
Abstract
We aimed to examine the distribution of SEPT4, SEPT5, and SEPT8 in the human eye. For each septin, five to six normal human eyes were examined by immunohistochemical staining of paraffin sections using polyclonal antibodies against SEPT4, SEPT5, and SEPT8 and an avidin biotin complex immunodetection system. SEPT4 immunoreactivity (IR) was detected primarily in the epithelium of cornea, lens, and nonpigmented ciliary epithelium; in the endothelium of cornea and vessels of iris and retina; and in the retinal nerve fiber layer, the outer plexiform layer, the outer segments of the photoreceptor cells, the inner limiting membrane of the optic nerve head, and optic nerve axons. SEPT5-IR was present in corneal endothelial cells, iris tissue, nonpigmented ciliary epithelium, and epithelial cells of the lens. SEPT8-IR almost paralleled that of SEPT4, except for a lower SEPT8-IR of the outer photoreceptor segments and a positive staining of the meningothelial cell nests in the subarachnoidal space of the bulbar segment of the orbital optic nerve. In conclusion, SEPT4, SEPT5, and SEPT8 are expressed in various ocular tissues, each revealing a distinct expression pattern. Both physiological and potential pathophysiological role of septins in the human eye deserve further investigation.
S
Ultrastructurally, mammalian septin filaments appear an irregular array of structural segments with obscure polarity. They have the potential to self-assemble into higher order structures by lateral stacking and tandem annealing, eventually forming uniformly curved bundles such as rings and coils. Septin-based structures may rearrange and disassemble in cells under control of diverse factors and covalent modifications; however, the exact mechanisms of assembly and disassembly have yet to be disclosed (Kinoshita 2003).
The range of properties of septins is remarkable. They appear to be essential for active membrane movement such as cytokinesis, cell polarity determination and maintenance, microtubule and actin function, cell movement, vesicle trafficking, exocytosis, and apoptosis (Hall and Russell 2004; Martinez and Ware 2004). Moreover, they play a role in oncogenesis (Longtine et al. 1996; Huizinga et al. 1997; Beites et al. 1999; Montagna et al. 2003; Hall and Russell 2004). In non-dividing cells, such as platelets and neurons, septins are implicated in exocytosis (Martinez and Ware 2004). Platelets from a SEPT5 knockout mouse showed an altered serotonin secretion and platelet aggregation, suggesting that SEPT5 is involved in granular secretion in platelets. Meanwhile, three platelet septins are known: SEPT4, SEPT5, and SEPT8 (Blaser et al. 2004).
Expression of SEPT5 and SEPT8 has also been demonstrated in human prostate, testis, and ovary, where they may be involved in exocytosis (Blaser et al. 2003). Moreover, SEPT5 and SEPT8 have been identified as interaction partners in brain and heart (Blaser et al. 2003). SEPT5 is also involved in serotonin release in platelets and may regulate synaptic vesicle dynamics (Zhang et al. 2000; Dent et al. 2002). Megonigal and colleagues demonstrated that acute myeloid leukemia was caused by the genomic translocation of MLL with the SEPT5 gene (Megonigal et al. 1998). It has been shown for SEPT4 that an alternatively processed mRNA results in cellular apoptosis (Larisch et al. 2000). Septins also accumulate into pathological cytoplasmic structures in common human neurodegenerative diseases such as neurofibrillary and glial tangles in Alzheimer's disease (Kinoshita et al. 1998) and Lewy bodies and glial cytoplasmatic inclusions in Parkinson's disease (Ihara et al. 2003). SEPT5 is one of the substrates of the E3 ubiquitin ligase parkin, whose loss-of-function mutations may cause juvenile parkinsonism (Kinoshita 2003). In a proteomic approach, abnormalities of septin expression have also been reported in Down syndrome (Cheon et al. 2001).
In a first attempt to disclose distribution and possible (patho-)physiological roles of the septins in the human eye, we examined SEPT4, SEPT5, and SEPT8 immunoreactivity (IR) by means of immunohistochemistry. We also compared the labeling pattern of these septins with that of the cell cycle-related protein proliferating cell nuclear antigen (PCNA) and the synaptic vesicle protein synaptophysin.
Materials and Methods
Tissue Preparation
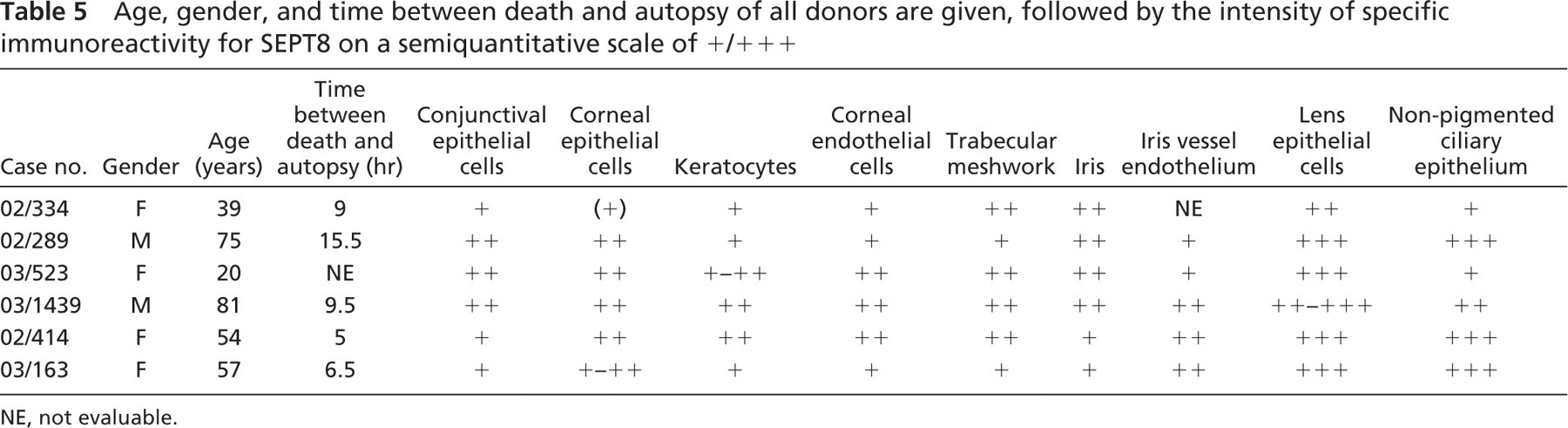
Human Tissue Samples. Paraffin-embedded human eyes, fixed in 4% paraformaldehyde, were obtained postmortem from six patients without ophthalmological or neurological diseases (M:F = 2:4; mean age: 54 years ± 23 SD) (Table 5). The cause of death was either heart failure or pneumonia or intracerebral sanguination in all cases. The sample collection was approved by the Ethics Committee criteria and followed the tenets of the Helsinki Declaration. Tissue samples were cut in a sagittal plane including the optic nerve head and pupil, and 4 μm-thick serial sections were made with a microtome.
Immunohistochemistry
Sections were mounted on gelatin chromalum-coated glass slides and deparaffinized. The antigen was finally visualized by peroxidase staining using the peroxidase substrate 3-amino-9-ethylcarbazole. The optimum concentration of the primary antibody was previously determined to be 1:200. The polyclonal antibody used to specifically detect SEPT4, SEPT5, and SEPT8. The IgG fraction of the anti-SEPT4-SEPT5-, and SEPT8-antibodies was prepared from rabbit serum as previously described (Blaser et al. 2003).
The polyclonal SEPT5 antibody was directed against the recombinant fragment of SEPT5 containing 387 residues (complete reading frame). The recombinant fragment had been cloned in a pET30b vector (65 N-terminal residues) (Zieger et al. 2000). The antibody was generated in rabbit (Blaser et al. 2002). The antibody recognizes a 45-kDa protein. SEPT5 antibody recognizes most probably both SEPT5 isoforms, because the two isoforms differ only concerning 18 N-terminal residues. The SEPT5 antibody did not cross-react with SEPT4 and SEPT8.
The polyclonal SEPT4 antibody was directed against the recombinant fragment of SEPT4 containing 102 N-terminal residues. The recombinant fragment had been cloned in a pET30c vector (65 N-terminal residues) (Zieger et al. 2000). The antibody was raised in rabbit. The antibody recognizes a 55-kDa protein (Blaser et al. 2004). SEPT4 antibody recognizes all known isoforms, because it is raised against 102 N-terminal residues, which are present in all the SEPT4 isoforms. The SEPT4-antibody did not crossreact with SEPT5 and SEPT8.
The polyclonal SEPT8-antibody was directed against the recombinant fragment of SEPT8 containing 105 N-terminal residues. The recombinant fragment had been cloned in a pET30c vector. The antibody was raised in rabbit and recognized a 60-kDa protein (Blaser et al. 2002). SEPT8 antibody recognizes SEPT8_v1 (58 kDa) and SEPT8_v2 (64 kDa). The SEPT8 antibody showed no cross-reactivity against SEPT4 and SEPT5.
The antibodies against SEPT4, SEPT5, and SEPT8 do not cross-react with the other septin proteins.
To test the specificity of the primary antibody, control sections were stained simultaneously, following the same procedure with the exception that the primary antibody was omitted (Figure 1C and Figure 3D). Uveal melanoma tissue served as positive control (Figure 1D). All sections were assessed for localization and intensity of specific immunoreactivity on a semiquantitative scale of + to + + + by two blinded observers (MP, PM). In case of disagreement, consent was achieved by discussion.
For PCNA (Dako; Glostrup, Denmark), the optimum concentration of the primary antibody was previously determined to be 1:2000.
For Synaptophysin (Novocastra; Newcastle-upon-Tyne, UK), the optimum concentration of the primary antibody was previously determined to be 1:100.
Results
SEPT4
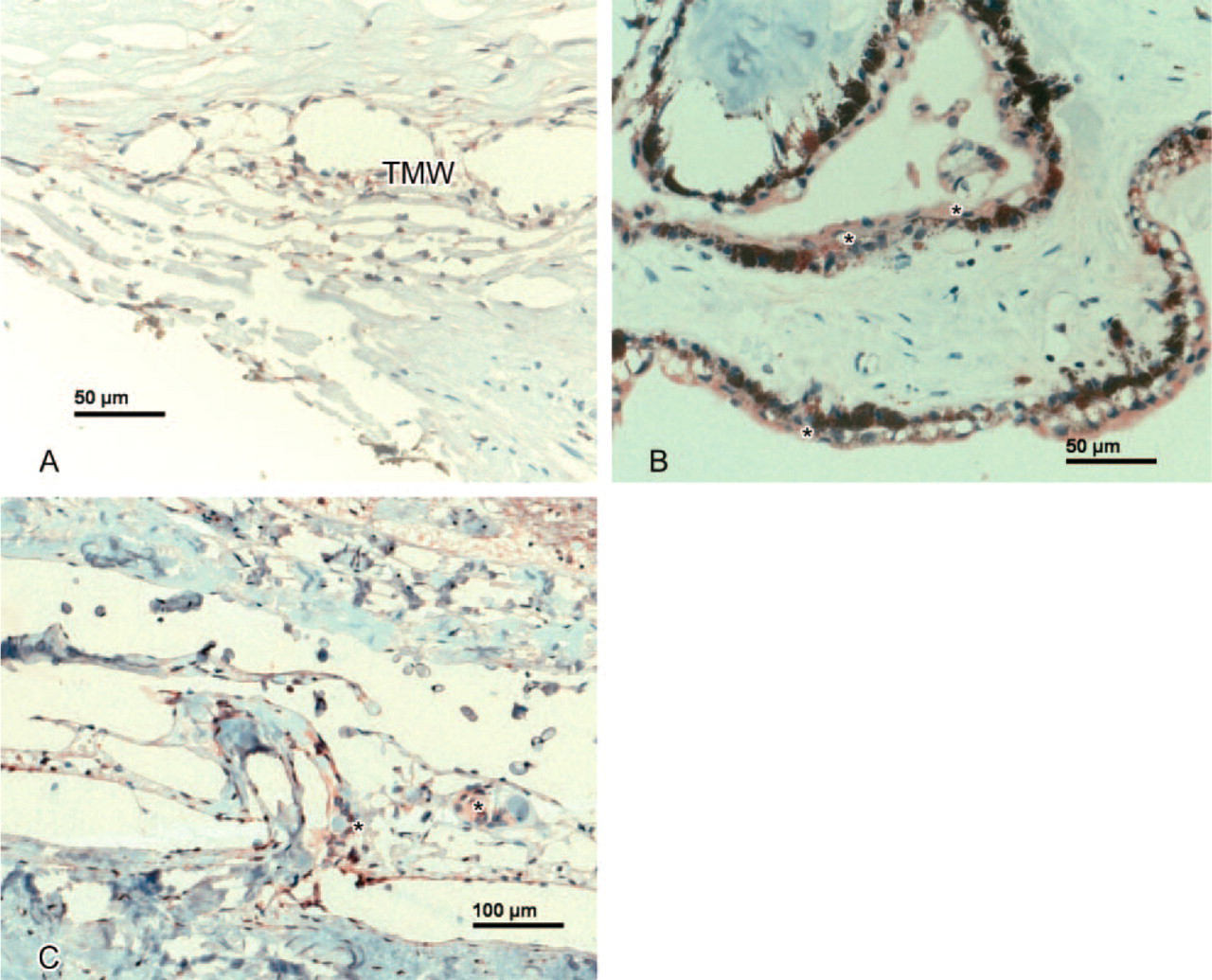
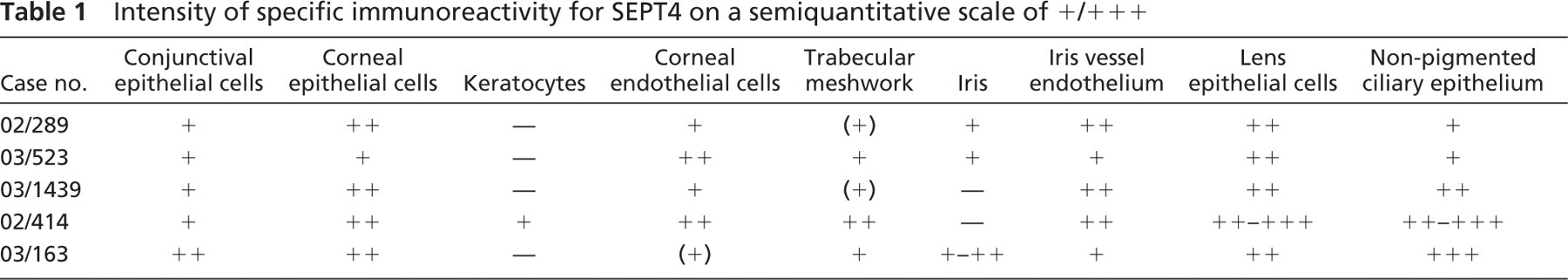
Anterior Segment. Weak expression of SEPT4 was observed in conjunctival epithelial cells. Weak to moderate SEPT4-IR was found in the endothelial cell layer and in the epithelial cell layer of the cornea (Figure 1A). In all but one case, keratocytes lacked SEPT4-IR. The trabecular meshwork showed weak positivity (Figure 2A). The iris tissue revealed weak SEPT4-IR in three of five cases and the iris vessel endothelium weak-moderate positivity in all cases. Lens epithelium showed moderate to strong SEPT4-IR. The nonpigmented ciliary epithelium showed weak to strong SEPT4-IR (Table 1).
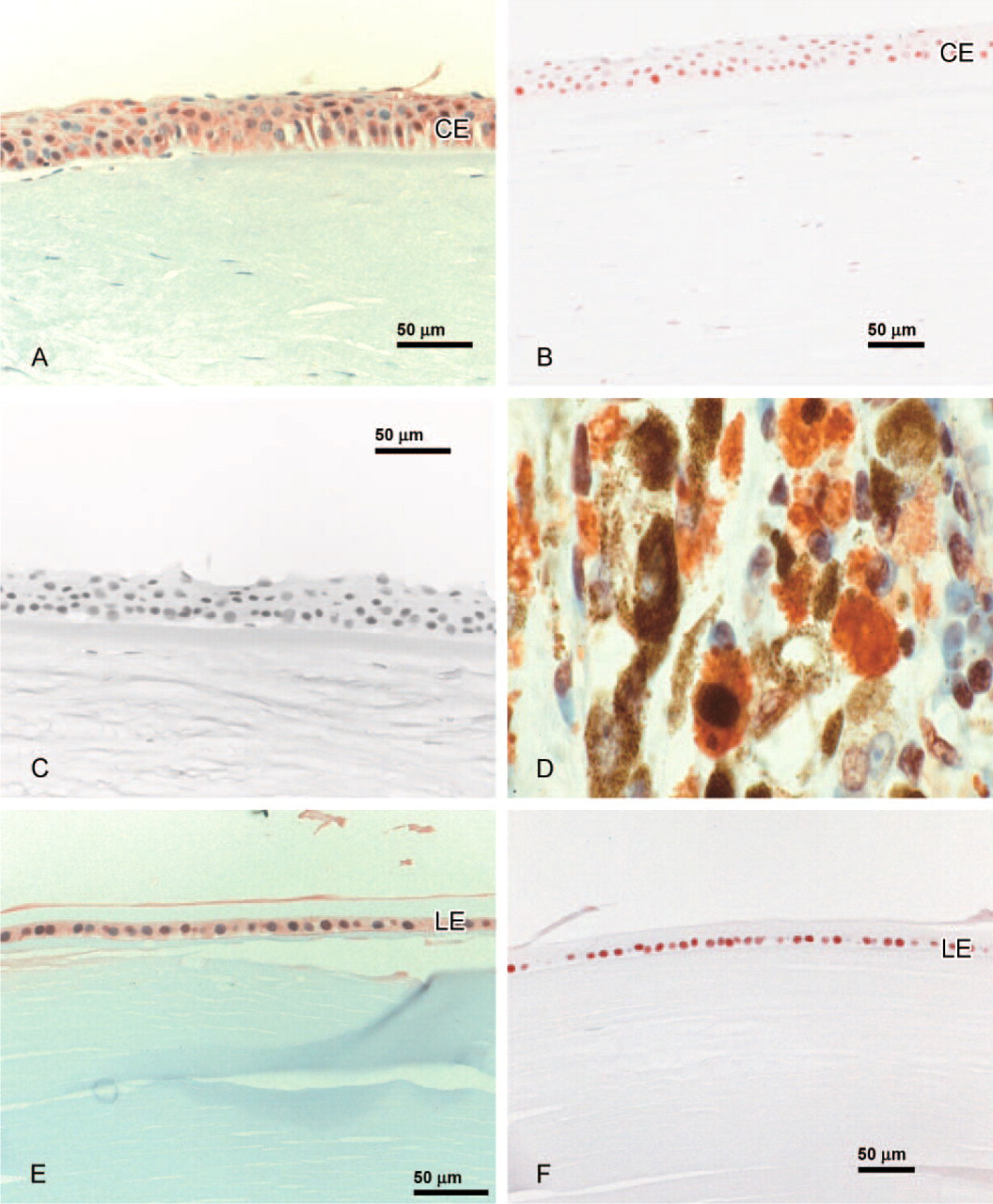
(
(
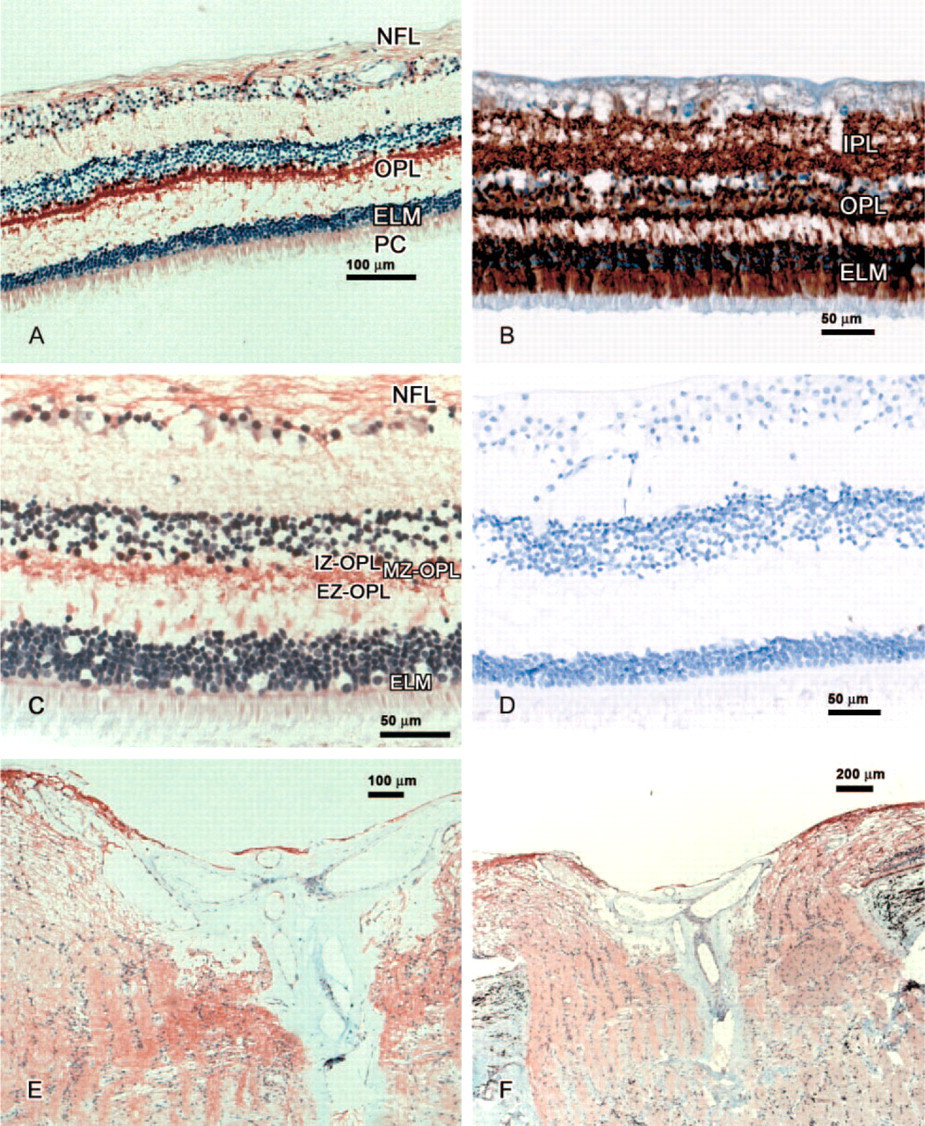
Posterior Segment. In the whole retina, some variation in intensity was observed, but specific SEPT4-IR was seen in the nerve fiber layer (moderate to strong intensity), internal zone of the outer plexiform layer (strong positivity), middle and external zone of the outer plexiform layer (moderate IR), external limiting membrane (mild to moderate IR), and outer segments of the photoreceptor cells (weak to moderate IR). Cell somata and inner segments appeared unlabeled (Figure 3A and Table 2).
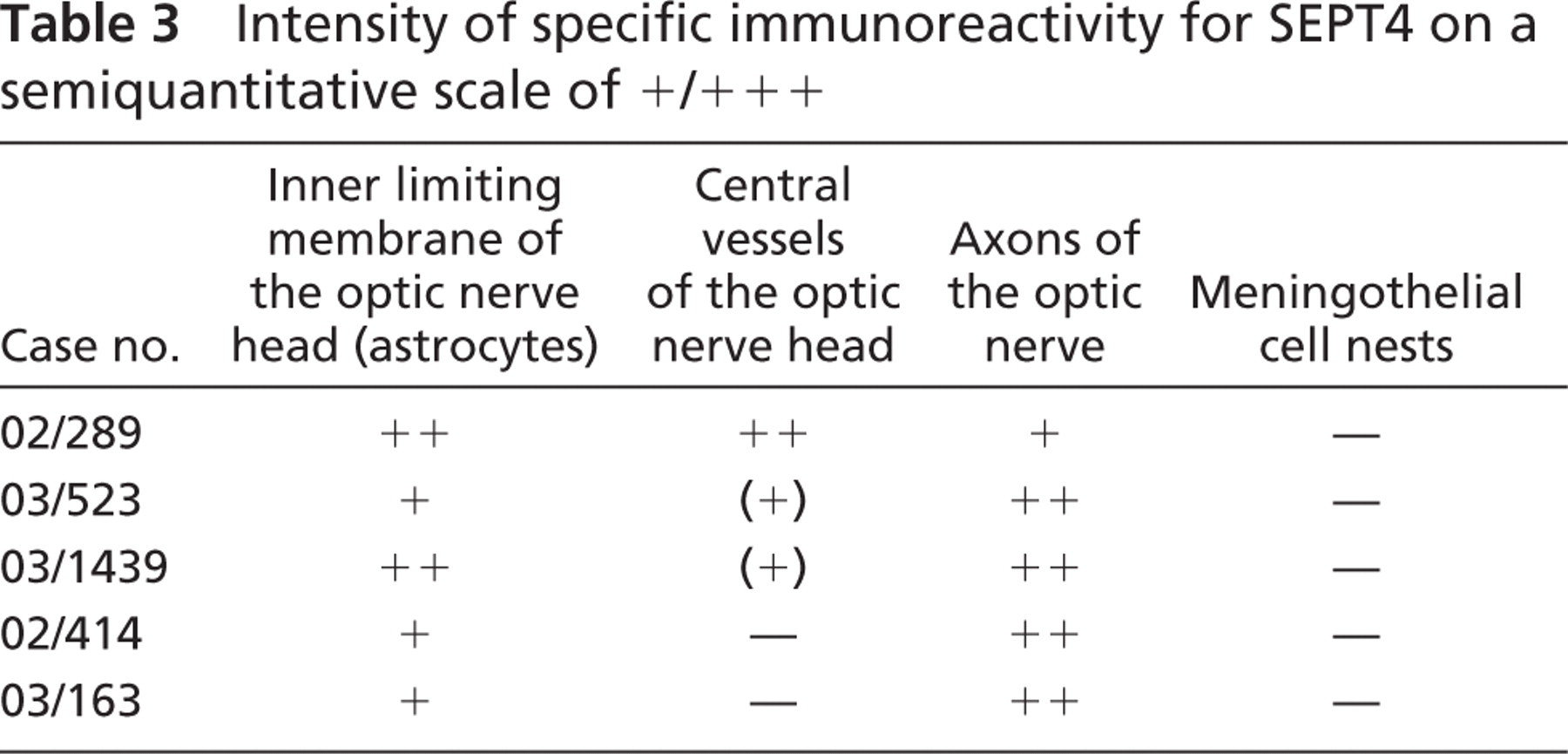
Intensity of specific immunoreactivity for SEPT4 on a semiquantitative scale of +/+ + +
(
Intensity of specific immunoreactivity for SEPT4 on a semiquantitative scale of +/+ + + for the retinal layers and the choroid

Optic Nerve. The inner limiting membrane of the optic nerve head (ONH) as well as the ONH axons showed weak to moderate SEPT4-IR. Some central vessels of the ONH revealed weak SEPT4-IR with no difference in the intensity of staining between arteries and veins (Figure 3E and Table 3).
Anterior Segment. Weak expression of SEPT5 was observed in the corneal endothelial cell layer in three of five cases. The iris tissue showed weak SEPT5-IR in all but one case. Lens epithelium showed weak SEPT5-IR in two of five cases. The nonpigmented ciliary epithelium showed weak to moderate IR in three of five cases (Figure 2B and Table 4).
Posterior Segment and Optic Nerve. No SEPT5-IR was observed in the posterior segment of the globe and in the optic nerve.
Anterior Segment. Weak to moderate expression of SEPT8 was observed in conjunctival epithelial cells. Corneal epithelial and endothelial cells and the keratocytes also revealed weak to moderate IR. The trabecular meshwork showed weak to moderate positivity. Iris tissue and iris vessel endothelium revealed weak to moderate SEPT8-IR. Lens epithelium showed moderate to strong SEPT8-IR (Figure 1E). The nonpigmented ciliary epithelium showed weak to strong IR (Table 5).
Intensity of specific immunoreactivity for SEPT4 on a semiquantitative scale of +/+ + +
Posterior Segment. In the whole retina, some variation of intensity was observed, but specific SEPT8-IR was seen in the nerve fiber layer (strong intensity in four cases), internal zone of the outer plexiform layer (strong positivity), and middle and external zone of the outer plexiform layer (moderate IR). Differing from SEPT4-IR, the outer segments of the photoreceptor cells showed only weak SEPT8-IR in 50% of the cases. Cell somata and inner segments appeared unlabeled. Weak to moderate staining of the retinal vessel endothelium and weak staining of the choroidal endothelium was seen (Figure 3C and Table 6).
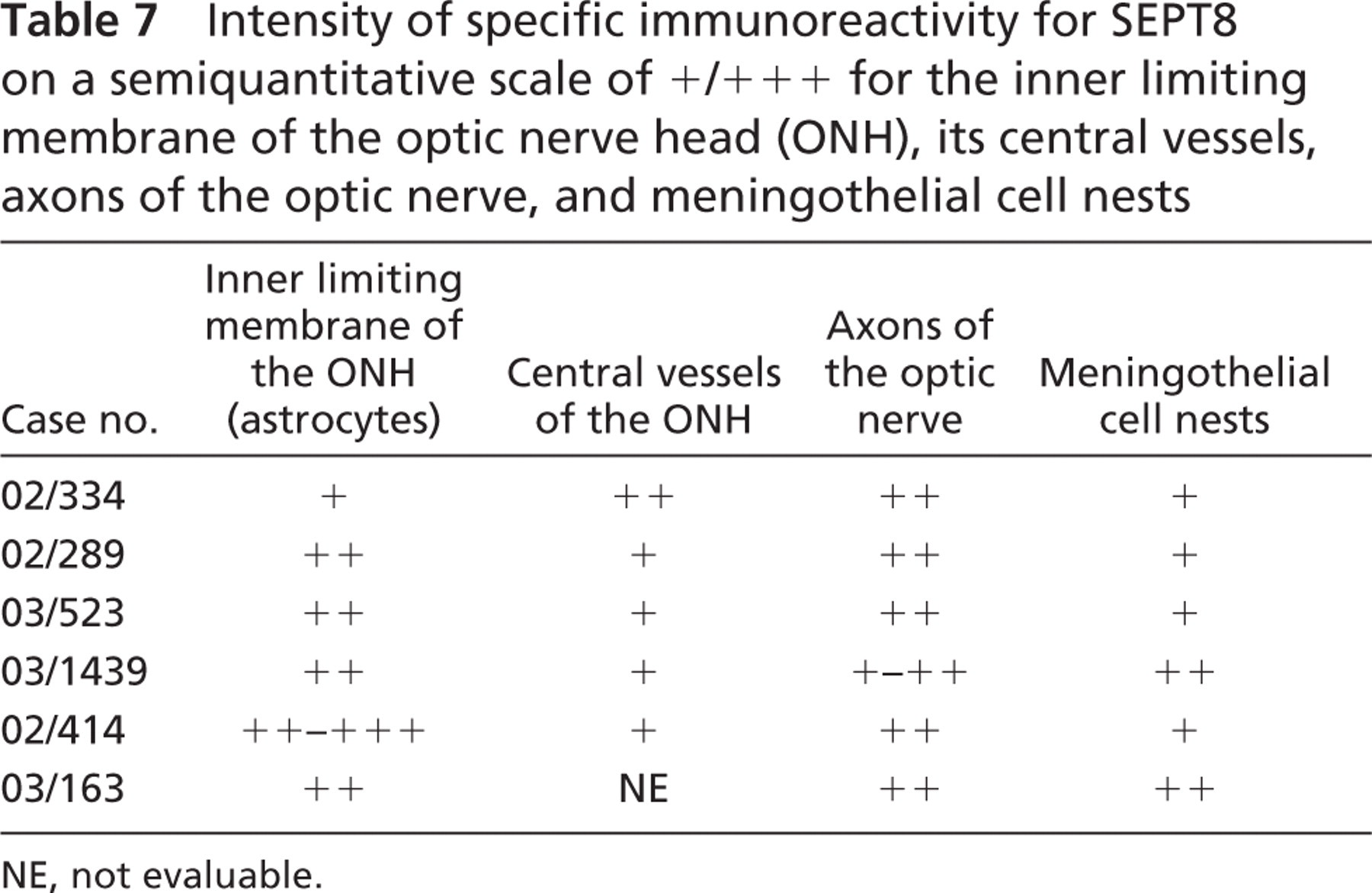
Optic Nerve. The inner limiting membrane of the ONH as well as the ONH axons showed moderate SEPT8-IR. The central vessels of the ONH revealed weak SEPT8-IR with no difference in the intensity of staining between arteries and veins (Figure 3F). Moreover, the meningothelial cell nests of the optic nerve showed weak to moderate SEPT8-IR (Figure 2C and Table 7).
SEPT4, SEPT5, and SEPT8 expression was localized in the cytoplasm.
PCNA. In all cases, corneal (Figure 1B) and lens epithelial cells (Figure 1F) and keratocytes showed strong nuclear PCNA-IR.
Synaptophysin. In all cases, strong synaptophysin-IR was seen in the inner plexiform layer, in the internal zone of the outer plexiform layer, and in the external limiting membrane of the retina (Figure 3B).
Discussion
We examined for the first time the patterns of expression of several septin family members, namely SEPT4, SEPT5, and SEPT8, in the human eye. We detected SEPT4-IR primarily in the epithelium of cornea, lens, and nonpigmented ciliary body epithelium; in the endothelium of cornea and vessels of iris and retina; and in the retinal nerve fiber layer, the outer plexiform layer, the outer segments of the photoreceptor cells, the inner limiting membrane of the optic nerve head, and optic nerve axons. SEPT5-IR was present in corneal endothelial cells, iris tissue, nonpigmented ciliary epithelium, and epithelial cells of the lens. SEPT8-IR almost paralleled that of SEPT4 except for a lower SEPT8-IR of the outer photoreceptor segments and a positive staining of the meningothelial cell nests in the subarachnoidal space of the bulbar segment of the orbital optic nerve.
Intensity of specific immunoreactivity for SEPT5 on a semiquantitative scale of +/+ + +

It is striking that SEPT5 was completely absent in the posterior segment of the globe, where SEPT4 and SEPT8 showed a positive immunostaining. Diversity and redundancy of the mammalian septin system might help to explain this finding. For example, loss of SEPT5 affects exocytosis in mouse platelets (Dent et al. 2002), but is tolerated in the brain, most probably because of compensatory upregulation of other septins (Peng et al. 2002).
The staining of corneal epithelial cells (SEPT8, SEPT4) and keratocytes (SEPT8) might suggest a role for septins in the mitotic rhythm of the cornea. Given the potential role of septins in cell mitosis, it was especially interesting that the staining of SEPT4 and SEPT8 in the epithelium of the cornea was paralleled by a positive staining for the cell cycle-related protein PCNA. Also, the keratocytes were positive for both SEPT8 and PCNA. The lens epithelium revealed moderate to strong SEPT4-IR and SEPT8-IR and also weak SEPT5-IR, which was again paralleled by positive PCNA-IR.
Trabecular meshwork and nonpigmented ciliary epithelium showed positivity for SEPT4 and SEPT8, with the highest IR for SEPT8. This might not only indicate the involvement of septins in the mitotic activity of the ciliary epithelium, but also allows speculations about a potential role for septins in aqueous humor production. The endothelial cells of the trabecular sheets contain a huge number of cytoplasmic organelles responsible for protein synthesis and secretion, and an involvement of SEPT4 and SEPT8 in these processes appears to be likely. SEPT4 and SEPT8 could also be involved in the cell turnover of the nonpigmented ciliary epithelium. Moreover, septins have been found to play an integral part in the compartmentalization of the plasma membrane into specialized regions (Barral et al. 1999). This is an important point in cell polarity, especially in the ciliary epithelium where metabolically dependent active transport of ions is essential for aqueous humor production (Bartels 1989).
Various structures of the retina with all its synaptic and exocytic activity showed a positive SEPT4-IR and SEPT8-IR. It was especially impressive that all photoreceptor cells revealed distinct SEPT4-IR in their outer segments. Each outer segment consists of piles of 600 to 1000 rhodopsin-bearing discs, which are continuously produced and shed. The discs, which are not connected to the cell membrane, are composed of two unit membranes. Because septins are associated with active membrane processes, it can be assumed that SEPT4 is crucially involved in disc formation and shedding of the outer photoreceptor segments.
It was also striking that SEPT4-IR and SEPT8-IR extended as an immunoreactive band throughout the internal zone of the outer plexiform layer. The internal or synaptic zone is almost exclusively composed of the arborizing processes of the bipolar and horizontal cells, and the multiple processes of the Müller cells. The dendrites of both bipolar and horizontal cells synapse with the axons of the rods and cones; interconnections between rod spherules and cone pedicles also are present in this zone. A growing number of septins has been identified as elements of the exocytic machinery in neurons, suggesting that they regulate the release of neurotransmitters (Huizinga et al. 1997; Beites et al. 1999). The high intensity of SEPT4-IR and SEPT8-IR found here supports the idea of an active role for the two septins in retinal neurotransmitter release. It is especially interesting in this context that the positivity for SEPT4 and SEPT8 in the internal zone of the outer plexiform layer and in the external limiting membrane was paralleled by a positive staining for the synaptic vesicle protein synaptophysin.
Age, gender, and time between death and autopsy of all donors are given, followed by the intensity of specific immunoreactivity for SEPT8 on a semiquantitative scale of +/+ + +
Intensity of specific immunoreactivity for SEPT8 on a semiquantitative scale of +/+ + + for the retinal layers and the choroid

NE, not evaluable.
The moderate SEPT4-IR and SEPT8-IR of the optic nerve axons may indicate the involvement of septins in axonal transport processes. Focusing on the retrobulbar portion of the optic nerve and its meningeal sheaths, we observed weak to moderate SEPT8-IR of the meningothelial cell nests in the subarachnoidal space. By covering the trabeculae of the arachnoid mater, meningothelial cells form the principal part of the arachnoid. They can proliferate to form cell nests, which usually become larger and hyalinized with age, and may even finally calcify, thereby forming so-called corpora arenacea (also known as Psammoma bodies or brain sand). The role for SEPT8 in the cell biology of these fibroblast-like cells, however, is unclear.
Intensity of specific immunoreactivity for SEPT8 on a semiquantitative scale of +/+ + + for the inner limiting membrane of the optic nerve head (ONH), its central vessels, axons of the optic nerve, and meningothelial cell nests
NE, not evaluable.
SEPT4-IR and SEPT8-IR was also present in the endothelium of iris vessels, retinal arteries and veins, including the central retinal arteries and veins, and in the choroid. Apart from being involved in the endothelial cell turnover, the selective immunostaining of ocular vessel endothelium might imply a role for septins in ocular vessel function.
In our study, the staining intensity was not identical in all cases, and, in some cases, even absent. It is difficult to find a clear explanation for this observation. Either age, autolytic state of the specimen, compensatory upregulation of other septins, and other yet unknown factors (for example, it has been demonstrated that psychotropic drugs can affect septin expression) (Simantov and Peng 2004) might be responsible for these differences.
Taken together, SEPT4, SEPT5, and SEPT8 are expressed in various ocular tissues, each revealing a distinct expression pattern. Both physiological and potential pathophysiological functions of septins in the human eye remain speculative and deserve further investigation.
Footnotes
Acknowledgements
The authors thank Miriam Vonlanten, Rita Epper, and Susanne Grieshaber for their technical assistance.