
Abstract
Galectin, an animal lectin that recognizes β-galactoside of glycoconjugates, is abundant in the gut. This IHC study showed the subtype-specific localization of galectin in the mouse digestive tract. Mucosal epithelium showed region/cell-specific localization of each galectin subtype. Gastric mucous cells exhibited intense immunoreactions for galectin-2 and galectin-4/6 with a limited localization of galectin-3 at the surface of the gastric mucosa. Electron microscopically, galectin-3 immunoreactivity coated indigenous bacteria on the gastric surface mucous cells. Epithelial cells in the small intestine showed characteristic localizations of galectin-2 and galectin-4/6 in the cytoplasm of goblet cells and the baso-lateral membrane of enterocytes in association with maturation, respectively. Galectin-3 expressed only at the villus tips was concentrated at the myosin-rich terminal web of fully matured enterocytes. Epithelial cells of the large intestine contained intense immunoreactions for galectin-3 and galectin-4/6 but not for galectin-2. The stratified squamous epithelium of the forestomach was immunoreactive for galectin-3 and galectin-7, but the basal layer lacked galectin-3 immunoreactivity. Outside the epithelium, only galectin-1 was localized in the connective tissue, smooth muscles, and neuronal cell bodies. The subtype-specific localization of galectin suggests its important roles in host-pathogen interaction and epithelial homeostasis such as membrane polarization and trafficking in the gut.
G
In the mouse, nine subtypes of galectin (galectin-1, −2, −3, −4, −6, −7, −8, −9, and −12) have been reported to be expressed in a tissue/cell-specific manner. The digestive tract is one of the organs rich in galectin; we previously showed at the mRNA level that at least six subtypes of galectin (galectin-2, −3, −4, −6, −7, and −9) were intensely and continuously expressed from the mouth to the anus. All of these subtypes were localized in the epithelium and exhibited characteristic distribution patterns (Nio et al. 2005). Interestingly, when some galectin subtypes were coexpressed in the glandular stomach and the small intestine, the predominant subtypes changed during epithelial differentiation (from galectin-2, through galectin-4/6, to galectin-3). Because galectin is involved in cell adhesion and growth in addition to mediating immunological events (Rabinovich et al. 2007b), the region-specific expression of galectin in the digestive tract is suggestive of its important roles in the epithelial homeostasis and gut diseases such as inflammatory bowel diseases and gastric cancer. A recent review by Demetter et al. (2008) described the different roles of galectin subtypes in the pathogenesis of gastrointestinal diseases.
Compared with the involvement of galectin in various pathological events, little is known about the cellular and subcellular localizations of galectin in the digestive tract under normal conditions. Moreover, the morphological data are inconsistent among researchers because of a lack of specific antibodies. In this study, we produced subtype-specific antibodies and showed the cellular localization of six major subtypes of galectin (galectin-1, −2, −3, −4, −6, and −7) in the digestive tract of normal mice.
Materials and Methods
All animals were treated according to the laboratory animal control guidelines of Hokkaido University (Approval 08–054), which conform to Guide for Care and Use of Laboratory Animals of U.S. Institute for Laboratory Animal Research.
Subtype-specific Antibodies for Galectin
Polyclonal antibodies to mouse galectin-1, −2, and −4 were raised against the amino acid residues 112–135 of galectin-1, 1–60 of galectin-2, and 307–326 of galectin-4. The amino acid sequences among galectin subtypes show <47% sequence homology, except for galectin-4 and galectin-6 (83%). The polypeptides were expressed as glutathione S-transferase (GST) fusion proteins using the pGEX4T-2 vector (GE Healthcare Biosciences; Uppsala, Sweden) and BL21 cells (Takara; Tokyo, Japan). The fusion protein was purified with glutathione-Sepharose 4B (GE Healthcare Biosciences), emulsified with Freund's complete or incomplete adjuvant (Difco; Detroit, MI) and injected SC into female New Zealand White rabbits and Hartley guinea pigs (Japan SLC; Shizuoka, Japan) at intervals of 2 weeks. Ten days after the fifth injection, affinity purified antibodies were prepared from serum, first using Protein G-Sepharose (GE Healthcare Biosciences) and then using antigen peptides coupled to CNBr-activated Sepharose 4B (GE Healthcare Biosciences). For the preparation of affinity media, antigen peptides free from GST were obtained by the elution of cleaved polypeptides after the in-column digestion by thrombin (Sigma; St. Louis, MO).
Antibodies for galectin-3 and galectin-7 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA) and Bethyl Laboratories (Montgomery, TX), respectively.
Western Blotting
Adult male ddY mice at 8 weeks of age (Japan SLC) were used for Western blotting analysis. Mice were killed by bloodletting from the heart under deep anesthesia with pentobarbital, and fresh tissues including the forestomach and small intestine were removed. Tissues were washed sufficiently with physiological saline and homogenized with ice-cold 10 mM Tris-HCl buffer (pH 7.5) containing 1 mM EDTA, 0.3 M lactose, and protease inhibitor cocktail (Complete Mini; Roche, Mannheim, Germany).
Soluble fractions of all samples (20 μg/lane) were subjected to 12% (for galectin-3 and −4) or 15% (for galectin-1, −2, and −7) SDS-PAGE under reducing conditions. The proteins were transferred to polyvinylidene difluoride membranes (Hybond-P; GE Healthcare Biosciences) and incubated with rabbit antibodies against galectin-1 (0.1 μg/ml), galectin-3 (0.1 μg/ml), galectin-4 (0.1 μg/ml), and galectin-7 (0.01 μg/ml), or guinea pig anti-galectin-2 antibody (0.1 μg/ml). The bound antibodies were visualized using peroxidase-labeled anti-rabbit or guinea pig immunoglobulins (IgG; 1:100,000 in dilution; Dako, Glostrup, Denmark) and an enhanced chemiluminescence system (ECL-plus; GE Healthcare Biosciences) according to the manufacturer's instructions.
IHC
Male ddY mice at 8 weeks of age (Japan SLC) were deeply anesthetized with pentobarbital and perfused through the aorta with physiological saline, followed by Bouin's fluid. The whole stomach, middle region of the small intestine, and the proximal colon were removed and immersed in the same fixative for an additional 12 hr at 4C. The Bouin-fixed tissues were dehydrated through a graded series of ethanol and embedded in paraffin according to a conventional method. Sections were cut at 4-μm thickness and mounted on gelatin-coated glass slides. They were deparaffinized and rinsed in PBS (pH 7.4). After preincubation with 10% normal goat serum for 30 min, the sections were incubated with rabbit antibodies against galectin-1 (1.0 μg/ml), galectin-3 (1.0 μg/ml), galectin-4 (1.0 μg/ml), galectin-7 (0.1 μg/ml), and α-smooth muscle actin (α-actin; 1:500 in dilution; Epitomics, Burlingame, CA), or guinea pig anti-galectin-2 antibody (1.0 μg/ml) overnight at room temperature. Sections were incubated with biotinylated goat antibody against rabbit IgG followed with peroxidase-labeled streptavidin by using Histofine Kit (Nichirei; Tokyo, Japan) (for galectin-1, −3, −4, −7, and α-actin) or peroxidase-labeled anti-guinea pig IgG (1:200 in dilution; Dako) (for galectin-2). The signals were visualized with 0.01% 3,3′-diaminoben-zidine in 0.05 M Tris-HCl buffer (pH 7.6) containing 0.003% H2O2 and counterstained with hematoxylin.
Another group of mice was perfusion-fixed with 4% paraformaldehyde for IHC for myosin. The small intestine was dissected and immersed in the same fixative for an additional 12 hr at 4C. Fixed tissues were immersed in 30% sucrose overnight, embedded in O.C.T. compound (Sakura Finetechnical Co.; Tokyo, Japan), and frozen with liquid nitrogen. Frozen sections, at 10-μm thickness were stained with rabbit anti-myosin antibody (dilution 1:1000; Biomedical Technologies, Stoughton, MA). The binding sites were visualized with biotinylated goat antibody against rabbit IgG followed by peroxidase-labeled streptavidin.
For IHC at the electron microscopic level, the paraformaldehyde-fixed stomach was used and processed as described in a previous study (Nio et al. 2006).
Double Fluorescent Staining
For double immunofluorescence of galectins and proliferating cell nuclear antigen (PCNA) or chromogranin A (CgA), Bouin-fixed paraffin sections were incubated with rabbit anti-CgA (Hashimoto et al. 2001) or anti-PCNA (1:200 in dilution; Santa Cruz Biotechnology, Santa Cruz, CA), and galectin-3 antibody overnight, followed by Cy3-conjugated anti-rabbit IgG (Jackson Immunoresearch Laboratories; West Grove, PA) for 2 hr. After rinsing in PBS, the sections were incubated overnight at room temperature with either anti-galectin-2 or anti-galectin-4 antibodies, the latter being labeled with Alexa Fluor 488 by using Zenon Rabbit IgG Labeling Kits (Molecular Probes; Eugene, OR). The binding sites of the galectin-2 antibody were visualized with FITC-conjugated donkey anti-guinea pig IgG (Jackson Immunoresearch Laboratories).
Double staining of galectin-2 and galactose-containing carbohydrate recognizing plant lectin, Erythrina cristagalli lectin (ECL), was performed on the same Bouin-fixed paraffin section. After the sections were immunostained with the galectin-2 antibody and Cy3-conjugated anti-guinea pig IgG, they were incubated with biotinylated-ECL (1:1000 in dilution; Vector Laboratories, Burlingame, CA) overnight at room temperature. The lectin-binding sites were visualized with FITC-streptavidin (1:100 in dilution; Zymed Laboratories, South San Francisco, CA) for 1 hr at room temperature. These sections were observed under a confocal laser scanning microscope (FV300; Olympus, Tokyo, Japan).
Results
Antibody Specificity
The antibodies used in this study were characterized by Western blot analysis using the extracts from the small intestine (galectin-1, −2, −3, and −4) and the forestomach (galectin-7). Each antibody detected a predominant immunoreactive band at the estimated molecular size (Figures 1A–1E). The antibody raised against the carboxyl terminal of galectin-4 exhibited two immunoreactive bands around 36 kDa. It is likely that the antibody recognized both galectin-4 and galectin-6 because of their high sequence homology (83% in whole amino acid sequences and 13/20 in amino acids of the antigen regions; Figure 1F)—described below as galectin-4/6. A minor immunoreactive band with the galectin-4 antibody appeared at a higher molecular level than the estimated size (Figure 1D). This may correspond to the dimer, because galectin-4 easily aggregates during protein extraction even if using a buffer containing lactose, a β-galactoside-specific sugar. Furthermore, the cross-reactivity among subtypes was excluded by antigen absorption tests in Western blotting (data not shown).
IHC
We previously showed at a mRNA level that the digestive tract of mice expressed at least six subtypes of galectin (galectin-2, −3, −4/6, −7, and −9) in the epithelium with subtype-specific patterns (Nio et al. 2005). This IHC study examined the localization of five epithelial type of galectin (galectin-2, −3, −4/6, and −7) and a stromal type of galectin (galectin-1) at a protein level throughout the mouse digestive tract. The specificities of the immunoreactions on sections were confirmed by a conventional protocol including absorption tests by use of antigens and by the fact that the staining results perfectly coincided with the mRNA distributions previously shown by ISH (Nio et al. 2005).
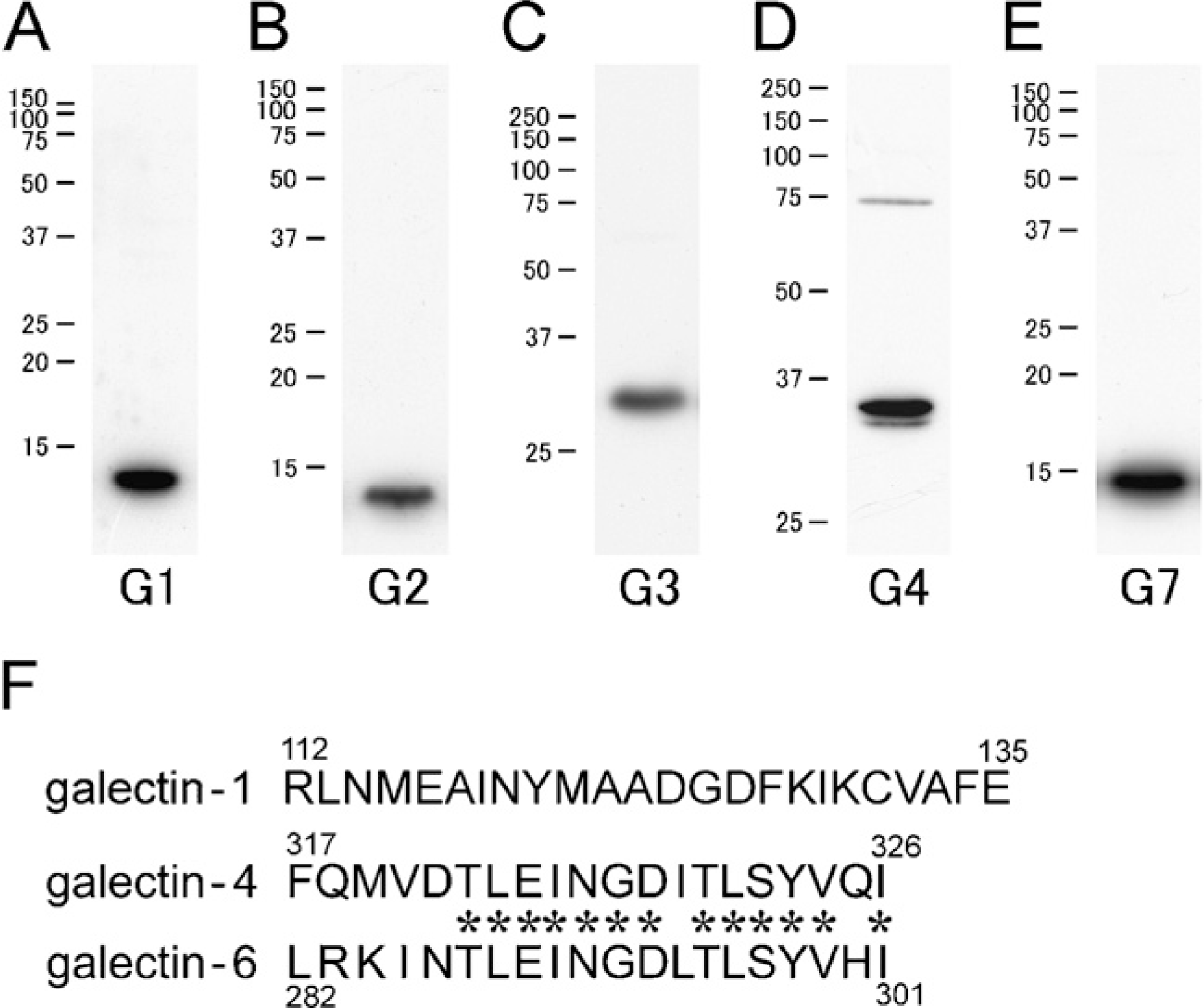
Western blot analysis with subtype-specific antibodies for galectin. Predominant immunoreactive bands are found around an estimated molecular mass: 14 kDa for galectin-1 (G1)
In the glandular stomach, intense immunoreactivities for galectin-2 and galectin-4/6 were found in the upper region of the mucosal layer, the former being more deeply distributed in the gastric glands (Figures 2A and 2B). Under higher magnification, cells immunoreactive for both galectins were identified as surface mucous cells and mucous neck cells (Figures 2C and 2D), but parietal cells and chief cells were free from the immunoreactions. The immunoreactivity for galectin-2 existed at more differentiated mucous neck cells than that for galectin-4/6, which was restricted to the proliferating zone (the isthmus). The galectin-2 immunoreactivity was diffusely localized in the cytoplasm, whereas galectin-4/6 showed an intensified immunoreactivity at the baso-lateral membrane of surface mucous cells (Figures 2C and 2D). In addition, a weak immunoreactivity for galectin-3 was found in the mucous cells only at the surface of the mucosa, where the positive reactions frequently appeared to be granular in the cytoplasm (Figure 2E). Interestingly, the immunostaining for galectin-3-labeled indigenous bacteria attached to the surface of the gastric mucosa (Figure 2F). Electron microscopically, gold particles showing the existence of galectin-3 aggregated at the surface of the microorganisms (Figure 2G). In the forestomach, unkeratinized cells of the stratified squamous epithelium were intensely immunoreactive for galectin-7 and moderately immunoreactive for galectin-3, but the basal layer lacked galectin-3 immunoreactivity (Figures 2H and 2I).
Epithelial cells in the small intestine showed a maturation-associated distribution of galectin-2, galectin-4/6, and galectin-3 (Figures 3A–3C), which essentially corresponded to the mRNA distribution shown by ISH (Nio et al. 2005). Immunoreactivity for galectin-2 gathered in the upper half of crypts and the base of villi, where the cytoplasm of epithelial cells was diffusely immunolabeled (Figure 3A); galectin-2 immunoreactivity in the enterocytes of villi sharply decreased upward. In contrast, the immunoreaction of galectin-2 appeared in goblet cells and increased in intensity toward the villus tips (Figure 3A). Lectin histochemistry using ECL, which recognizes the galactose-containing carbohydrate, selectively detected mucous granules of goblet cells. In double staining with galectin-2 antibody and ECL, galectin-2 was localized in the cytoplasm of goblet cells, but not in ECL-positive mucous substances (Figure 3D). An intense immunoreactivity for galectin-4/6 was distributed from the upper half of crypts to the lower three quarters of the villi but disappeared at the villus tips (Figure 3B). Immature enterocytes at lower regions of the villi were diffusely immunolabeled with the galectin-4/6 antibody, including the brush border and the baso-lateral membrane. Mature enterocytes at the upper region of the villi showed the predominant localization of galectin-4/6 in the cell membrane, especially on the baso-lateral side (Figure 3E). Galectin-3 immunoreactivity was present only at the villus tips (Figure 3C). The galectin-3 immunoreactivity exhibited a concentrated localization in an apical cytoplasm (Figures 3F and 3G). Compared with the immunolocalization of α-actin and myosin, galectin-3 was localized at the myosin-rich terminal web but not in the α-actin-rich brush border (Figures 3G–3I).
The crypt epithelium contained immunoreactivities for galectin-2 and galectin-4/6 in the cytoplasm. The upper two thirds of the crypts, corresponding to the proliferative zone, were immunoreactive for these galectins, but Paneath cells occupying the bottom of crypts lacked any immunoreactivities. The galectin-positive region in the crypts contained proliferating cells with proliferating cell nuclear antigen (PCNA) immunoreactivity in nucleus (Figure 3J) (only the data for galectin-2 is shown). When double staining with CgA, a marker protein of endocrine cells (Hashimoto et al. 2001), was performed, CgA-positive endocrine cells were negative in immunoreactions for both galectin-2 and galectin-4/6 (Figures 3K and 3L).
In the large intestine, intense immunoreactions for galectin-3 and galectin-4/6 were found in the covering epithelium and crypts, whereas galectin-2 immunoreactivity was totally absent in epithelial cells, including goblet cells. Galectin-3 immunoreaction was restricted to the upper half of crypts (Figure 4A), whereas galectin-4/6 was distributed as deep as the bottoms of crypts (Figure 4B). Both immunoreactivities, especially for galectin-4/6, tended to gather at the baso-lateral membrane (Figures 4A and 4B).
Immunostaining of galectin-1, a stromal type of galectin, intensely labeled the connective tissue and muscle layer but not the epithelium throughout the gastrointestinal tract. The immunoreactivities in the connective tissue were localized in fibroblasts of lamina propria and tela submucosa (Figures 5A and 5B). The smooth muscle cells of lamina muscularis mucosae and muscular layer were intensely immunoreactive for galectin-1 (Figures 5C and 5D), whereas tunica media of blood vessels were weakly immunolabeled (Figure 5C). In the small intestine, some populations of leukocytes at lamina propria and tela submucosa and a small number of intraepithelial lymphocytes were labeled with the galectin-1 antibody (Figure 5B). Neuronal cell bodies in submucous and myenteric nerve plexuses were also immunoreactive for galectin-1 (Figure 5D). Galectin-1 immunoreactivity was diffusely distributed in the cytoplasm of all these cells (Figures 5B–5D).
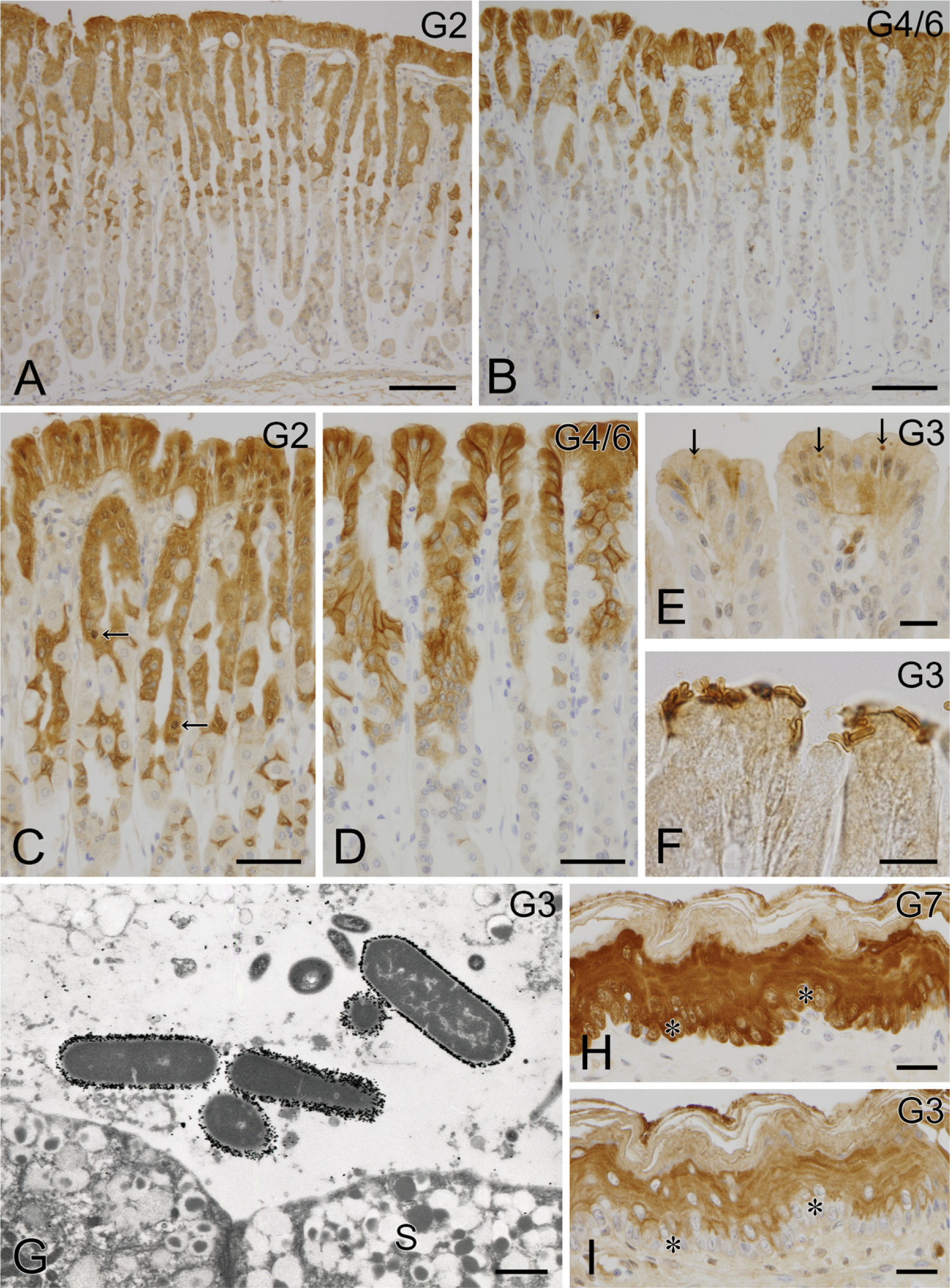
IHC localization of galectin in the stomach. Intense immunoreactivities for galectin-2
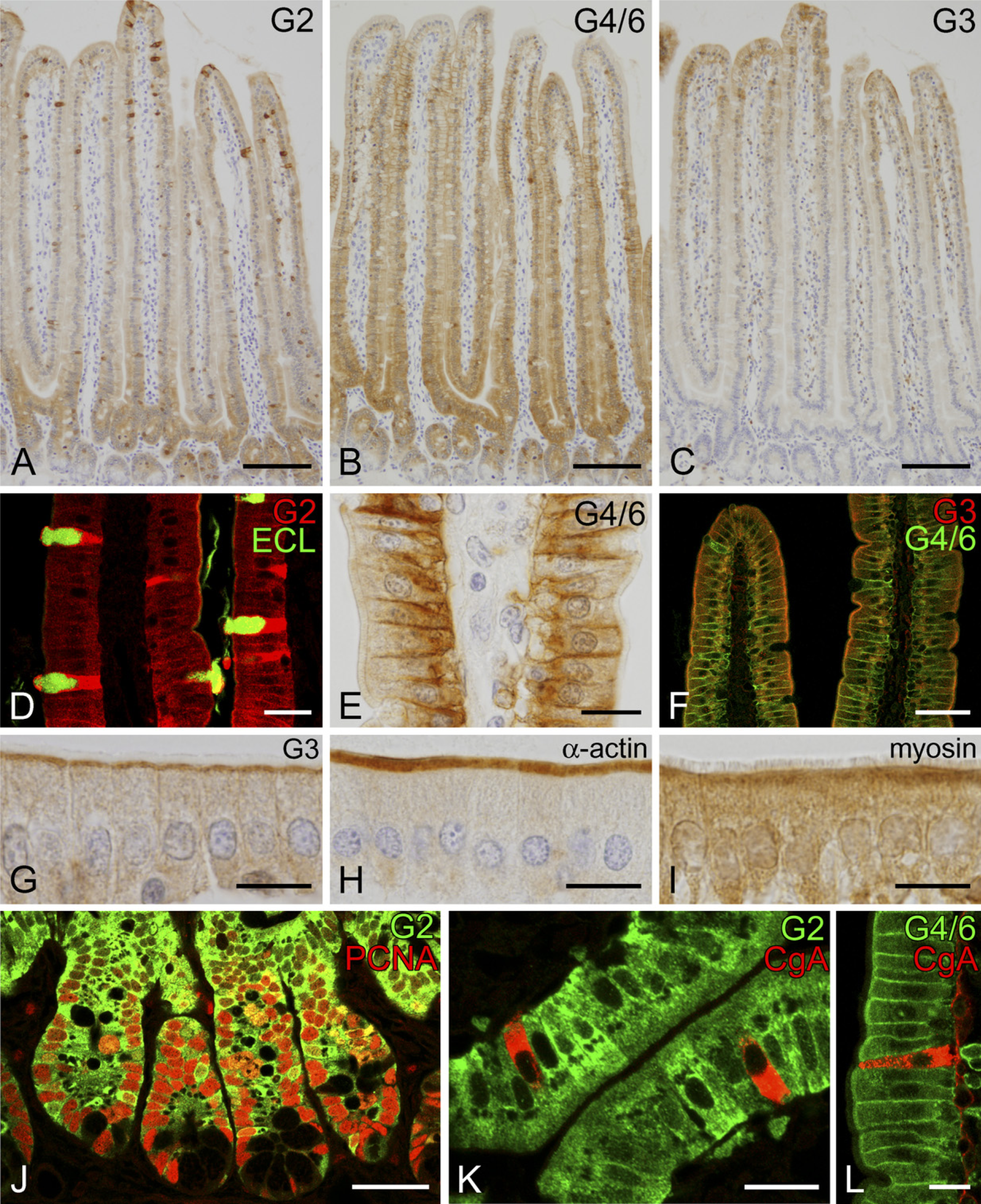
IHC localization of galectin in the small intestine. Diffuse immunoreactivity for galectin-2 is seen in the upper region of crypts and the base of villi
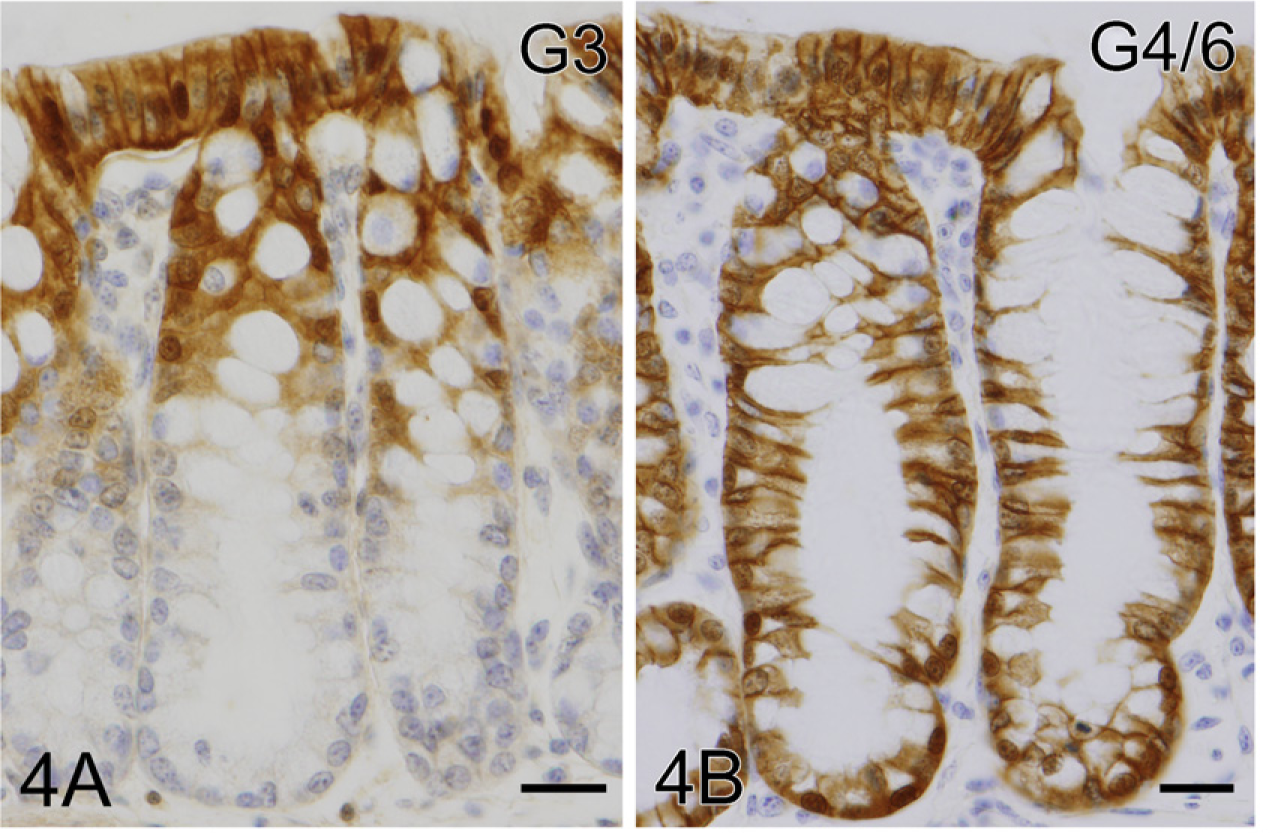
IHC localization of galectin in the proximal colon. The covering epithelium and upper half of crypts contain intense immunoreactions for galectin-3
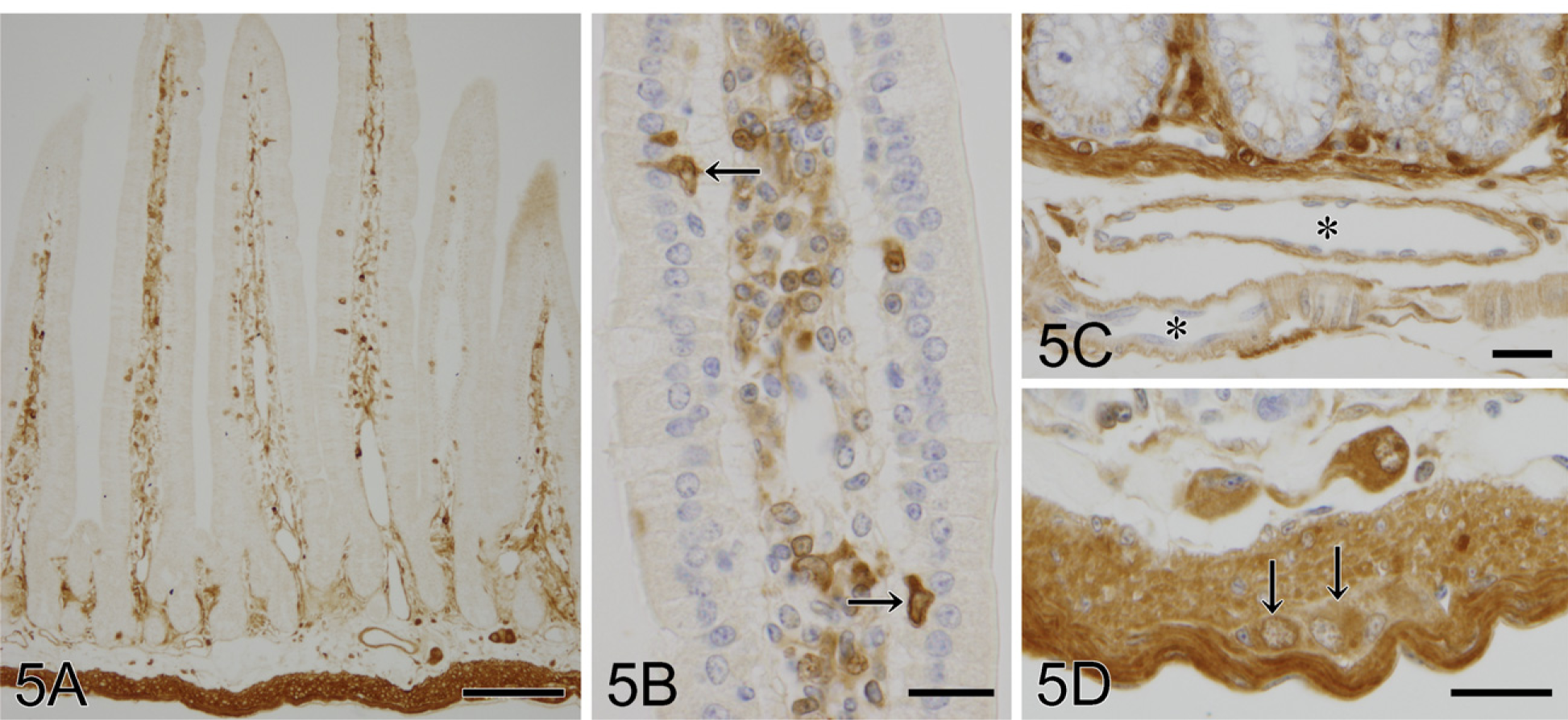
Galectin-1-immunoreactive cells in the small intestine
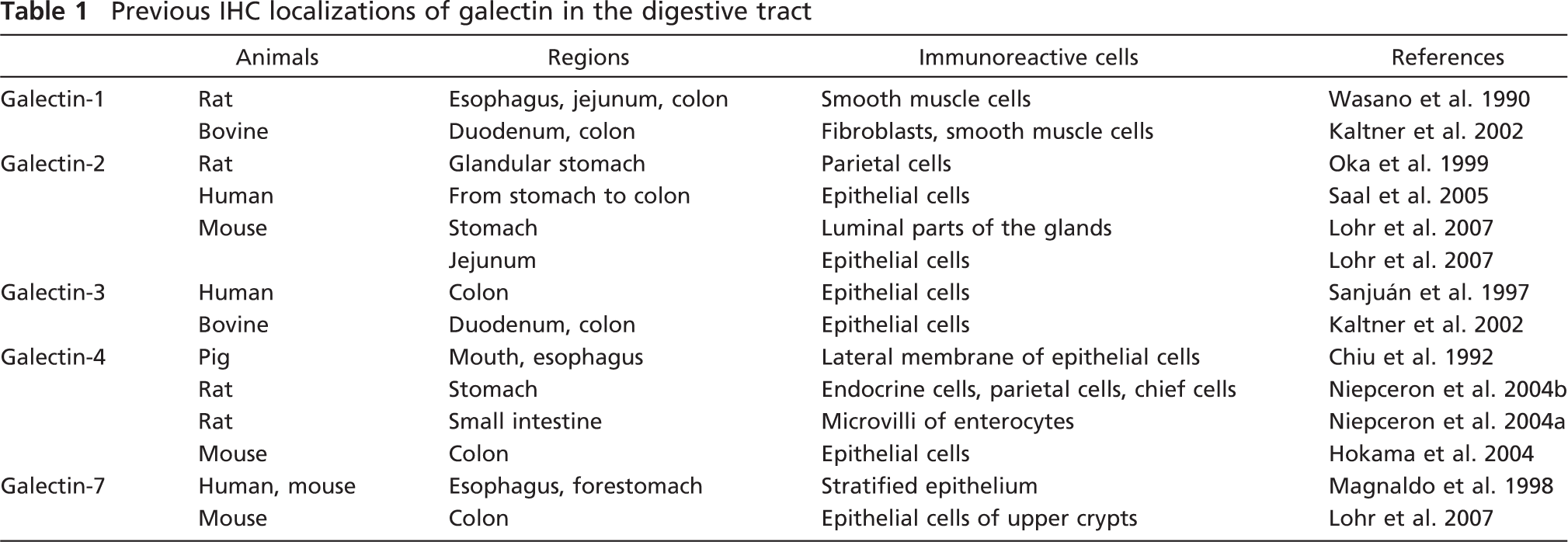
Previous IHC localizations of galectin in the digestive tract
Discussion
Galectin is defined by having a conserved carbohydrate-recognition domain with affinity for β-galactosides of glycoconjugates (Barondes et al. 1994). To date, 15 mammalian galectins have been identified that can be subdivided into three groups by their chemical structures (Hirabayashi and Kasai 1993): proto type (galectin-1, −2, −5, −7, −10, −11, −13, −14, −15), tandem-repeat type (galectin-4, −6, −8, −9, −12), and chimeric type (galectin-3). This IHC study clearly showed the broad and characteristic localizations of major six subtypes of galectin (galectin-1, −2, −3, −4/6, and −7) in the mouse digestive tract. Previous studies have reported the IHC localization of galectin in the digestive tract of mammals as summarized in Table 1. Some of the galectin-immunoreactive cells shown by this study are coincident to cell types documented in the previous studies, but others are controversial. The discrepancy may be partially caused by differences in antibodies and fixatives used. In a preliminary study, we could obtain the best immunostaining results without any diffusion of antigens and background noise if tissues were treated with 5–10% acid-containing fixatives. This study used such materials fixed with Bouin's fluid, which contained ∼5% acetic acid, and the obtained immunostaining results perfectly coincided with the expression pattern of galectin mRNA shown in our previous study (Nio et al. 2005). Thus, this systematic study for galectin could show intense and subtype-specific distributions of galectin-2, −3, −4/6, and −7 in the mucosal epithelium of the gut (Table 2). The maturation-associated expression of galectin was confirmed at a protein level: the predominant subtype in the glandular stomach and the small intestine changed according to the epithelial differentiation. Furthermore, this staining found that, in association with maturation, the subcellular localization of galectin-2 and galectin-4/6 focused to the cytoplasm of goblet cells and baso-lateral membrane of enterocytes, respectively. Fully matured enterocytes at the villus tips exhibited a unique localization of galectin-3 in the terminal web. These findings showed that galectin is diffusely distributed in the cytoplasm of undifferentiated cells, whereas it gathers at special sites in differentiated cells, suggesting the acquisition of the subtype-specific function.
Because it is generally accepted that proteins present in the cytoplasmic compartment of eukaryotic cells are not glycosylated, galectin should function extracellularly. However, many studies have documented the cytoplasmic and/or nuclear localizations of galectin. This study also confirmed the abundant localization of galectin in the cytoplasm of epithelia throughout the gastrointestinal tract, implying the existence of intracellular ligands for galectin. A recent study has identified several intracellular ligands for galectin: oncogenic H-Ras as ligands for galectin-1, and K-Ras, Bcl-2, and β-catenin as ligands for galectin-3 (Paz et al. 2001; Elad-Sfadia et al. 2004; Shimura et al. 2004). Although it is unclear whether the interaction between galectin and these ligands depends on carbohydrate recognition or a protein-protein interaction, the cellular localization of galectin suggests its intracellular function in the gut epithelia. For example, the selective localization of galectin-2 in the cytoplasm of goblet cells reminds us of a special role of galectin-2 in mucus-producing cells, and a further study on the function of galectin-2 in mucus-secreting cells is underway in our laboratory. Galectin-3 expressed only at the villus tips may affect the shedding off and apoptosis of fully matured epithelial cells. In several in vitro studies, galectin-3 suppressed apoptosis by binding to various ligands such as Bcl-2 and Ras proteins (Nangia-Makker et al. 2007).
Galectin-expressing cells in the mouse gastrointestinal epithelium

This study showed the diffuse localization of galectin-4/6 in the cytoplasm of immature enterocytes and a shift to the baso-lateral membrane in matured enterocytes, suggesting its involvement in membrane trafficking or stabilization. Danielsen and van Deurs (1997) showed that galectin-4 is localized in the brush border membrane isolated from porcine enterocytes and may regulate the formation and stabilization of lipid rafts that are engaged in the intracellular transport of glycosphingolipids and membrane proteins to the apical plasma membrane. This idea is supported by another in vitro study by Delacour et al. (2005), showing that galectin-4 could cross-link glycolipids to form raft platforms for the apical sorting. However, this IHC study showed the predominant localization of galectin-4/6 in the baso-lateral membrane of enterocytes. The baso-lateral localization of galectin-4 was also documented by Chiu et al. (1992) and Lindstedt et al. (1993) in the porcine oral epithelium and a human colon-derived cell line (T84 cells). Therefore, our findings on the subcellular localization of galectin-4/6 suggest that intestinal galectin-4/6 may function in baso-laterally oriented trafficking rather than in the transportation to the apical cell surface.
In contrast, this immunostaining for galectin-3 showed its concentrated localization at the myosin-rich terminal web of fully matured epithelial cells at the villus tips. The apical localization of galectin-3 was previously observed in T84 cells and other polarizing epithelial cell lines (Lindstedt et al. 1993; Huflejt et al. 1997). Galectin-3 is required for intracellular sorting and correct targeting of glycoproteins to the apical plasma membrane (Delacour et al. 2006). In accordance with this, an analysis of galectin-3 knockout mice showed that actin and villin abundant in the brush border become abnormally distributed along baso-lateral membranes (Delacour et al. 2008). Interaction of galectins, including galectin-3, with actin (Chiu et al. 1992; Joubert et al. 1992), cytokeratins (Goletz et al. 1997), and possibly tubulin (Ozaki et al. 2004) has been indicated. Although it remains unknown whether or not galectin-3 binds to myosin, galectin-3 at the terminal web may function to stabilize the brush border membrane or to support the apical translocation of brush border enzymes by interaction with cytoskeletal proteins.
Secretion and the extracellular localization of galectin have been noted by in vivo and in vitro studies, at least for galectin-1 and galectin-3 (Cooper 1997; Ochieng et al. 2004). It is generally believed that secretory galectin is transported through an atypical secretory route because they lack a signal sequence required for export by vesicle-mediated exocytosis (Hughes 1999). In this study, only galectin-3 was detected extracellularly, whereas the other galectin subtypes were found in neither extracellular matrix nor secretory granules. Interestingly, the galectin-3 immunoreactivity concentrated around indigenous bacteria on the gastric surface mucous cells, as observed at the electron microscopic level. This finding supports previous in vitro studies showing that galectin-3 is secreted and can interact with bacterial surface carbohydrates such as lipopolysaccharide (Rabinovich et al. 2002). Because the secretory pathway of galectin has not been established, a more detailed study at electron microscopic levels is needed.
In conclusion, this study clearly showed the subtype-specific localization of galectins in the mouse digestive tract. Particularly, it is worth noting that the cellular and subcellular localizations of each galectin subtype were specialized in mature epithelial cells. Although the ligands of galectin remain unknown, these histochemical data contribute to a better understanding of the functions and pathological involvements of galectin in the digestive tract.
Footnotes
Acknowledgements
This work was supported by a Grant-in-Aid for Young Scientists (B) (19790146) from the Ministry of Education, Culture, Sports, Science, and Technology, Japan.
The authors thank Prof. Masahiko Watanabe and Dr. Hidemi Shimizu, Hokkaido University Graduate School of Medicine, for kind advice to produce subtype-specific antibodies for galectin. The authors are also grateful to Prof. Ken-Ichi Kasai, Teikyo University, for his encouragement.