
Abstract
5-Bromo-2'-deoxyuridine (BrdU) and 5-chloro-2'-deoxyuridine (CldU) were sequentially administered intraperitoneally into mice at 1-hr intervals. After one additional hr, the small intestines were resected, fixed, and embedded in paraffin. In histological sections stained with monoclonal antibody Br-3 reactive to both BrdU and CldU, and CldU antibody reactive only to CldU, three types of staining could be identified in the proliferating zone. Cells with nuclei stained only with Br-3 antibody were estimated to have completed DNA replication during the first 1 hr and were fixed in G2/M-phase. Those nuclei were frequently found in apical areas of the simple columnar epithelium of the intestine, whereas other nuclei were located basally in the cells. This observation suggested intracellular movement of cell nuclei in G2/M-phase. Identification of cells in early S-phase became possible using these antibodies in combination with DAB and fluorescence stainings. Replication sites in early S-phase nuclei were found to be numerous, whereas in late S-phase they were larger in size and much smaller in number.
I
Materials and Methods
Antibody
Ten mg of 5-fluorouridine (Sigma-Aldrich; St Louis, MO) was dissolved in 500 μl of distilled water and mixed with 10 mg of sodium periodate dissolved in 500 μl of distilled water. After 30 min, 10 mg of rabbit serum albumin (Calzyme Laboratories; San Luis Obispo, CA) dissolved in 500 μl of 0.2 M phosphate buffer (pH 7.4) was added and the mixture was left for 2 hr. After addition of 5 mg of sodium boro-hydride, the 5-fluorouridine-RSA conjugate was dialyzed against 20 mM PBS (pH 7.4) overnight, diluted to make up 10 ml of PBS solution, and stored frozen until use. One ml of the conjugate was mixed with 1 ml of Freund's adjuvant and injected monthly into the back of a rabbit. The immunoglobulin fraction prepared from antiserum obtained from the rabbit was added with BrdU (approximately 6 mg/10 ml antiserum) and left overnight. The mixture was then applied to an affinity column coupled with 5-fluorouridine on amino cellulofine (Seikagakukogyo; Tokyo, Japan). The column was washed and eluted with glycine-HCl buffer (pH 2.5) to obtain fractions reactive with CldU. The eluate was again applied to a BrdU affinity column to remove BrdU-reactive immunoglobulins. The final pass-through fraction was called the CldU antibody in the present study.
DNA Double Labeling
Male mice aged 4 or 10 weeks (Slc/ICR) were injected intraperitoneally with 50 mg/kg body weight of BrdU (TCI Tokyo Kasei Kogyo Co., Ltd.; Tokyo, Japan) dissolved in physiological saline. After 1 hr, they received an injection of 50 mg/kg of CldU (ICN Biomedicals; Aurora, OH). After an additional hr, the animals were sacrificed under deep anesthesia, and the small intestines were removed, fixed in 70% ethanol, and routinely processed to 5-μm-thick paraffin sections.
To establish the specificity of the antibody, mice injected with either CldU or BrdU were prepared and their intestines were used for examination of binding.
Immunohistochemistry
After deparaffinization and rehydration through graded series of ethanol, sections were pretreated in a microwave oven in sodium citrate buffer (pH 6.0) for 5 min at 100C. After they were cooled down in NaCl-TB (300 mM NaCl in 50 mM Tris buffer, pH 7.4) for 5 min, they were incubated with 1% skimmed milk in NaCl-TB for 5 min to block nonspecific binding.
For double staining with DAB and fluorescent dye, sections were incubated with CldU antibody (10 μg/ml) overnight. After they were exposed to biotinylated goat anti-rabbit IgG (Lab Vision Corporation; Fremont, CA) for 60 min, they were incubated with 1:2000 diluted horseradish peroxidase avidin D (Vector Laboratories; Burlingame, CA) for 60 min, then 0.02% of 3, 3'-diaminobenzidine tetrahydrochloride (DAB, Dojinkagaku, Kumamoto, Japan) in 100 mM imidazole-HCl solution, pH 7.4, with 300 mM NaCl and 0.006% of H2O2 for 15 min. After they were washed, they were treated again in a microwave oven in sodium citrate buffer for 5 min at 100C and incubated with Br-3 antibody (1:10000) overnight, followed by Alexa Fluor 594 goat antimouse IgG (1:400; Molecular Probes, Eugene, OR) in NaCl-TB with skimmed milk for 3 hr. Finally they were mounted in mounting medium containing DAPI (Vector Laboratories).
For double immunofluorescent staining, after pretreatment in a microwave oven, sections were incubated with CldU (10μg/ml) and Br-3 (1:10000) antibodies overnight. Then the sections were incubated with a mixture of Alexa Fluor 594 goat anti-rabbit IgG (1:400) and Alexa Fluor 488 goat anti-mouse IgG (1:400) for 3 hr and mounted in the medium containing DAPI. Sections from mice injected with either CldU or BrdU were also treated with the same method to determine reactivities of antibodies used in the present study.
All stained sections were examined under a fluorescent microscope equipped with a digital camera (Nikon DXM 1200; Tokyo, Japan). Photographs of DAB stained sections were treated with a computer program (Photoshop) to reverse the tone of the pictures and merged with corresponding fluorescence images.
DNA Double Labeling of Cultured Cells
Neural progenitor cells were cultured according to the method described by Shinohara et al. (2004). Dissociated cells (4–5 × 105/dish) were seeded onto 35-mm dishes with 12 × 9 mm coverslips precoated with poly-L-lysine (Sigma) and propagated in serum-free medium (SFM) with fibroblast growth factor-2 (FGF-2) for 24 hr. After 24 hr, some cultured cells were incubated in SFM with FGF-2 and BrdU (10 μM) for 60 min. After the incubation, the culture medium was exchanged to SFM with FGF-2 and CldU (10 μM), and the cells were cultured for another 60 min (pulse-chase-pulse-chase experiment). Other cultured cells were incubated in SFM with FGF-2 and BrdU (10 μM) for 60 min. Then CldU (10 μM) was added to the culture medium and the cells were incubated for another 60 min (no-chase experiment). Finally, the cultured cells were fixed in 70% ethanol for 20 min and then stored in PBS with sodium azide until use.
The methods of double immunostaining were the same as those of histological sections.
Estimation of Cell Cycle Length
To calculate the duration of S-phase (Ts) and the length of a cell cycle (Tc), we counted the number of nuclei of the intestinal epithelial cells that were stained only with Br-3 antibody, and with both Br-3 and CldU antibodies, respectively, and the total number of nuclei comprising the proliferating zone in the same section. The duration of S-phase (Ts) was calculated as the number of double stained cells/Br-3 stained cells × t, where t is the time interval between injection of the two labels (in this study 1 hr). The length of a cell cycle (Tc) was expressed as Ts/CldU labeling index (LI) × GF, where GF is the “growth fraction,” assumed to be 100% in this case. The CldU LI is the percentage of CldU-labeled nuclei among the DAPI positive nuclei.
Results
The CldU antibody recovered from a 5-fluorouridine affinity column and passed through a 5-bromouridine affinity column proved reactive with nuclei of CldU injected mice but not those of BrdU-injected mice, indicating high specificity to CldU. On the other hand, Br-3 was reactive to both BrdU and CldU (Figure 1). Specimens from non-injected mice were not immunostained with either CldU or Br-3 antibodies, and positive immunostaining was not seen when the primary antibody was replaced by non-immune mouse or rabbit IgG (Figure 2).
Specificity of CldU and Br-3 antibodies. (
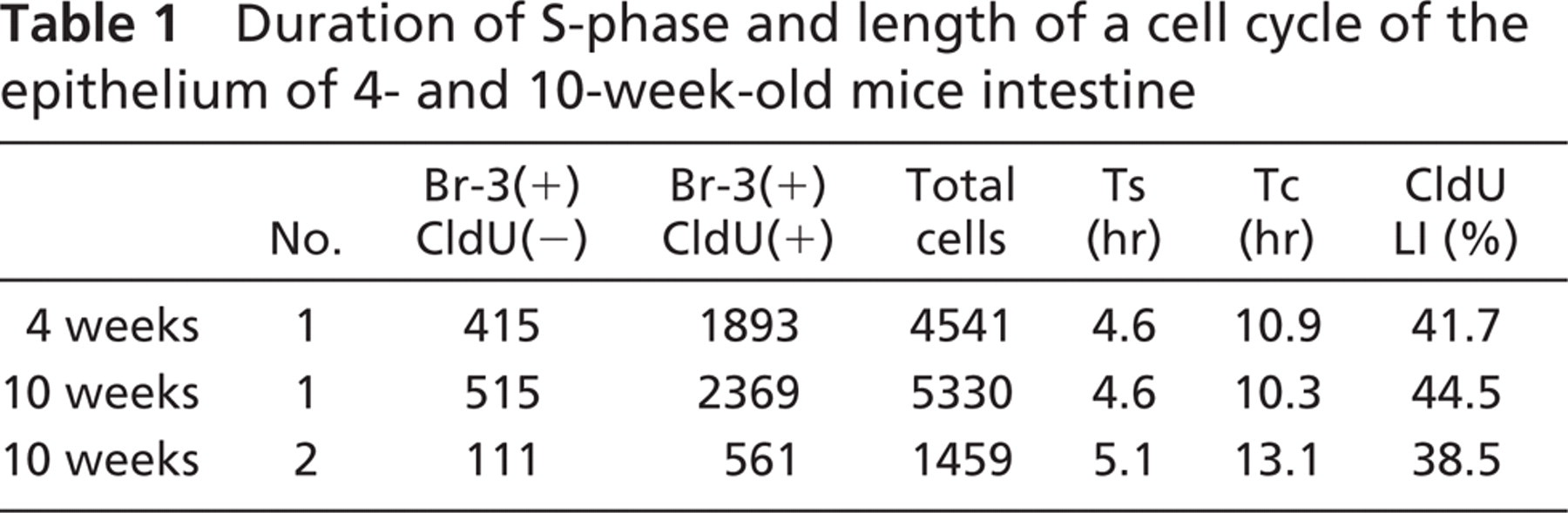
When a mouse was injected with BrdU, it was incorporated by cells in S-phase (Figure 3A). After the mouse was injected with CldU, cells in S-phase incorporated both BrdU and CldU (Figure 3B). However, cells that had finished DNA replication between the injections did not incorporate CldU (Figure 3B). Numbers of nuclei stained only with Br-3 antibody and with both Br-3 and CldU antibodies, and those labeled with DAPI (i.e., total number of cells) were counted in 4- and 10-week-old mice (Table 1). On the cultured cells of the pulse-chase-pulse-chase experiment and the no-chase experiment, numbers of nuclei stained only with Br-3 antibody and with both Br-3 and CldU antibodies, and those labeled with DAPI were counted similarly (Table 2). Based on the data, the durations of S-phase and a cell cycle were calculated for each mouse and culture experiment. The duration of S-phase (Ts) and the length of a cell cycle (Tc) of the histological sections of one 4-week-old and two 10-week-old mice were 4.6, 4.6, 5.1 hr and 10.9, 10.3, 13.1 hr, respectively (Table 1). On the other hand, Ts and Tc of the cultured cells of pulse-chase-pulse-chase experiment and no-chase experiment were 13.3 and 12.5 hr and 31.8 and 29.7 hr, respectively (Table 2).
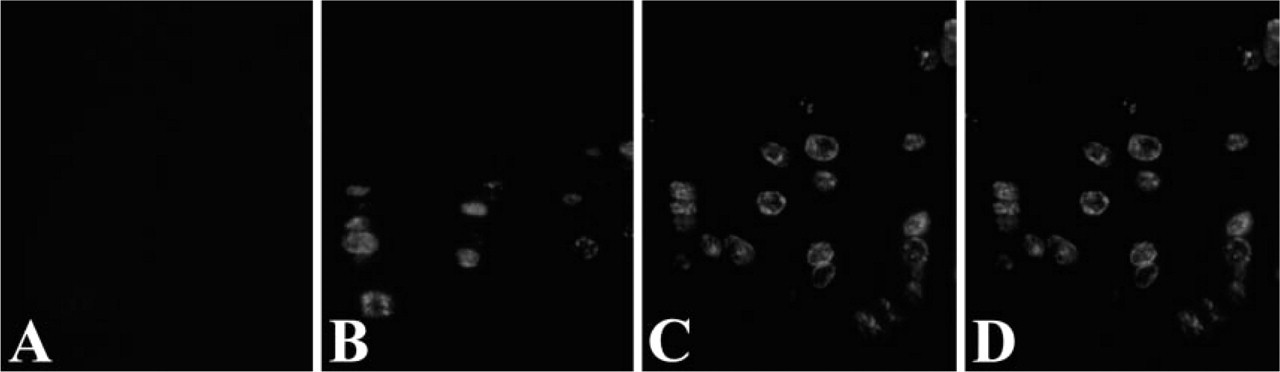
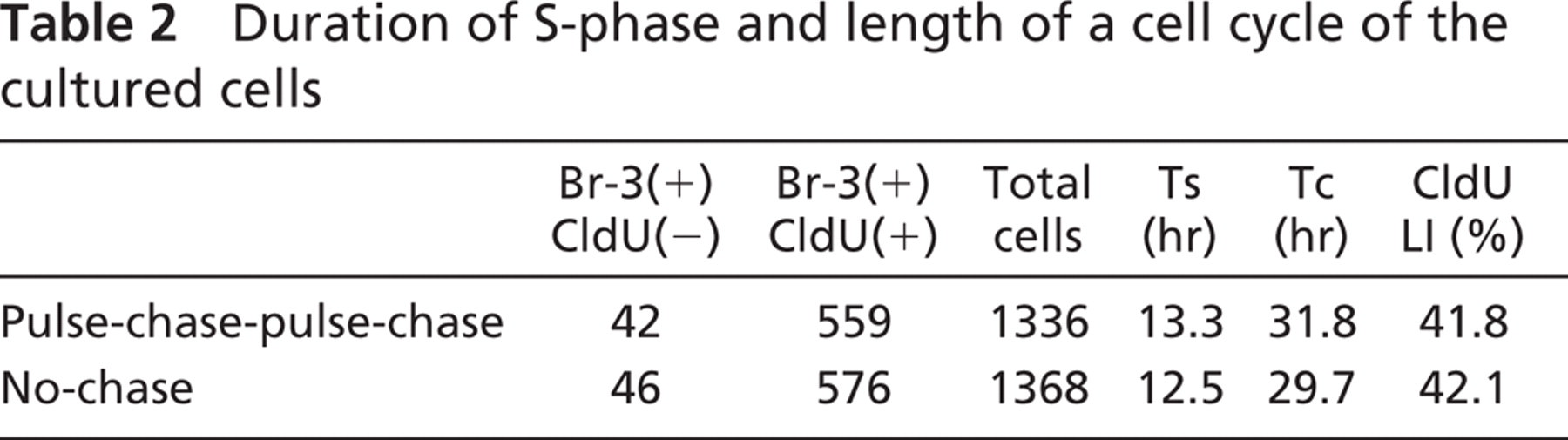
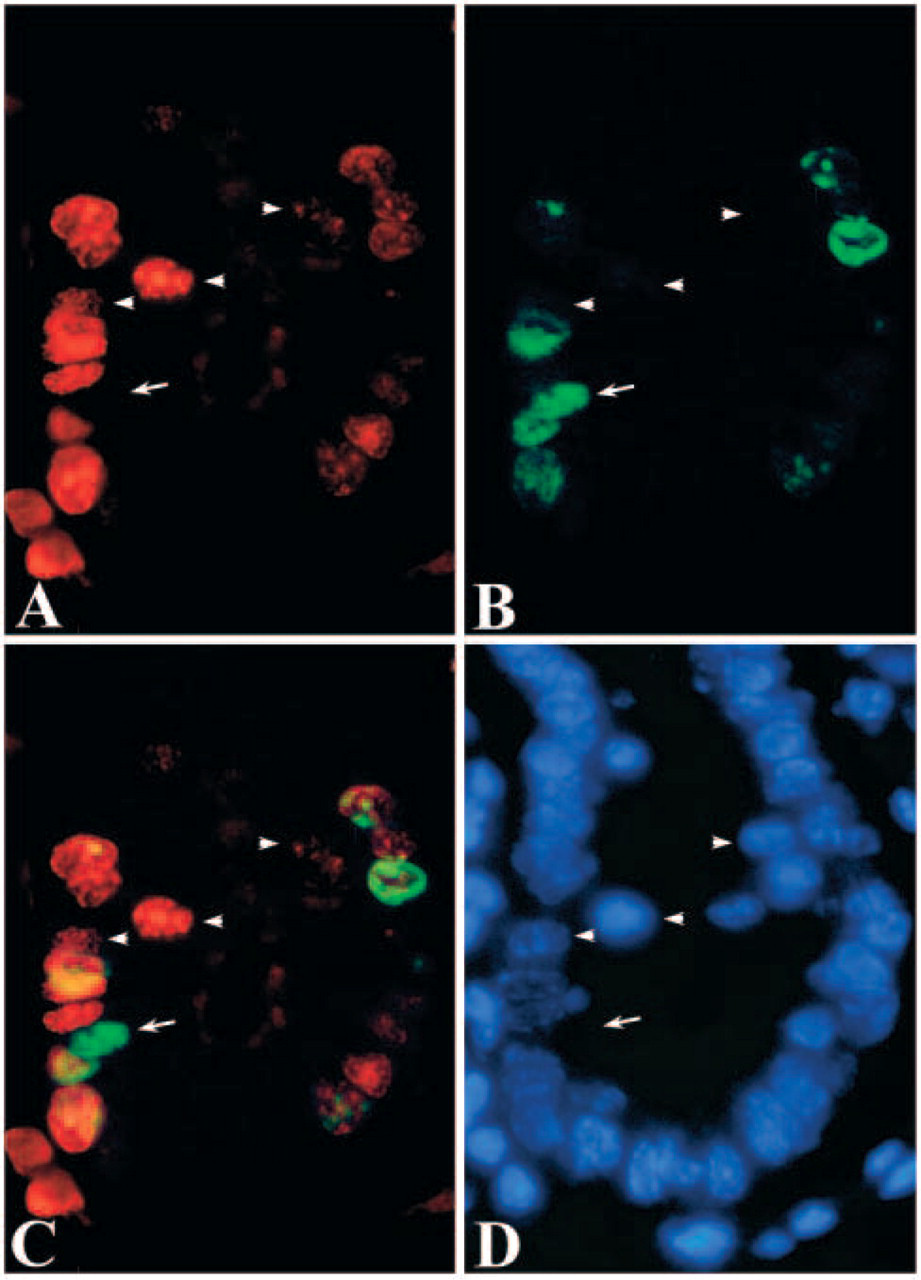
When the two photographs of DAB and fluorescent dye were merged, three types of cells were identified; those stained only with the fluorescent dye, those stained only with DAB and those stained with both DAB and the fluorescent dye (Figure 4C). They were estimated to be the cells which had finished DNA replication in the first 1 hr, cells entering S-phase within the second 1 hr, and those in mid to late S-phase, respectively. On the other hand, two types of cells were recognized in pulse-chase-pulse-chase experiment and no-chase experiment of the cultured cells; those stained only with the green fluorescent dye and those stained with both the red and green fluorescent dyes (Figure 5). In higher magnification of the sections and the cultured cells, stainings of the cells that had finished DNA replication in the first 1 hr were sometimes found as a small number of relatively large dots localized at the periphery of the nuclei (Figure 4 and Figure 5). In histological sections, these nuclei were frequently found to be localized in the apical regions of the cytoplasm while other nuclei were found in basal regions of the cells (Figure 4).
Negative controls of the non-injected mice (
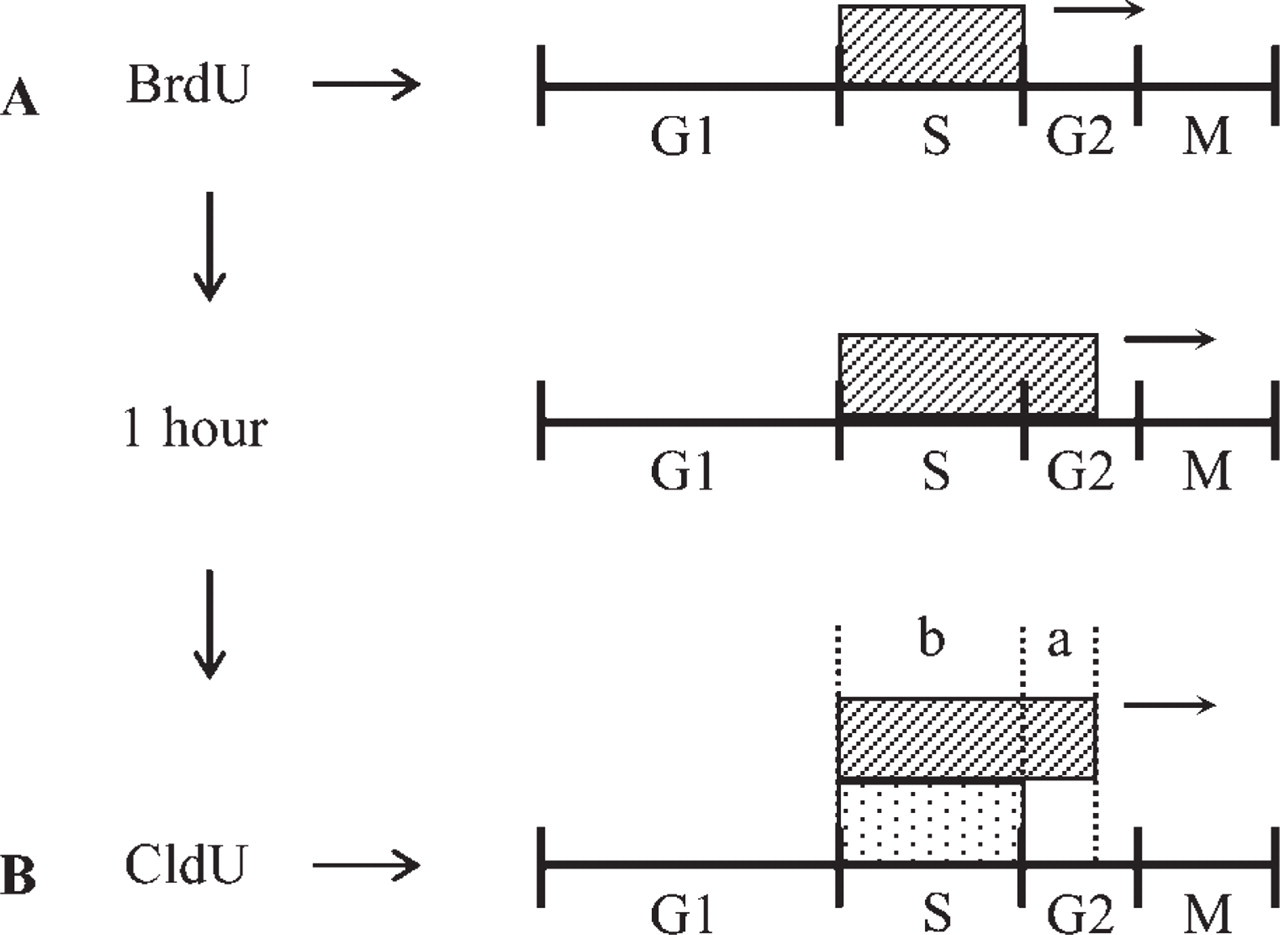
Findings after injection of both BrdU and CldU. When BrdU was injected, it was incorporated by cells in S-phase (
Discussion
Thymidine analogs such as BrdU, IdU and CldU are known to be incorporated into cellular DNA during DNA replication, and have been widely used for identification of proliferating cells (Vanky et al. 1998; Calvo et al. 2000; Uni et al. 2000). They have also been employed to reveal the duration of S-phase and the length of a cell cycle (Nowakowski et al. 1989; Takahashi et al. 1993; Kornack and Rakic 1998). Although BrdU and IdU have been used for double labeling of proliferating cells, IdU is difficult to dissolve in water and a large volume of solution is necessary for administration to mice. Moreover, it was difficult in our hands to obtain antibodies reactive to IdU but not to BrdU from IdU-conjugate injected chicken. In contrast, CldU is readily soluble in water and the volume for injection to mice is small, so that its application is much easier. In addition we could readily purify an antibody fraction reactive to CldU but not to BrdU from our 5-fluorouridine-conjugate injected rabbit (Figure 1). Using the CldU and Br-3 antibodies, it was here possible ble to count the number of nuclei stained by each of the antibody and calculate the duration of S-phase and length of a cell cycle in one animal. The values obtained were almost the same as those reported from previous laborious work (Potten et al. 1979). Because BrdU may remain in the animal body after the second injection (CldU), cells in S-phase incorporate both CldU and BrdU. We then tried a pulse-chase-pulsechase experiment of cultured cells and compared the result with that of a no-chase experiment. The duration of S-phase and the length of a cell cycle in the cultured cells were not different between the two experiments (Table 2). It seems that the present CldU antibody and application of it to a double labeling study is useful and reliable for calculating the duration of S-phase or cell cycle in histological tissue sections as well as in cultured cells.
Duration of S-phase and length of a cell cycle of the epithelium of 4- and 10-week-old mice intestine
Duration of S-phase and length of a cell cycle of the cultured cells
Double staining with CldU and Br-3 antibodies in mice. (
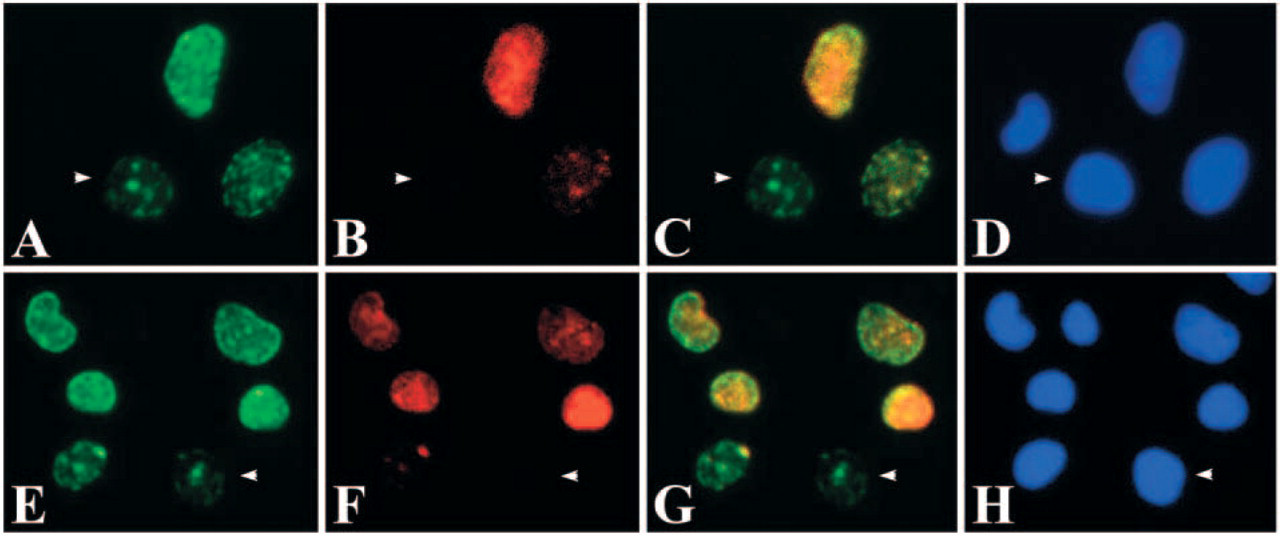
Double staining with CldU and Br-3 antibodies in cultured cells. (
Identification of cells in early S-phase has been performed in cultured cells (Nakayasu and Berezney 1989; O'Keefe et al. 1992; Manders et al. 1992, 1996; Ma et al. 1998). In the present study, identification of those cells in tissue sections was attempted. It is known that eukaryotic chromosomal DNA is divided into hundreds to thousands of independent subunits of replication named replicons (Hand 1978). When BrdU and CldU were incorporated into a replication site simultaneously, DAB pigmentation due to CldU staining can quench the colocalized fluorescence due to BrdU, while when they were incorporated sequentially, fluorescence of BrdU will not be quenched by subsequent DAB staining because the replication sites moved during the 1 hr interval between the administrations. In the present study, cells entered the S-phase during the second 60 min were expected to incorporate both BrdU and CldU simultaneously and will be stained only with DAB (Figure 4). In practice, cell nuclei stained only with DAB were found and the staining pattern was diffuse suggesting many replication sites. In previous reports on cultured cells, replication sites in early S-phase consisted of many small domains scattered throughout the nucleoplasm, whereas in late S-phase they were larger in size and smaller in number (Manders et al. 1992, 1996; Ma et al. 1998). An electron microscopic study revealed that cells in early S-phase display a diffuse pattern of labeling with many spots, whereas cells in late S-phase show labeling confined to large domains, often at the periphery of the nucleus or associated with nucleolus (Jaunin et al. 1998). Thus, the present observation was in good agreement with these reports on cultured cells (Figure 4 and Figure 5).
Considering the good correlation between the present double-labeling study and the previous ones, we can conclude that our double labeling technique using CldU and BrdU in combination with DAB and fluorescent staining enabled to identify cells in early S-phase in tissue sections. By shortening the interval between the 2nd injection and the fixation, it will be possible to identify the cells in very early S-phase.
In embryos, cell nuclei of the neuroepithelium, a kind of pseudostratified epithelium, are positioned in a variety of levels and were reported to change the level depending on their cell kinetics (Fujita 1962). In the present study, most of nuclei of the intestinal columnar epithelium are positioned in basal level, while nuclei stained only with Br-3 antibody were frequently found in apical level (Figure 4). Because the cells stained only with Br-3 antibody were thought to have finished DNA synthesis during the first 60 min, the displaced nuclei stained only with Br-3 antibody suggest intracellular movement of nuclei in the G2- or M-phase in columnar epithelium. Because the displaced nuclei were frequently not oval in shape, they may be in M-phase.
Footnotes
Acknowledgements
We wish to thank Dr. Tomiko Asano (Institute for Developmental Research, Aichi Human Service Center) for her support through the cell culture experiment.