
Abstract
CYP2S1 is a recently discovered dioxin-inducible member of the cytochrome P450 superfamily. It has been shown to be involved in the metabolism of some aromatic hydrocarbons as well as retinoic acid, suggesting a role in biotransformation of both exogenous and endogenous compounds. In this study, we used mRNA in situ hybridization and immunohistochemistry to investigate the cellular localization of CYP2S1 in various human tissues using tissue microarrays. High expression levels were observed mainly in epithelial cell types, especially in the epithelia frequently exposed to xenobiotics. In the respiratory tract, the expression was strong in nasal cavity, bronchi, and bronchioli, whereas it was low in the alveolar lining cells. Similarly, CYP2S1 was highly expressed in the epithelial cells throughout the gastrointestinal tract. Strong epithelial expression was also observed in uterine cervix, urinary bladder, and skin. In many exocrine glands (e.g., adrenal gland and pancreas), secretory epithelial cells showed moderate to strong expression levels. In the liver, the expression was low. CYP2S1 was highly expressed in epithelial cells that are major targets for carcinogen exposure and common progenitor cells to tumor development. Indeed, we found strong CYP2S1 expression in many tumors of epithelial origin.
Keywords
C
Most of the CYP genes have the highest expression in the liver, which has a dominant role in the clearance of foreign compounds. Many CYPs, however, are also expressed in the extrahepatic tissues (Hukkanen et al. 2002; Ding and Kaminsky 2003). The reactive intermediates resulting from CYP-catalyzed reactions are often unstable, and on-site metabolism is therefore considered of major importance for the susceptibility of tissues to toxic and carcinogenic compounds as well as for effectiveness of pharmacological treatments (Pelkonen and Raunio 1997; Ding and Kaminsky 2003).
CYP2S1 is a recently discovered member of the CYP superfamily (Rylander et al. 2001; Rivera et al. 2002). The human CYP2S1 gene, a sole member of the CYP2S subfamily, has recently been shown to be polymorphic, with two amino acid-changing allelic variants detected in Caucasians (Saarikoski et al. 2004). It is located in a cluster of CYP2 family members on chromosome 19q. In addition to CYP2S1, this cluster includes the CYP2A6, CYP2A13, CYP2B6, and CYP2F1 genes (Hoffman et al. 2001). The proteins encoded by these genes are the closest relatives to human CYP2S1, with 47–49% sequence identity.
Our studies have indicated that even though it belongs to the CYP2 family, CYP2S1 exhibits many features typical of members of the CYP1 family, including dioxin-inducibility mediated by Aryl hydrocarbon receptor (AHR) and Aryl hydrocarbon receptor nuclear translocator (ARNT) (Rivera et al. 2002). Inducibility by dioxin suggests that CYP2S1, similar to dioxin-inducible CYPs of the CYP1 family, may participate in the metabolism of toxic and carcinogenic compounds (Rivera et al. 2002; Shimada and Fujii-Kuriyama 2004). This proposal is supported by a recent report noting that small aromatic hydrocarbons, indeed, constitute substrates for CYP2S1 (Karlgren et al. 2005). Furthermore, CYP2S1 was recently shown to be induced in psoriasis patients after topical treatment with crude coal tar, an abundant source of polycyclic aromatic hydrocarbons (PAHs) (Smith et al. 2003). The same study reported that CYP2S1 metabolizes all-trans retinoic acid, indicating that, in addition to xenobiotics, CYP2S1 may be important in metabolism of endogenous substrates important for cell proliferation and differentiation.
CYP2S1 has been found to have low expression levels in the liver but to be highly and constitutively expressed in many extrahepatic tissues (Rylander et al. 2001; Rivera et al. 2002; Choudhary et al. 2003; Smith et al. 2003). The cell types that express CYP2S1 in different tissues have not yet been characterized, however. Because of the reactive and short-lived nature of many of the metabolites generated in CYP-mediated reactions, the cell type-specific metabolism has a central role for the biological effects of the substrate compounds. Therefore, in the present study, we applied in situ hybridization to mRNA and immunohistochemistry to investigate cellular localization of CYP2S1 expression in various human tissues, including tumor samples.
Materials and Methods
Tissue Microarrays
The study was performed using tissue microarrays, which included paraffin-embedded material from various types of healthy organs and tumorous tissues (slides AA7 and BC4; SuperBioChips Laboratories, Seoul, Korea). The data available for the samples included the age, gender, and diagnosis of the patient.
In Situ Hybridization
The probe used for in situ hybridization comprised of a 160-bp sequence spanning the 3’ end of the protein-coding region of CYP2S1 cDNA (GenBank accession NM 030622). The fragment was cloned to pGEM-T vector, and the [35S]-labeled single-stranded antisense RNA probe was synthesized by using the Riboprobe Combination System SP6/T7 kit (Promega, Madison, WI) according to manufacturer's instructions, as described earlier (Saarikoski et al. 1998).
For in situ hybridization, the tissue arrays were first de-paraffinized by baking for 1 hr in 60C, dewaxing in a series of xylene, and hydrated through a series of graded ethanol. The pretreatments and in situ hybridization procedure were otherwise the same as described earlier for cryosections (Saarikoski et al. 1998), except that an additional pretreatment with proteinase K (10 μg/ml, 30 min at RT) was added. The hybridization was carried out at 50C, and the posthybridization washes were carried out with 2 × SSC and 0.1 × SSC at 51C. For a negative control, some of the tissue slides were treated with RNase A (before proteinase K treatment). The slides were counterstained with eosin.
Immunohistochemistry
The rabbit polyclonal anti-human CYP2S1 antibody used was raised against a 15-amino acid CYP2S1-specific sequence located on the C terminus of CYP2S1 (Rylander et al. 2001; Smith et al. 2003).
The immunohistochemical analyses were performed as described earlier (Wikman et al. 2004), except that the procedure was carried out manually instead of using the immunostainer. In brief, the slides were deparaffinized and hydrated as described previously for in situ hybridization. The sections were pretreated for antigen retrieval in a microwave oven (4 × 5 min) using Dako Target Retrieval Solution (DakoCytomation, Glostrup, Denmark). The antibody staining was performed using ChemMate Peroxidase/AEC detection kit (DakoCytomation). The incubation with the CYP2S1 antibody was carried out using a 1:250 dilution of the antibody (30 min at RT). Negative controls were performed by replacing the primary antibody with buffer. The slides were counterstained with hematoxylin. The immunoreactivity was examined by two authors (STS and HW) and scored as negative/marginal, weak, moderate, or strong positive based on the intensity of the immunostaining.
Results
In the present study, we investigated cellular localization of CYP2S1 expression in human tissues using both in situ hybridization and immunohistochemistry. For this purpose, we examined tissue microarrays that included samples from various normal tissues as well as from tumor tissues. The in situ hybridization was performed using a 160-bp RNA probe spanning a specific region on the 3’ end of the CYP2S1 cDNA. In addition, immunohistochemistry was applied for protein expression using a peptide antibody shown to be specific for the CYP2S1 (Rylander et al. 2001). Investigation of both mRNA and protein expression ensured the specificity of the data and comparison between the two levels of gene expression. In serial sections, CYP2S1 mRNA and protein showed comparable expression patterns (Figures 1A and 1B, 1E and 1F, 1G and 1H, 2A and 2B). Both methods indicated expression of CYP2S1 in the majority of the tissues examined. In the positive tissues, the cellular staining was mainly cytoplasmic, but staining was also observed in the nuclei of several tissues. The data on expression of CYP2S1 in different human tissues are summarized in Table 1.
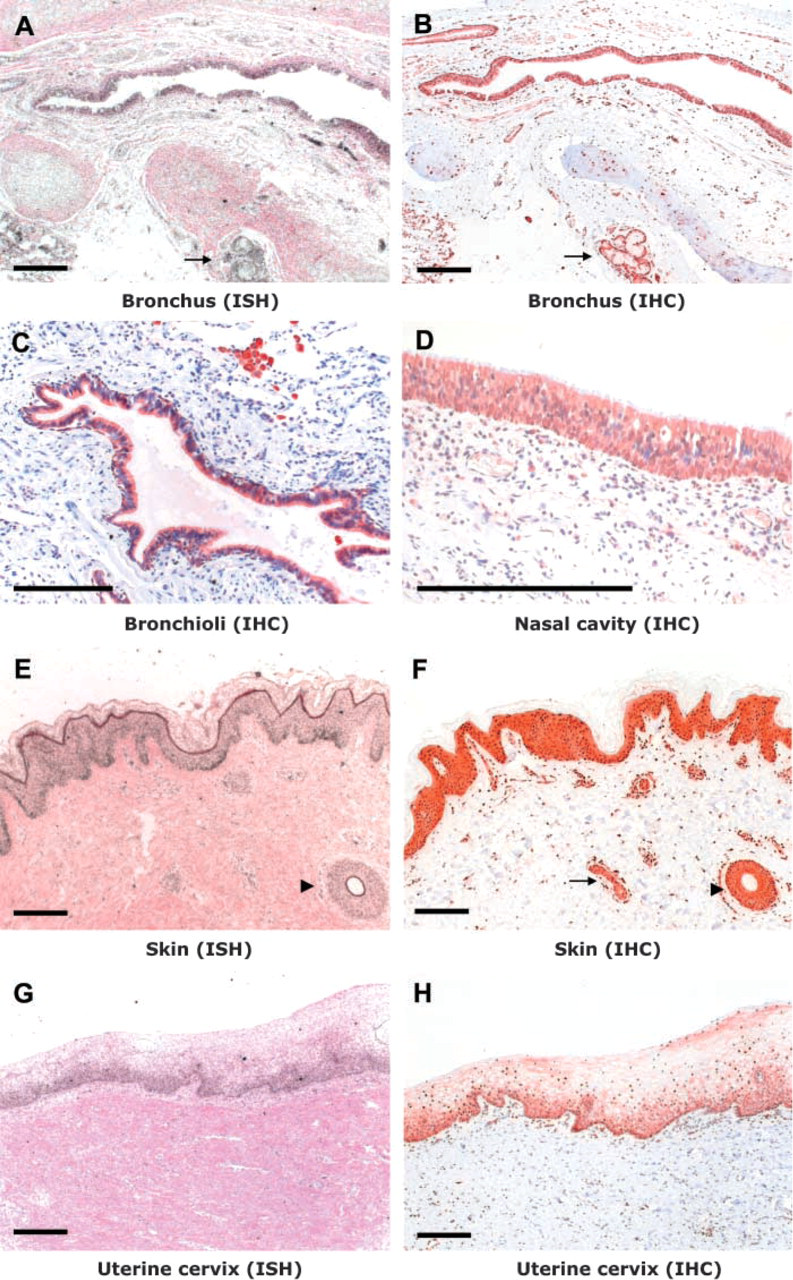
Detection of CYP2S1 expression in human tissues by in situ hybridization to mRNA (ISH) and immunohistochemistry (IHC). Bronchial glands in the lung (
Respiratory Tract
Both in situ hybridization and immunohistochemistry indicated that the expression of CYP2S1 was strong in the epithelia in the upper and lower respiratory tract. In the nasal cavity, CYP2S1 was highly expressed in the respiratory epithelium of the nasal mucosa (Figure 1D). In the lung, CYP2S1 expression was strong in the respiratory epithelium of the bronchus (Figures 1A and 1B) and bronchioles (Figure 1C) in both cuboidal and columnar types of epithelia. The expression was low in the centrilobular area of the peripheral lung. In squamous alveolar epithelial (type I) cells, the expression was either weak or undetectable in all four of the individuals studied, even in samples where the bronchiolar epithelium showed high expression levels. Type II pneumocytes mainly showed no or weak expression of CYP2S1. The bronchial glands showed high expression levels, whereas the expression was weak in the smooth muscle layers around the bronchi (Figures 1A and 1B).
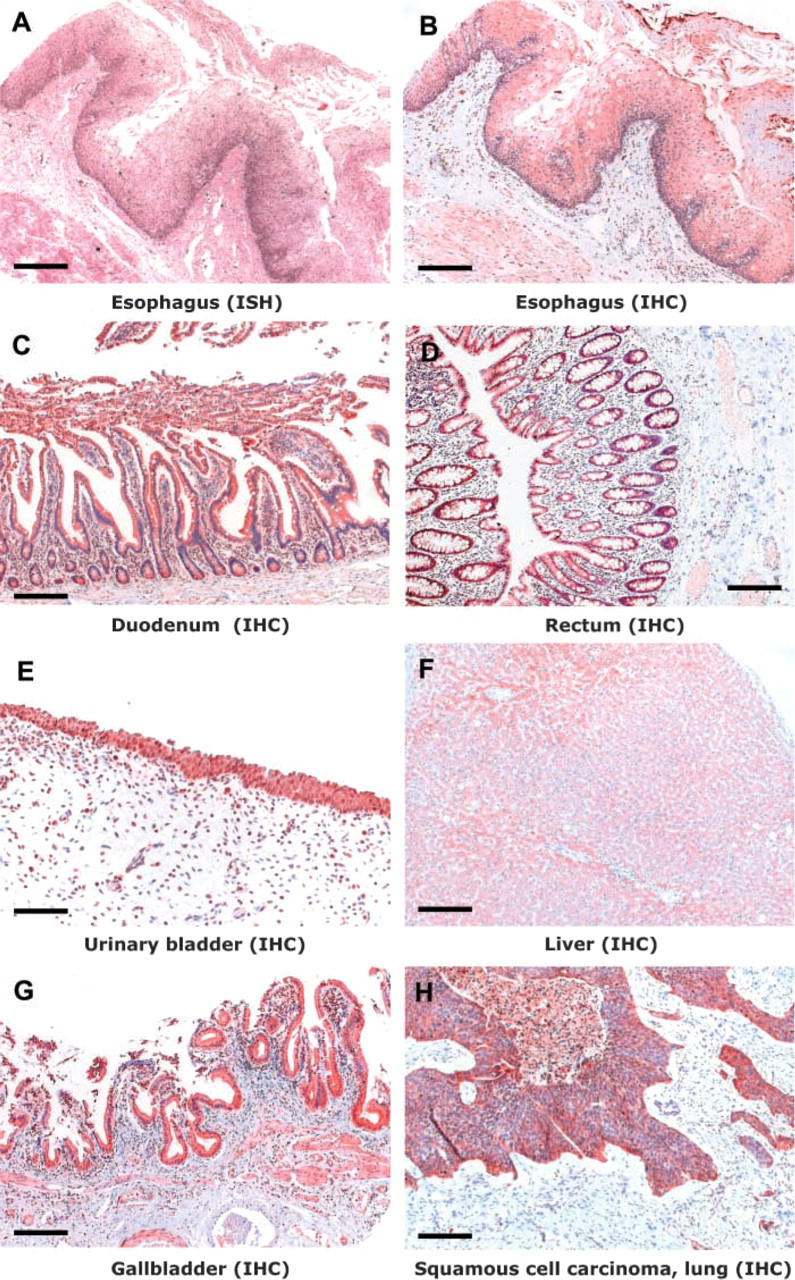
Expression of CYP2S1 in various human tissues. (IHC, immuno-histochemistry; ISH, in situ hybridization). Bar = 100 μm.
Evaluation of staining intensity in epithelial cells of different human tissues

Number of individuals analyzed.
The average intensity of staining graded as absent (-), low (+), moderate (+ +), or strong (+ + +).
One of the two samples strong, the other one negative.
Skin
In the two skin samples analyzed, both mRNA and protein expression were strong throughout the epidermis, except in the cornified layer (Figures 1E and 1F). The epithelial cells in sweat gland and in hair follicle also exhibited strong expression. In situ hybridization, especially, indicated clearly higher grain densities in the basal cells of the germinal layer than in the upper layers of the epidermis. Immunostaining showed that CYP2S1 was expressed in both the cytoplasm and the nuclei.
Reproductive Systems
For the male reproductive system, tissue samples from testis, seminal vesicle and prostate were included (Table 1). The spermatocytes exhibited weak nuclear staining. In the seminal vesicle, weak expression was observed in the columnar epithelium consisting of secretory cells. Weak expression was detected in the epithelium of the prostatic glands where the expression was more intense in the basal cell layer than in the secretory cells of the luminal layer (data not shown).
The female reproductive organs exhibited mainly moderate to low CYP2S1 expression in epithelial cells (Table 1). The uterine cervix expressed CYP2S1 with moderate intensity (Figures 1G and 1H). The intensity of the signal was stronger in the basal layer of the stratified squamous epithelium, the difference in signal intensity being more pronounced with in situ hybridization. Of the two placental samples examined, one exhibited a very strong CYP2S1 expression in the villous epithelium, whereas the other sample was completely negative. In breast tissue, moderate expression levels were observed in the ductal and lobular epithelium.
Gastrointestinal Tract
As in the respiratory tract, CYP2S1 was highly expressed in the epithelial cells throughout the gastro-intestinal tract (Table 1). In the esophagus, the epithelial expression was high, especially in the basal cell layer of the stratified squamous epithelium. This basal expression pattern was more obvious with mRNA in situ hybridization than with the immunohistochemical detection of the protein (Figures 2A and 2B). In the stomach, the expression was strong in the mucus-secreting epithelial cells of the glandular mucosa covering the fundus and body of the stomach (data not shown). Similarly, in the duodenum the expression was strong in the columnar epithelia lining the villi and crypts as well as in the cells surrounding glands in the lamina propria (Figure 2C). The glandular epithelium covering the colon and rectum (Figure 2D) also showed high expression levels.
Urinary System
Of the three kidney samples studied, two had marginal or low CYP2S1 expression levels, but a moderate level of cytoplasmic expression was detected in one sample (data not shown). The expression was more intense in the distal convoluted tubules than in the proximal ones. In the urinary bladder, strong expression was observed in the urothelium in both the two samples of different origin (Figure 2E; the sample exhibits signs of infection).
Liver
All three of the normal liver samples analyzed exhibited CYP2S1 expression. The expression was low in two samples and moderate in one sample. The expression was higher around the central and portal veins than in the parenchyma (Figure 2F). CYP2S1 expression was often concentrated in the granules of the hepatocytes.
Exocrine and Endocrine Glands
In addition to the gastrointestinal glands discussed previously, CYP2S1 was also moderately or highly expressed in the epithelial cells of many other endogenous and exogenous glands, including gallbladder, pancreas, adrenal gland, and salivary glands (Table 1). In the two gallbladder samples studied, the simple columnar epithelium showed moderate or strong expression (Figure 2G). In the exocrine pancreas, CYP2S1 expression was strong, especially in the secretory cells of the acini. The staining was most intense in the cells located in the periphery of the pancreatic islets. In the cuboidal epithelium of the ducts, the expression was moderate. The endocrine tissue of the pancreas (i.e., the islets of Langerhans) also showed only moderate immunostaining. In the salivary gland, CYP2S1 expression was restricted to the columnar epithelial cells of the excretory ducts, the secretory cells being negative. The cortex of adrenal gland exhibited strong CYP2S1 expression, whereas the expression was moderate in the medulla (data not shown).
Other Non-tumorous Tissues
The organs of the immune system (i.e., thymus, spleen and lymph nodes) exhibited a uniform, weak expression (data not shown). Lymphocytes present in tissues suffering from inflammation showed relatively high expression levels, however. CYP2S1 expression was detected in endothelial cells of blood vessels throughout the tissues examined. In many tissues, weak to moderate expression was also detected in the smooth muscle cells surrounding the small arterioles. In the brain tissue samples, weak staining was detected in the nuclei of the white matter.
Tumor Tissues
The expression of CYP2S1 in different tumors varied from undetectable to strong. High CYP2S1 expression levels were observed in one of the two cases of lung squamous cell carcinomas studied, whereas the other sample of lung squamous cell carcinoma and the adenocarcinoma specimen exhibited only weak or undetectable expression levels, respectively. Strong expression was also observed in the squamous cell carcinoma of the uterine cervix as well as in the two tumors detected in the ovaries (i.e., a metastatic adenocarcinoma from the appendix and a mucinous cystadenocarcinoma).
Discussion
In the present study, we used in situ hybridization and immunohistochemistry to study the cellular localization of CYP2S1 mRNA and protein in various human tissues. These methods enabled us to determine the specific cell types expressing CYP2S1 as well as the subcellular localization of the expression in semiquantitative fashion. Our results indicate that CYP2S1 is expressed in a variety of healthy human tissues, the expression being mainly restricted to the epithelial cell types. Strong CYP2S1 expression was detected in both simple and stratified types of epithelium as well as in the endothelium. Expression was often intense, especially in the germinal basal cell layers. Considerable expression of CYP2S1 was also detected in many organs that develop as invaginations of epithelial surfaces, such as kidney and exocrine and endocrine glands.
The epithelial expression of CYP2S1 was high, especially in many tissues that are exposed to xenobiotics (e.g., the respiratory, gastrointestinal, and urinary tracts and the skin). Previous studies have reported Northern blotting and RT-PCR results showing that CYP2S1 is highly expressed, especially in the lungs (Rylander et al. 2001; Rivera et al. 2002; Smith et al. 2003; Choudhary et al. 2003). Our earlier data indicated that CYP2S1 is constitutively expressed in the lungs, and the mRNA levels were increased by threefold after dioxin treatment in mouse lung tissue and in a human lung cell line (Rivera et al. 2002). Our present results enabled us to localize the pulmonary expression to epithelia of bronchi and bronchioles. In contrast, the expression was weak in the alveolar lining cells. We have previously observed a similar pulmonary expression pattern for another AHR-dependent, PAH-inducible CYP gene, CYP1A1 (Saarikoski et al. 1998). Our present findings showing strong CYP2S1 expression in cells that are exposed to inhaled toxic and carcinogenic compounds (e.g, those present in cigarette smoke) is interesting, because these cells are also common progenitors for tumor initiation. Indeed, high expression of CYP2S1 was observed in lung squamous cell carcinoma.
The gastrointestinal tract serves as the portal of entry for orally ingested xenobiotics, including therapeutic drugs as well as those present in food. Therefore, it is noteworthy that strong epithelial expression was observed throughout the gastrointestinal system, both in the stratified epithelium of the esophagus and in the glandular epithelium of the lower parts of the digestive tract (i.e., stomach, duodenum, small bowel, colon, and rectum). The high epithelial expression of CYP2S1 in the digestive tract resembles the expression pattern observed previously for CYP1A1 in mice treated with 3-methylcholanthrene (Dey et al. 1999).
In addition to the portal-of-entry tissues that are exposed to a considerable load of foreign compounds, CYP2S1 was also highly expressed in many tissues that are not directly exposed to xenobiotics, such as pancreas and adrenal gland. This finding is in agreement with the previous Northern blotting and RT-PCR data on the tissue distribution of CYP2S1 expression (Rylander et al. 2001; Rivera et al. 2002; Smith et al. 2003; Choudhary et al. 2003). The constitutive and relatively ubiquitous expression pattern is in line with the accumulating evidence of CYP2S1 involvement in metabolism of polycyclic aromatic hydrocarbons (Rivera et al. 2002; Smith et al. 2003; Karlgren et al. 2005) as well as all-trans retinoic acid (Smith et al. 2003), indicating a role in both exogenous and endogenous metabolism. In addition, ultraviolet irradiation has been reported to induce CYP2S1 in the skin (Smith et al. 2003), where we detected very strong expression.
When compared with the tissue distribution obtained from other CYPs, the tissue-distribution pattern of CYP2S1 expression most resembles those observed for other AHR-regulated CYPs of the CYP1 family, especially CYP1B1 (Choudhary et al. 2003; Ding and Kaminsky 2003). The extrahepatic expression pattern detected in this study for CYP2S1 follows the immunohistochemical localization reported earlier for human CYP1B1 (Muskhelishvili et al. 2001). Like CYP2S1, CYP1B1 was expressed either in cytoplasmic or nuclear cellular compartments or both. In addition to the human data, RT-PCR-based studies on mice have indicated that CYP1B1 and CYP2S1 share a similar expression pattern (Choudhary et al. 2003). Because retinoids are known to be crucial for embryonic development (Marill et al. 2003), CYPs that are known to participate in metabolism of retinoids, including CYP1B1, have been suggested to play an important role in embryogenesis (Stoilov et al. 2001; Otto et al. 2003). Experiments on mice have shown that CYP2S1 is one of the few CYP isoforms (6/40) that was expressed in all fetal stages studied, and it was consequently postulated to have a specific role in embryonic development (Choudhary et al. 2003).
Of the genes belonging to the CYP2 gene cluster at 19q13.2, CYP2A6 and CYP2B6 are predominantly expressed in the liver, whereas CYP2A13 and CYP2F1 are highly expressed in the respiratory tract, as is CYP2S1 (Hukkanen et al. 1997; Su et al. 2000; Ding and Kaminsky 2003; Nishimura et al. 2003). CYP2A13 and CYP2F1, however, unlike CYP2S1, do not exhibit strong expression in other extrahepatic tissues. Cyp2a5, the mouse homologue of CYP2A6, has recently been found to exhibit dioxin inducibility mediated by AHR and ARNT (Arpiainen et al. 2004).
High expression levels of various CYPs have been found previously in many tumors. In addition to biotransformation of carcinogenic compounds, CYPs have also been suggested to convert endogenous substrates to metabolites that facilitate cancer development and to participate in the metabolism of anticancer drugs (Murray 2000; Oyama et al. 2004). Of the different CYP isoforms, overexpression of CYP1B1 is the one most often detected in various tumors (Murray 2000). Our study also showed that CYP2S1 is highly expressed in many tumors of epithelial origin. The expression levels are similar to those in the corresponding healthy epithelia, indicating maintenance of expression levels detected in the epithelial progenitor cells.
CYP2S1 exhibits both constitutive and inducible expression characteristics (Rivera et al. 2002; Smith et al. 2003). Within our study material, some interindividual differences were observed in the expression of CYP2S1. This variation may be caused by differential exposure of the individuals to CYP2S1 inducers. Because CYP2S1 is inducible by PAHs (Rivera et al. 2002; Smith et al. 2003; authors’ unpublished data), exposure to tobacco smoke might have affected the expression of CYP2S1. To address the possible modifying role of cigarette smoke, we examined CYP2S1 mRNA and protein expression in a separate set of placental samples from smoking and nonsmoking women and found that cigarette smoking has no major effect on CYP2S1 expression (Hakkola et al., unpublished data). Differential exposure to tobacco smoke is thus not likely to explain the observed variation in the tissue array specimens. In addition, genetic variation may lead to interindividual differences in the expression levels. After recent identification of genetic polymorphisms of the human CYP2S1 (Saarikoski et al. 2004), we continue the work with further studies investigating the role of these variations on the expression of CYP2S1.
To conclude, CYP2S1 expression was mainly restricted to epithelial cell types. High expression levels were observed, especially in the epithelia of xenobiotic-exposed tissues, such as the respiratory, gastro-intestinal, and urinary tracts and the skin. CYP2S1 was expressed also, however, in other organs of epithelial original (e.g., some exocrine glands), thus supporting other data pointing to the importance of CYP2S1 in the metabolism of endogenous substances. This ubiquitous epithelial expression pattern is particularly interesting in view of the involvement of CYP2S1 in retinoic acid metabolism and the importance of retinoids for differentiation and proliferation of epithelial cells. Furthermore, it is noteworthy that epithelial cells are also the major targets for carcinogen exposure and constitute common progenitor cells to tumor development. Indeed, CYP2S1 was highly expressed in many tumors of epithelial origin.
Footnotes
Acknowledgements
This work was financially supported by the Academy of Finland (grant no. 52276) and the Work Environment Fund (grant no. 100389).