-3 briefly documented the positive reactivity in epithelial cells of the human colon (Sanjuan et al. 1997) and of the bovine duodenum and colon (Kaltner et al. 2002). In the present observation, the expression of galectin-3 mRNA in mice was characterized by an intensified expression on the luminal side of the gastric and intestinal mucosa. A similar tendency for galectin-3 expression was noted in immunostaining of galectin-3 in the human (Sanjuan et al. 1997) and the bovine colons (Kaltner et al. 2002). Interestingly, galectin-3 suppresses apoptosis in vitro through a cell death inhibition pathway that involves Bcl-2 (Yang et al. 1996). The expression of galectin-3 has also been identified in various types of tumor cells both in vivo (Oda et al. 1991; Woo et al. 2001; Takenaka et al. 2003) and in vitro (Cebo et al. 2002), suggesting a possibility that it stimulates tumor cell proliferation by inhibiting cell death. It is well known that the major population of aged epithelial cells in the gut move to luminal sides and drop off into the lumen for epithelial renewal; therefore, galectin-3 may act to prevent epithelial cells from cell death at the terminal sites of maturation.
In adult mouse tissues, the expression of galectin-4/6 was assayed by Northern and Western blots only in gastrointestinal tissues (Gitt et al. 1998). An RNase protection assay, which permitted their separate detection, demonstrated a high expression of galectin-4 in the small intestine and colon but a lower level of expression in the stomach, whereas galectin-6 exhibited a relatively even distribution from the stomach to the colon (Gitt et al. 1998). Because of the high sequence homology between galectin-4 and galectin-6, it is hard to distinguish their expression by the use of available histochemical techniques. A previous in situ hybridization study reported the combined expression of galectin-4/6 in the intestinal epithelium of mice (Gitt et al. 1998); however, the precise cellular localization has not been described. We demonstrated here the intense and consistent expression of galectin-4/6 in the epithelium of the whole small intestine from crypts to villous tips. The transcripts of galectin-4/6 in enterocytes were maintained at equal or more intense levels in the large intestine, thus resulting in the finding that galectin-4/6 is the most dominant subtype in the large intestine. Recently, galectin-4 was immunohistochemically localized in the human colon epithelium and functionally related to the promotion of colon cancer growth and exacerbation of colitis (Hokama et al. 2004; Huflejt and Leffler 2004). Although stratified epithelial cells of the oral mucosa and esophagus contain immunoreactivity for galectin-4 in the pig (Chiu et al. 1992) and the rat (Wasano and Hirakawa 1995), we detected a faint expression of galectin-4/6 mRNA only in the forestomach among the stratified epithelia. Confining its description to the simple epithelium, galectin-4/6 is regarded as a subtype that is most intensely and consistently expressed from the glandular stomach to rectum, suggesting a fundamental function shared by gastric and intestinal epithelial cells. In accordance with this suggestion, it is reported that galectin-4 resides on the extracellular side of membrane in enterocytes and has roles in the stabilization of adhesion devices (Chiu et al. 1992, 1994; Huflejt et al. 1997) and membrane-associated molecules (Danielsen and van Deurs 1997; Braccia et al. 2003).
Galectin-7 displayed a limited expression in the stratified squamous epithelium of the lip, tongue, esophagus, forestomach, and of the anus. This finding was consistent with data from Western blot (Sato et al. 2002a) and immunohistochemical studies (Magnaldo et al. 1998). However, there is a slight difference in the detailed cellular expression within the stratified epithelium. Intense labelings with a galectin-7 riboprobe and antibody were spread all over the thickness of the nucleated epithelium in the human epidermis (Magnaldo et al. 1995, 1998), in contrast to the present study showing that signals for galectin-7 mRNA were restricted to the basal half of the stratified epithelium. Nevertheless, the specific expression of galectin-7 by keratinocytes is beyond doubt and further supported by the finding that the vaginal epithelium expresses galectin-7 mRNA, whereas the respiratory pseudostratified epithelium and urinary transitional epithelium do not (Nio J, et al., unpublished data).
Galectin-9 is distributed ubiquitously and is more abundantly expressed in the small intestine, liver, and thymus at a nucleotide level (Wada and Kanwar 1997). One of the unique features of galectin-9 is the presence of its alternate splicing isoform exclusively expressed in the small intestine, suggesting certain functions of the intestinal isoform that are related to the biology of intestinal epithelium but also shared by other galectin subtypes (Wada and Kanwar 1997). When we hybridized sections of visceral organs using probes detecting both isoforms, significant signals were detected from the glandular stomach to the rectum as well as the liver (Nio J, et al., unpublished data). Galectin-9 in the gut was roughly identical to galectin-4/6 in distribution, although the intensity was much weaker than that of galectin-4/6 and remarkably so in the large intestine. Because of the existence of the isoforms, a further detailed investigation of galectin-9 is underway.
As summarized in Figure 7A, the digestive tract of mice expressed five subtypes of galectin mRNAs in the epithelium with region-dependent and cell-specific distributions. Their restricted expression in the epithelium suggests the involvement in the following: (a) digestion/absorption of food, (b) interaction with resident or pathogenic microorganisms, and (c) homeostasis of the epithelium including the regulation of cell kinetics. Although no studies have dealt with the direct relation of galectin in digestion and absorption, its interaction with digestive enzymes and mucin is proof enough to consider an indirect role in digestion. Danielsen and van Deurs (1997) have discussed how the porcine galectin-4 present in the intestinal brush border may keep digestive enzymes at the cell surface by preventing them from release into the gut lumen and also from undergoing endocytosis. One of the major ligands for galectin is mucin (Raz and Lotan 1987; Wasano and Hirakawa 1997), and especially galectin-3 modulates the expression of MUC2 mucin in human colon cancer cells (Bresalier et al. 1996). The intense expression of galectin in the mucus-secreting goblet cells and gastric pits cells as shown in the present study suggests a role in the production and secretion of mucin and epithelial cell surface glycocalyces. As an example of the second function, the production of galectin (galectin-3 and possibly galectin-4) is upregulated in response to microbial infection and secreted to interact with microorganisms (Huflejt and Leffler 2004; Sato and Nieminen 2004). It has also been proposed that galectin-3, which is present in macrophages or the mucosal epithelium, can bind to β-galactoside residues of lipopolysaccharide on pathogens to mediate the interaction between a pathogen and host cells (Mandrell et al. 1994). To date, most studies have focused on the third function, namely, the regulation of cell growth, differentiation, apoptosis, adhesion, and malignant transformation. Taking into consideration the regulation of epithelial homeostasis, it is worth noting the changes in galectin subtypes expressed by the epithelium according to the maturation process. Namely, galectin-2 expressed at the proliferating zone of crypts changes, via galectin-4/6, to galectin-3 expressed in fully matured epithelial cells at villous tips (Figure 7B). The maturation-dependent expression of galectin subtypes was also found in the mucus-secreting cells of the stomach and in the epithelial cells of the large intestine. This phenomenon provides an example of a periodical shift of galectin subtype expressed by the same cell lineage (proto type → tandem repeat type → chimera type). Likewise, the maturation-dependent expression is important when taking into consideration interaction and switching among subtypes. Galectin-1, -3, or -1/3 null mutant mice appear healthy (Poirier and Robertson 1993; Colnot et al. 1998; Pugliese et al. 2001), suggesting that their functions could be compensated for by other galectins with a similar carbohydrate specificity.
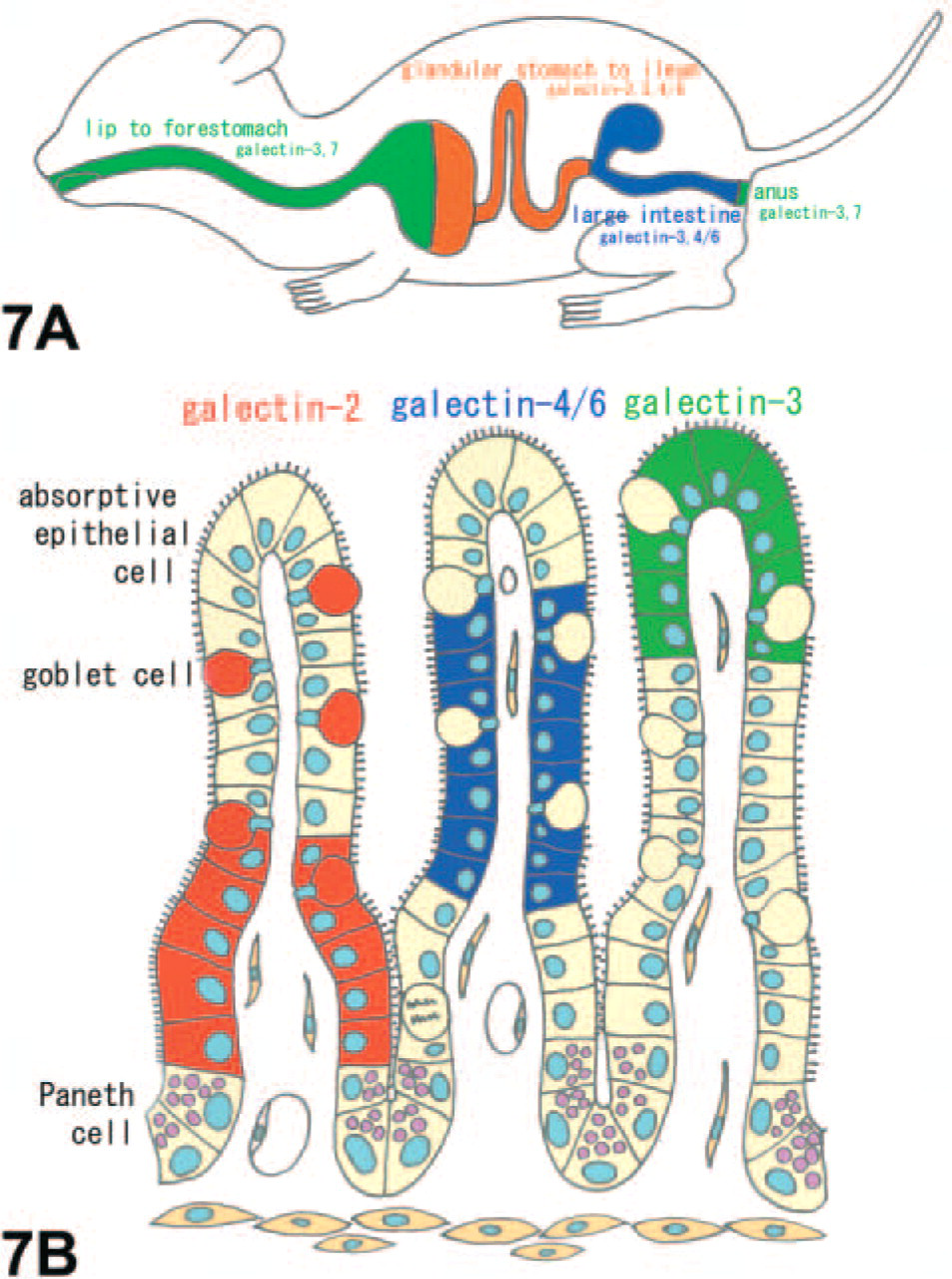
Schemes summarizing the distribution of galectin subtypes in the mouse digestive tract. The digestive tract of mice expresses five subtypes of galectin mRNAs (galectin-2, -3, -4/6, and -7) in the epithelium with region dependency (
Galectins are an evolutionally conserved family of animal lectins, and their functions are manifold. The region-dependent and cell-specific localization of galectin subtypes as well as their intense expression throughout the digestive tract strongly suggest their contribution to epithelial homeostasis, host-pathogen interactions, or mucosal immunity in the digestive tract.
Footnotes
Acknowledgements
This work was supported by a grant from the Ministry of Education, Science, Sport, and Culture, Japan (number 15390052 to TI).