
Abstract
Nerve growth factor (NGF) promotes proliferation via its high affinity receptor (TrkA). Its precursor proNGF promotes apoptosis via the pan-neurotrophin-receptor p75. Recently, we have identified NGF and p75 as important hair growth terminators. However, if proNGF is involved or if NGF can also promote hair growth via TrkA is unclear. By RT-PCR we found that NGF/proNGF mRNA levels peak during early anagen in murine back skin, whereas NGF/proNGF protein levels peak during catagen, indicating high turnover in early anagen and protein accumulation in catagen. By immunohistochemistry, NGF and TrkA are found in the proliferating compartments of the epidermis and hair follicle throughout the cycle. In contrast, strong proNGF is found in the highly differentiated inner root sheath and adjacent to the p75+ regressing epithelial strand in catagen. Commercial 7S NGF, which contains both NGF and proNGF, promotes anagen development in organ-cultured early anagen mouse skin, whereas it promotes catagen development in late anagen skin. Together, our findings suggest an anagen-promoting or anagen-supporting role for NGF/TrkA, and a catagen-promoting role for proNGF/p75 interactions. This has important implications for the future design of specific neurotrophin receptor ligands as novel pharmaceuticals in the modification of tissue remodeling processes such as hair growth or wound healing.
S
In epithelial tissue homeostasis, NGF has been shown to promote proliferation of murine and human keratinocytes via its high affinity receptor tyrosine kinase A (TrkA) (Paus et al. 1994; Pincelli et al. 1994; Pincelli and Marconi 2000) and, because keratinocyte proliferation is essential to hair growth (Stenn and Paus 2001), one might expect that NGF also acts as a hair growth promoter—for example to hair follicles in the well-known C57BL/6 mouse hair cycle model (Paus et al. 1994; Müller-Röver et al. 2001; Stenn and Paus 2001) similar to other keratinocyte growth factors (Danilenko et al. 1995). However, despite the extensive literature published on neurotrophin hair growth modulatory effects (see the following section for details), NGF as a hair growth promoter has not been studied.
Interestingly, proliferation under NGF treatment in murine epidermal and hair follicle keratinocytes occurs only in skin with all hair follicles in the resting stage of the hair cycle (telogen) (Paus et al. 1994). During the late growth phase (high anagen), NGF inhibits proliferation of keratinocytes (Paus et al. 1994). This may be explained by binding of NGF to the panneurotrophin-receptor p75, a member of the tumor necrosis factor receptor family, which commonly induces apoptosis and is a potent player in hair follicle regression (catagen) (Botchkarev et al. 2000). However, NGF binds to p75 only with low affinity and, if TrkA is available, NGF will not transmit a death signal and TrkA signaling is amplified by p75 (Pincelli and Marconi 2000; Peters et al. 2003).
Very recent and intriguing data have now shown that the NGF precursor, proNGF, which was long thought to serve no biological activity, is the long searched for high-affinity ligand of p75 and that proNGF/p75 interaction induces apoptosis in neuronal cells (Lee et al. 2001; Chao and Bothwell 2002; Ibanez 2002 671). Thus, we hypothesize that proNGF/p75 interaction may act as a potent catagen inducer, for example, in the murine hair follicle.
In this context, it is important to note that all commercially available NGF protein preparations tested for the presence of NGF and proNGF do contain both compounds whether they are purified from mouse submaxillary gland or recombinant (Reinshagen et al. 1997, 2000). Thus, in vivo and in vitro experiments employing NGF should be considered as experiments employing an NGF/proNGF cocktail and interpretation of the respective biological responses demands knowledge on receptor expression in the treated tissues and cells.
However, the shared precursor mRNA and the heretofore lack of commercially available antibodies to proNGF did not allow to differentiate between NGF and proNGF production and expression, and there are very few studies published to date that consider proNGF as an important additional player in their equations.
With the study presented here, we aim to fill the gap in the analysis of neurotrophin skin homeostasis using the highly instructive C57BL/6 mouse model for hair cycle-associated cyclic skin reorganization. In this model, we determine NGF, proNGF, TrkA, and p75 expression throughout the hair cycle to learn from expression patterns about possible functional interactions. We employ semiquantitative TaqMan PCR, ELISA technique, and immunohistochemistry. Functional relevance of the respective findings is then tested by the analysis of commercial 7S NGF as a hair growth modulatory agent in organ-cultured murine hair follicles.
Materials and Methods
Mice
Six to 9-week-old, syngenic, female C57BL/6 mice in the telogen stage of the hair cycle were purchased (Charles River; Sulzfeld, Germany). The mice were housed in community cages under 12-hr light periods at the animal facilities of the Center for Biomedical Research, Charité, University-Medicine Berlin, Berlin, Germany. Animal care and experimental procedures were followed according to institutional guidelines and conformed to the requirements of the state authority for animal research conduct (LaGetSi, Berlin Germany).
Hair Cycle Induction
Anagen was induced in the back skin of mice by depilation as described (Paus et al. 1990). This procedure induces the highly synchronized development of anagen follicles that are morphologically indistinguishable from spontaneous anagen follicles (Müller-Röver et al. 2001).
Real-Time RT-PCR for NGF
Real-time RT-PCR was employed to obtain quantitative data on differences between NGF mRNA expression throughout the hair cycle. This assay exploits the 5’ nuclease activity of AmpliTaq Platin (Invitrogen; Karlsruhe, Germany) DNA polymerase to cleave a fluorogenic probe designed for NGF (TipMolBiol; Berlin, Germany) and, to normalize our samples, a fluorogenic probe for the housekeeping gene hypoxanthine phosphoribosyl transferase (HPRT) was used in real-time RT-PCR. The sets of primers and probes were designed as follows (Peters et al. 2004). NGF probe: sequence 6FAM-gAT Cgg CgT ACA ggC AgA ACC gTA CAC AgA T XT A-PH; reverse primer 5′: Tag TCC AgT ggg CTT CAg ggA; forward primer 5′: TTT gCC AAg gAC gCA gCT T); HPRT probe: sequence 6FAM-TTg CAg ATT CAA CTT gCg CTC A XT CTT-PH; reverse primer 5′: CAC Agg ACT AgA ACA CCT gC; forward primer 5′: gTT ggA TAC Agg CCA gAC TTT gT.
The real-time RT-PCR reactions were developed as previously published (Knackstedt et al. 2003), except that we used 0.2 μl probe at 20 μM. Each analysis was normalized to HPRT by calculating the difference between the cycle time (cT) for HPRT and the cT for NGF as ΔCT = CT HPRT – CT NGF. Amount mRNA was calculated using the formula (1/2ΔCT) X 1000 as described before (Knackstedt et al. 2003).
NGF-ELISA
Approximately 1 mg deep frozen skin was crushed in liquid nitrogen and 1 ml lysing buffer (0.025% NP-40, 0.01% NaN3, 12.5 mM EDTA in PBS) was added. The samples were than exposed to ultrasound for 30 sec, incubated for 20 min at 37C, and centrifuged. After centrifugation, protein content was determined by BCA protein assay according to the manufacturer's instructions (Pierce; Rockford, IL). NGF content was determined by NGF Emax Immunoassay System according to the manufacturer's instructions (Promega; Madison, WI). Last, the NGF content was calculated for 100 μg total protein.
Immunohistochemistry
Skin was cryopreserved as described at defined days after de pilation (unmanipulated telogen skin = day 0, anagen I = 5 day 3, anagen IV = day 5, anagen VI = day 12, catagen = 19; N per day/stage analyzed = 5) (Müller-Röver et al. 2001). For simplicity, skin with all follicles in telogen is referred to as “telogen skin” and the other stages as “anagen/catagen skin.”
Immunofluorescence histochemistry was performed following established protocols (Botchkarev et al. 1997, 2000; Peters et al. 2002, 2004). Briefly, cryostat sections (10 μm) were incubated overnight at room temperature in a humidity chamber with the primary antisera to NGF (1:50; Santa Cruz Biotechnology, Santa Cruz, CA; rabbit affinity purified polyclonal antibody raised against the N terminus of mouse NGF), TrkA (1:50; Santa Cruz; rabbit affinity purified polyclonal antibody raised against a peptide near the C terminus of human TrkA, recommended for mouse TrkA detection), proNGF (1:150; Chemicon, Temecula, CA; rabbit affinity purified polyclonal antibody raised against a synthetic peptide spanning amino acids 84–104 of rat proNGF, which is 100% homologous with the mouse sequence and not present in mature NGF), or p75 (1:100, Chemicon; rat affinity purified monoclonal antibody raised against murine p75) as published previously (Botchkarev et al. 2000; Peters et al. 2002, 2004; Tometten et al. 2004). This was followed by an incubation of 1 hr at 37C with tetramethylrhodamine-isothiocyanate (TRITC) or fluorescein-isothiocyanate (FITC)-conjugated F(ab)2 fragments of goat anti-rabbit or goat anti-rat IgG (Dianova; Hamburg, Germany) at a dilution of 1:200. Washing steps with TBS were interspersed between all staining steps. All antibodies were diluted in TBS at pH 7.4 containing 2% normal goat serum. Cell nuclei were counterstained with DAPI (Mecklenburg et al. 2000).
For double-immunofluorescence detection of TrkA-positive cells and Ki67-IR, the protocol for the TrkA staining was combined with the manufacturer's protocol for Ki67-immunohistochemistry (Magerl et al. 2001). TrkA was detected by TRITC and Ki67-IR by FITC and the sections were counterstained by DAPI.
The presented staining patterns have been reproduced at least five times per staining protocol. In addition, four types of negative controls were run to confirm specificity: (1) slides were incubated with the secondary antibody alone; (2) slides were incubated with the primary antibody after preincubation with control peptides for the specific anti-sera (Sigma; St Louis, MO; 10–20 μg/ml, 37C, 1 h); (3) slides were incubated with rabbit/rat IgG as primary antibody; and (4) stainings were performed on skin obtained from p75 and TrkA knockout mice as described previously (Botchkarev et al. 2000). All of these negative controls resulted in significant reduction (dose dependently for control peptides) or complete disappearance (after obliteration or replacement of primary antibody) of the observed specific immunoreactivity patterns. As positive controls, whole mount mouse fetuses were used. We found differential expression of all investigated antigens in the brain and lung of these embryos, as had been published previously (Braun et al. 1998; Botchkarev et al. 2000). In addition, extrafollicular compartments of adult skin, known to contain p75 (nerve fibers), NGF (mast cells), and TrkA (basal epidermal keratinocytes) served as intrinsic positive controls. Specificity of antibody labeling was also confirmed by use of alternative primary antibodies obtained from alternative companies or private donations and raised against different epitopes then the antibodies obtained from Santa Cruz. For TrkA and p75, this has been published previously (Botchkarev et al. 2000). NGF staining patterns were confirmed by application of an alternative polyclonal NGF-antiserum generated in rat (1:100; R & D Systems, Minneapolis, MN). In the case of proNGF, no alternative antibody was commercially available. These very extensive positive and negative controls confirmed the specificity and sensitivity of the employed immunohistological technique. The use of antibody abbreviations in expressions like “NGF-immunoreactive” or “NGF+” implies labeling with a specific antibody in recognition that an antibody could possibly crossreact with some other antigen.
Histomorphometry
Sections were examined at 400X magnification under a Zeiss Axioscope fluorescence microscope (Zeiss; Jena, Germany). The expression patterns of NGF, proNGF, and TrkA were scrutinized in defined cutaneous and follicular compartments such as the dermis, the subcutis, the arrector pili muscle, blood vessels, the dermal papilla, the inner root sheath, the outer root sheath, the epithelial strand, the hair matrix, the isthmus and bulge region, the sebaceous gland, mast cells, and nerve fibers. For each stage of hair follicle cycling, at least 10 microscopic fields each of three to five different mice, containing a minimum of 30 different hair follicles per mouse, were studied and the immunoreactivity patterns were recorded qualitatively in computer-generated recording schemata. Photo documentation was carefully performed by a digital image analysis system (Visitron Systems; Puchheim, Germany; Spot advanced software, version 3.5.2), adjusting exposure of each individual image to capture the brightest sections still with a point-by-point differentiation between color and brightness values. For optimal reproduction, brightness and contrast of the images were later improved digitally.
Neurotrophin Administration in Early and Late Anagen Skin Organ Culture
Four-mm, full-thickness skin punch biopsies were prepared under sterile conditions from C57BL/6 mouse back skin, containing a homogeneous population of early anagen hair follicles (day 3 after depilation, containing telogen and anagen I–II hair follicles) or late anagen hair follicles (day 16 after depilation, containing anagen VI hair follicles) (Chase 1954; Müller-Röver et al. 2001), following previously described skin organ culture protocols (“histoculture,” Li et al. 1992; Botchkarev et al. 1999b; Peters et al. 2001). Twelve randomized skin fragments from two different mice were placed dermis down on prehydrated gelatin sponges (six per sponge and well; Gelfoam, Upjohn Co., Kalamazoo, MI) in six-well plates, containing 5 ml DMEM, 10% FBS, 50 μg/ml
Hair Cycle Staging and Statistical Analysis
For the evaluation of the pharmacological experiments, histomorphometric hair cycle staging was performed on 10 μm cryosections as previously described (Maurer et al. 1997; Botchkarev et al. 1999b; Müller-Röver et al. 2001). Briefly, for the precise identification of the defined stages of hair follicle cycling, histochemical detection of endogenous alkaline phosphatase activity was used, because this visualizes the morphology of the dermal papilla as a useful morphological marker for hair follicle staging (Handjiski et al. 1994; Müller-Röver et al. 2001). The hair cycle score and percentage of hair follicles in defined anagen stages was calculated per group as described (Chase 1954; Müller-Röver et al. 2001).
The data were pooled from three to five different mice per investigated time point or group, and the means + SEM calculated. p Values were determined by Mann-Whitney U test for non-parametric samples.
Results
NGF/proNGF mRNA Expression Shows Distinct Hair Cycle–dependent Expression Patterns
On the message level, NGF and proNGF cannot be distinguished because they are both encoded by the same mRNA. The NGF/proNGF mRNA levels peak in very early anagen on day 1 after anagen induction by depilation after hair growth is initiated (Figure 1). Toward day 3, when maximal proliferative activity is present in the hair follicle epithelium, NGF/proNGF mRNA levels fall again, and stay low throughout the rest of the anagen development to reach levels significantly below the telogen level on day 12 after depilation, when all hair follicles have reached the final stage of anagen development, anagen VI (Figure 1). Compared with anagen skin, NGF/proNGF mRNA expression increases substantially in catagen skin to reach almost telogen levels on day 19 after depilation, when most hair follicles have reached the final catagen stages (Figure 1).
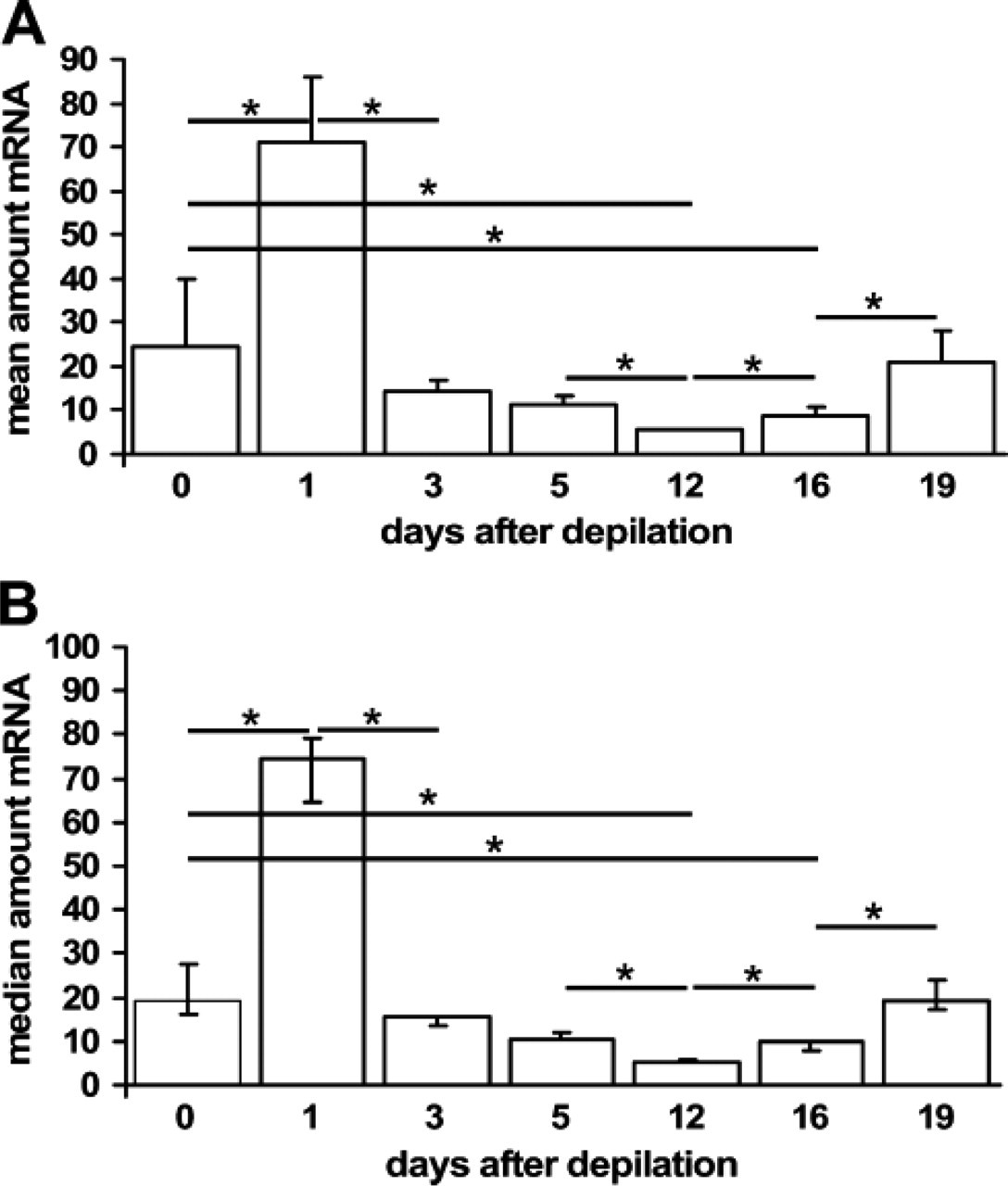
Nerve growth factor (NGF)/proNGF mRNA levels throughout the murine hair cycle. The figure shows NGF/proNGF mRNA amounts in arbitrary units as detected in three different mice per time point. Quantitative TaqMan PCR was performed on whole skin extracts derived from back skin with all their hair follicles in telogen (day 0, unmanipulated skin), anagen I–II (day 1 after anagen induction by depilation), anagen III [day 3 post depilation (p.d.)], anagen V (day 5 p.d.), anagen VI (day 12 p.d.), catagen I–III (day 16 p.d., note that anagen-catagen transition occurs spontaneously around day 16 p.d.), or catagen V–VIII (day 19 p.d.). NGF mRNA level was determined in triplicate for each mouse skin extract. After confirmation of significant differences between groups by performing a Kruskal-Wallis Test, p values were determined by Mann-Whitney U: ∗ < 0.05. p values were adjusted to preclude the possibility of type 1 errors as is common practice with statistical analysis using SPSS (SPSS Inc.; Chicago, IL). Data are shown as mean with standard error (
NGF/proNGF Protein Expression is Highest During Anagen-Catagen Transition
To determine the NGF protein content in murine back skin, we employed the ELISA technique on full-thickness skin biopsies. This technique does not distinguish between NGF and proNGF because the employed antibody detects both. In telogen skin, the NGF/proNGF protein concentration is in between anagen and catagen levels (Figure 2), and throughout anagen development, the concentrations vary greatly. However, the lowest NGF/proNGF protein level is detectable in mid-anagen, on day 5 after induction of anagen. This concentration differs significantly from telogen and catagen skin (Figure 2). During catagen development, NGF/proNGF concentration peaks in late catagen with the highest detectable NGF/proNGF concentration as compared with all other hair cycle stages (Figure 2).
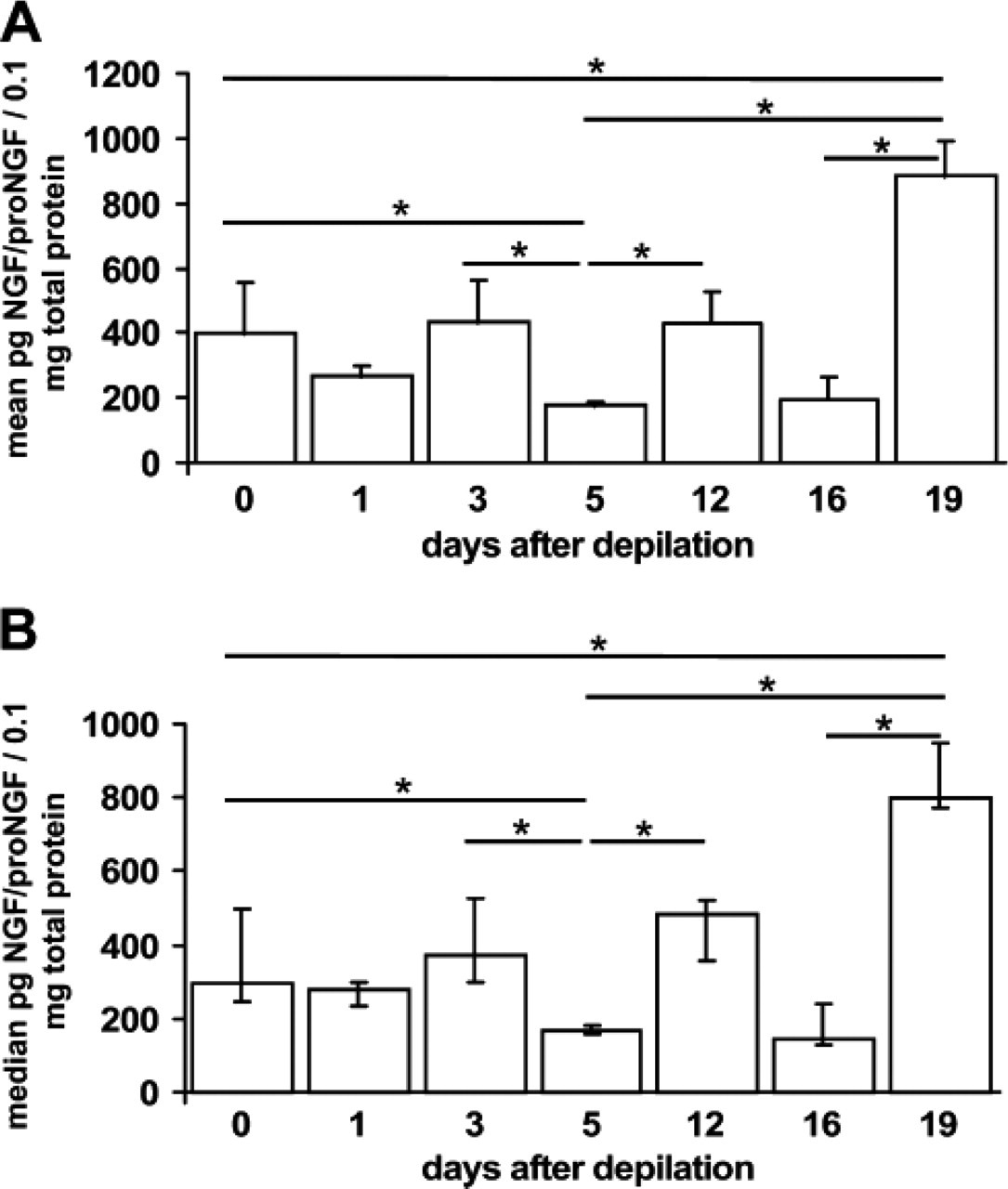
Nerve growth factor (NGF)/proNGF protein levels throughout the murine hair cycle. The figure shows NGF/proNGF protein content per 0.1 mg total protein as detected in three different mice per time point. Protein was extracted from full thickness skin biopsies derived from back skin with all their hair follicles in telogen (day 0, unmanipulated skin), anagen I–II (day 1 after anagen induction by depilation, p.d.), anagen III (day 3 p.d.), anagen V (day 5 p.d.), anagen VI (day 12 p.d.), catagen I–III (day 16 p.d., note that anagen-catagen transition occurs spontaneously around day 16 p.d.), or catagen V–VIII (day 19 p.d.). After confirmation of significant differences between groups by performing a Kruskal-Wallis Test, p values were determined by Mann-Whitney U: ∗ < 0.05. p values were adjusted to preclude the possibility of type 1 errors as is common practice with statistical analysis utilizing SPSS. Data are shown as mean with standard error (
NGF and proNGF Show Distinct Immunoreactivity Patterns in the Hair Follicle Epithelium Throughout the Hair Cycle and Vary in their Receptor Colocalization
Throughout the hair cycle, immunoreactivity to NGF, TrkA, proNGF, and p75 are detected in the nonremodeled skin compartments. NGF, TrkA, and p75 are detected in the basal, proliferating layer of the epidermis, whereas p75 is also found in the suprabasal, terminally differentiating epidermal layers (Figure 3, Figure 4, and Figure 6) and proNGF stains the arrector pili muscle (Figure 3, Figure 4, and Figure 6). In the hair follicle compartments that are remodeled during hair cycling, however, expression varies hair cycle dependently as described in the following section.
In telogen skin, expression of NGF, TrkA (Figures 3A and 3B), and p75 (Figures 3E and 3F) is found the outer most layer of the distal hair follicle epithelium, whereas the lower hair follicle, including the bulge region, is only reactive to TrkA antibody and the sebaceous gland only to NGF antibody (Figures 3A and 3B). By contrast, proNGF does not stain the hair follicle epithelium of telogen hair follicles (Figures 3C and 3D). None of the four markers can be detected in the dermal papilla of telogen hair follicles (Figure 3).
The staining patterns for NGF and TrkA are largely the same in the anagen hair follicle (Figures 4A–4E) when compared with telogen expression. However, the positive double staining in the basal outer root sheath is now extending deep into the subcutis because of the increased length of the growing anagen hair follicle as compared with the telogen hair follicle. TrkA is also present in the hair bulb and matrix (Figures 4D and 4E) where it colocalizes with a marker for proliferation (Ki67; Figure 5). Only weak TrkA outer root sheath expression is present in the isthmus and bulge region, which shows single cells with strong TrkA immunoreactivity (Figures 4A and 4B). These cells are located where keratinocyte and melanocyte stem cells reside in the hair follicle epithelium (Nishimura et al. 2002) and posses tiny dendrites. In contrast, proNGF staining is prominent in the terminally differentiating inner root sheath of anagen hair follicles (Figures 4F–4I), where mostly p75 expression is also found (Figures 4J–4N). In addition, p75 expression is found in the proximal outer root sheath (Figures 4M and 4N), whereas proNGF localizes to the connective tissue sheath adjacent to the proximal hair bulb (Figure 4I). Interestingly, p75 expression can be found in the dermal papilla of the few hair follicles that fulfill the morphological criteria of early regressing hair follicles (catagen I–II) as early as day 12 after induction of hair growth by depilation (Figure 4N).
In the catagen hair follicle, NGF/TrkA immunoreactivity is extended to the entire outer root sheath and hair bulb with the exception of the epithelial sack surrounding the club hair (Figures 6A and 6B), which stains strongly with proNGF (Figure 6D). proNGF is also positive in the arrector pili muscle (Figure 6) and in the regressing inner root sheath (Figure 6D). In the compartment where keratinocyte apoptosis is prominent during catagen development, the epithelial strand, and in and around the epithelial sack, strong p75 immunoreactivity is present (Figure 6F) as is in the differentiating distal epidermis (Figure 6E), whereas single cells in epithelial strand are strongly TrkA+ (Figure 6).
NGF/proNGF Promotes Hair Growth in Early but Terminates Hair Growth in Late Anagen Skin Organ Culture
Our early anagen skin organ culture model employs murine skin on day 3 after anagen induction by depilation. On this day, the majority of hair follicles is in stages telogen–anagen II. These hair follicles express NGF, TrkA, and p75 immunoreactivity in their hair follicle epithelium, but not proNGF and display maximal keratinocyte proliferation. In this model, we found an increased hair cycle score and a higher percentage of hair follicles in later hair cycle stages in the biopsies treated with a commercial NGF-protein preparation, which contains both NGF and proNGF, 7S NGF (Sigma) (Reinshagen et al. 1997) (Figures 7A–7E). Thus, in early anagen, NGF/proNGF promotes anagen progression.
In contrast, in organ-cultured late-anagen hair follicles biopsied from skin on day 16 after anagen induction by depilation, the same NGF preparation produces an increased hair cycle score and a higher percentage of catagen hair follicles (Figures 7E–7H). These hair follicles are biopsied just before the spontaneous entry of anagen hair follicles into catagen and by immunohistochemistry express low NGF and TrkA but high p75 and proNGF in their hair bulbs. Thus, in late anagen, NGF/proNGF promotes the anagen-catagen transition.
Discussion
Bearing in mind the earlier published expression patterns of p75 during the anagen-catagen transition (Botchkarev et al. 2000), the distribution patterns of NGF, proNGF, TrkA, and p75, observed here throughout the entire hair cycle, suggest an anagen-promoting or anagen-supporting role for NGF/TrkA signaling, and a catagen-promoting role for proNGF/p75 interactions. We conclude that the differential control of hair follicle keratinocyte apoptosis and proliferation by NGF, proNGF, TrkA, and p75 reflects local changes in NGF expression and NGF processing, and in the availability and coexpression of high- versus low-affinity NGF receptors.
We find a high mRNA expression for proNGF/NGF in early anagen skin indicating the high requirement for this growth factor during this stage of hair growth. Interestingly, at the protein level, proNGF/NGF amounts are not high during early anagen, which suggest a high turnover rate for this growth factor in early anagen skin. However, the enzymes responsible for proNGF to NGF cleavage and further NGF processing (e.g., plasmin, NGFgamma, trypsin, furin, prohormone convertases, some matrix metalloproteinases) (Seidah et al. 1996; Lee et al. 2001) are abundant and redundant and therefore do not facilitate quantification of processing by determination of their levels in our hair cycle model.
Differential expression of mRNA and protein may also reflect translational control during the early stages of hair growth. During these stages, mRNA may be accumulated to be prepared for massive protein synthesis during later hair cycle stages. Also, thickness of the skin increases substantially during mid- to late anagen and than decreases again in late catagen. It is not known, how this affects the relative content of keratinocytes, fibroblasts, or blood vessels in the whole-skin extracts, though all measurements have been adjusted to total mRNA or protein content. Immunohistomorphometry provides some insight into the compartmental protein expression. Future analysis of individual compartments for example by laser microdissection will provide more exact data on compartmental mRNA content.
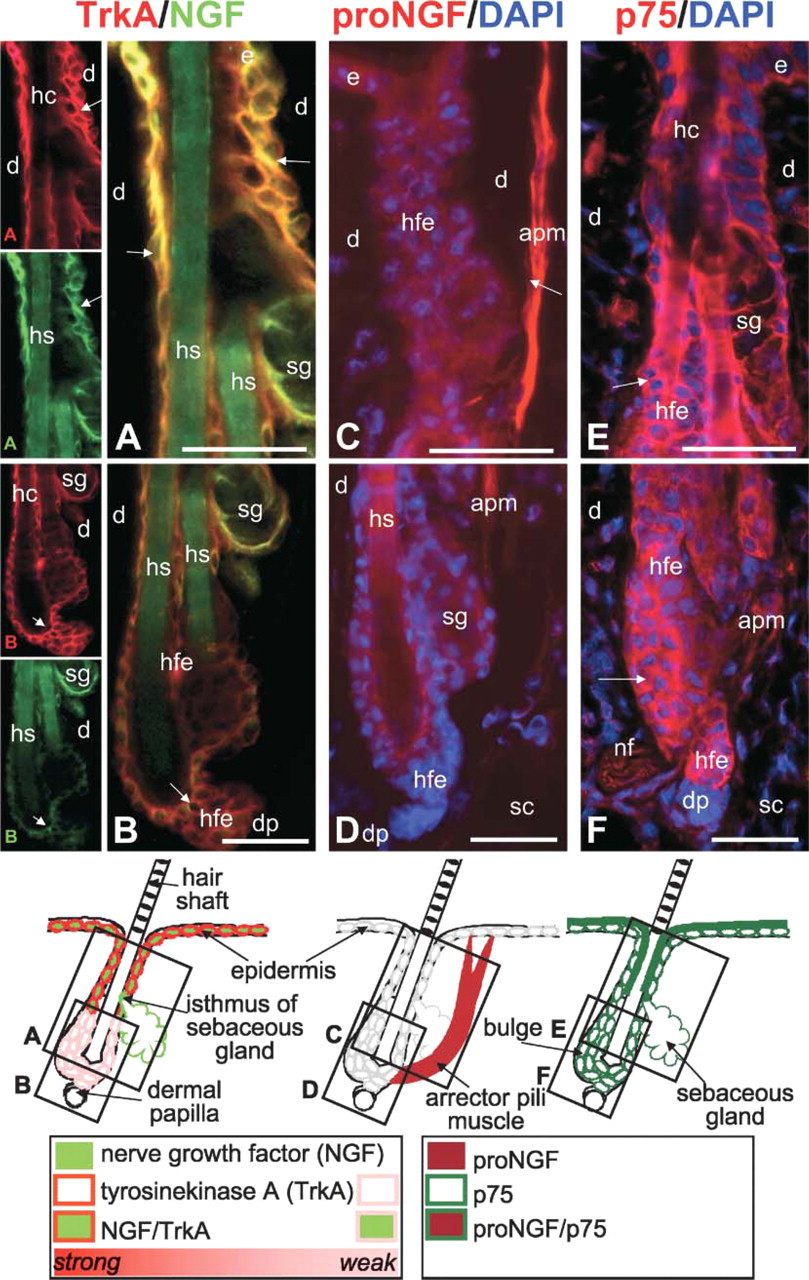
Immunohistochemical localization of nerve growth factor (NGF), TrkA, proNGF, and p75 in the murine telogen hair follicle. Cryo-sections of murine back skin with all hair follicles in the resting phase of the hair cycle (telogen) were immunolabeled with rhodamine (red) to detect TrkA, proNGF, and p75 protein or with fluorescein (green) to detect NGF as described in Materials and Methods. Cell nuclei have been counterstained with DAPI (blue). Schematic drawings indicate the most representative staining pattern as scrutinized from 10 microscopic fields in five different mice per staining. Black boxes in schematic drawings indicate localization and proportion of the corresponding photomicrographs labeled with the identical upper case letter in the lower left corner. Orange staining indicates overlay of NGF and TrkA staining. (
Evidence for high NGF requirement, however, and for the pro-proliferative effect of NGF/TrkA interaction in anagen, is given by their colocalization in epithelial cells bound to continuously proliferate (Müller-Roöver et al. 2001; Stenn and Paus 2001) in the basal layer of the epidermis and the basal distal outer root sheath as well as in the hair bulb, where TrkA colocalizes with the proliferation marker Ki67. In addition, we find TrkA and p75 expression in the proximal hair bulb of telogen and early anagen hair follicles, where proliferation of keratinocytes is switched on and promoted during hair growth (Müller-Röver et al. 2001; Stenn and Paus 2001), whereas proNGF is absent from these compartments. This hypothesis is further supported by our finding that 7S NGF promotes hair growth in early anagen skin organ culture.
NGF promotion of keratinocytes proliferation via TrkA (Paus et al. 1994; Pincelli et al. 1994; Pincelli and Marconi 2000) may involve mast cell differentiation (Welker et al. 1998), degranulation (Paus et al. 1994), and synergism with other growth factors such as insulin-like growth factor (Wilkinson et al. 1994). Moreover, TrkA signaling inhibits keratinocyte apoptosis via upregulation of Bcl-2 in human keratinocytes (Pincelli et al. 1997). Thus, the hair growth–promoting effect of NGF in early anagen occurs most likely via its high affinity-receptor TrkA.
By contrast, p75 expression concentrates in differentiated keratinocytes of the epidermis and hair follicle and in the regressing epithelial strand of catagen hair follicles and proNGF is found in the same or directly adjacent cell populations. This proNGF expression pattern in murine hair follicles corresponds with the only other hair follicle analysis employing proNGF antibodies performed on ovine hair follicles (Yardley et al. 2000). In addition, p75 is found in the dermal papilla before the onset of catagen. Thus, during anagen-catagen transition, the situation changes and proNGF/p75 interaction outplays NGF/TrkA interaction.
These findings are further supported by earlier reports on hair follicle regression (catagen) showing coexpression of p75 with TUNEL in apoptotic “hot spots” (Lindner et al. 1997), which lack coexpression of TrkA (Botchkarev et al. 2000). Neurotrophin catagen induction is absent in p75 null skin or when p75 is antagonized (Lindner et al. 1997; Botchkarev et al. 1999a, 2000, 2003). In our study, catagen is induced in late anagen skin by treatment with 7S NGF containing both NGF and proNGF. Moreover, p75 involvement was also shown in NGF-mediated, stress-triggered hair growth termination (Peters et al. 2004).
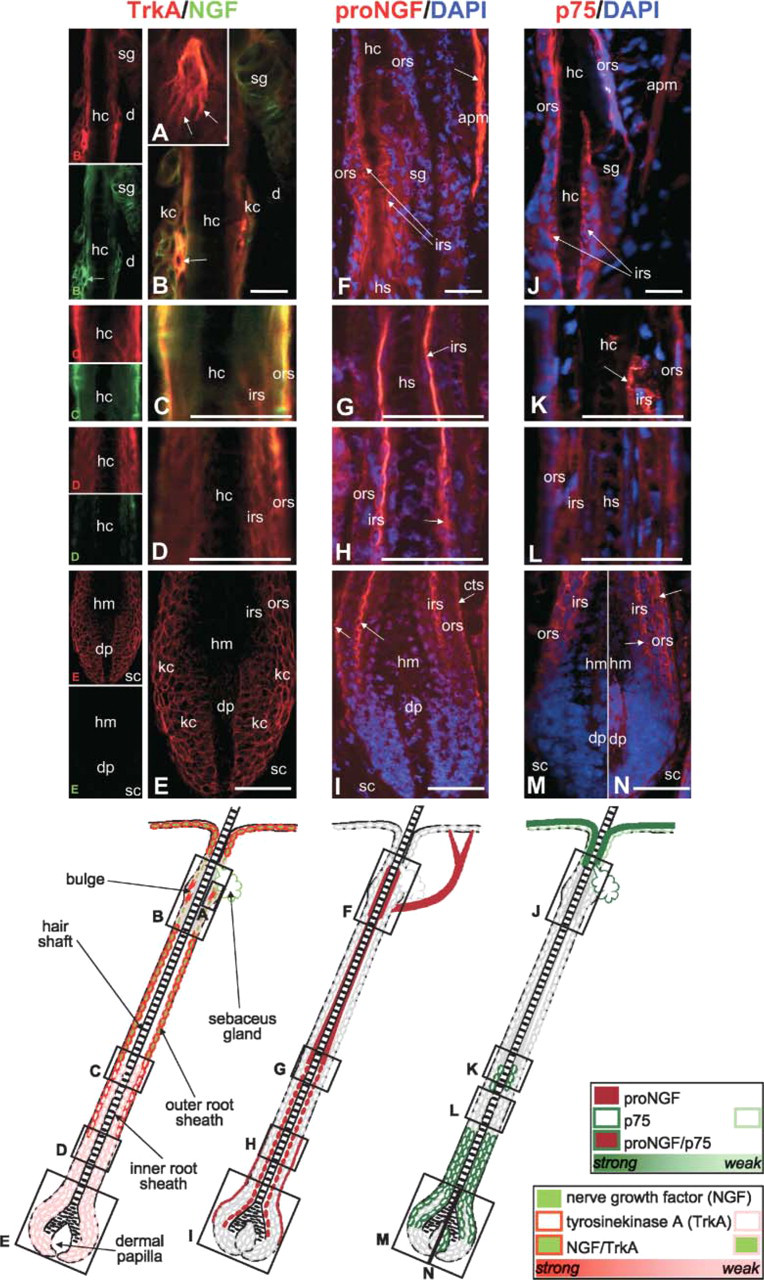
Immunohistochemical localization of nerve growth factor (NGF), TrkA, proNGF, and p75 in the murine anagen hair follicle. Cryosections of murine back skin with all hair follicles in the growing phase of the hair cycle (anagen VI, day 12 after hair growth induction by depilation) were immunolabeled with rhodamine (red) to detect TrkA, proNGF, and p75 protein or with fluorescein (green) to detect NGF as described in Materials and Methods. Cell nuclei have been counterstained with DAPI (blue). Schematic drawings indicate the most representative staining pattern as scrutinized from 10 microscopic fields in five different mice per staining. Black boxes in schematic drawings indicate localization and proportion of the corresponding photomicrographs labeled with the identical upper case letter in the lower left corner. Orange staining indicates overlay of NGF and TrkA staining. (
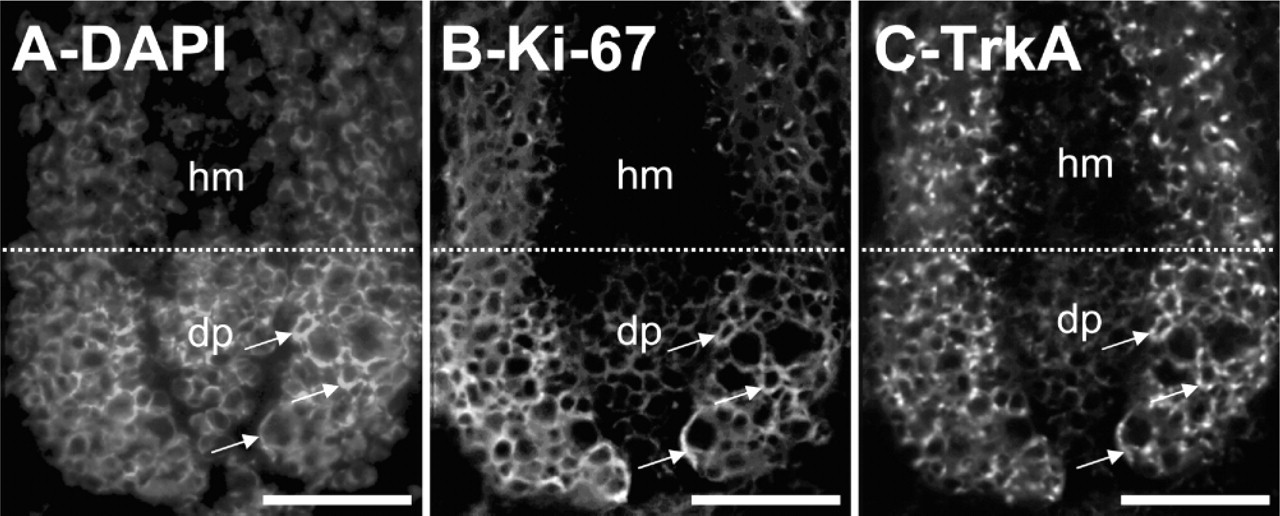
TrkA immunoreactive cells label with the proliferation marker Ki67. Anagen VI skin was double labeled with the antiserum against TrkA and Ki67 and a representative anagen VI hair bulb is shown. (
On the mRNA level, NGF/proNGF is low in catagen skin while it is high on the protein level. High protein levels indicate low turnover and proNGF to NGF processing with subsequent high levels of proNGF. Interestingly, NGF protein can increase in the skin whereas mRNA levels stay stable (e.g., when nerve sprouting is required after injury) (Harper et al. 1999) or, presumably in our case, when proNGF is required for catagen development. In this context, it is also noticeable that mice with a targeted overexpression of NGF in their skin—who presumably also overexpress the precursor—exhibit accelerated catagen development (Botchkarev et al. 2000). Thus, we assume that the catagen induction we observe in our tissue culture model results from proNGF interaction with the p75 receptor available in abundance in the hair follicle at the onset of catagen development, whereas TrkA availability is decreased.
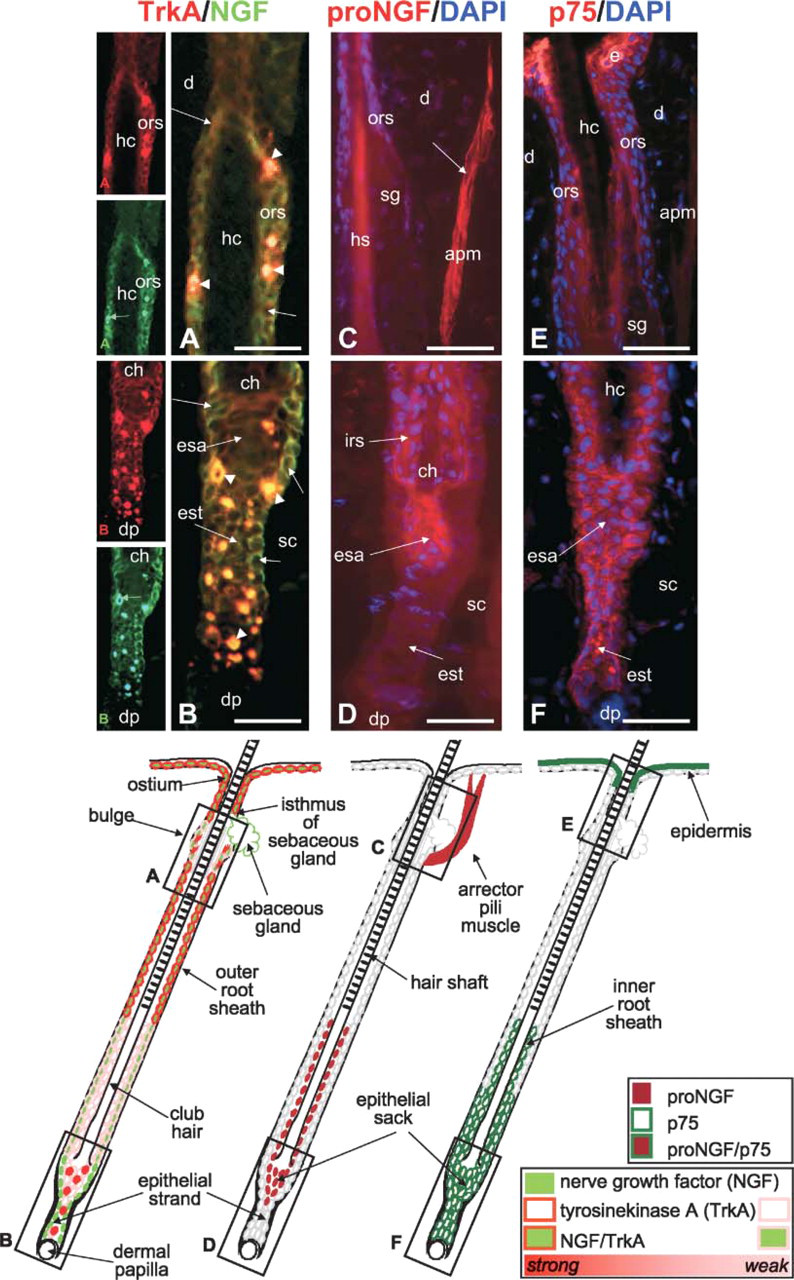
Immunohistochemical localization of nerve growth factor (NGF), TrkA, proNGF, and p75 in the murine catagen hair follicle. Cryosections of murine back skin with all hair follicles in the growing phase of the hair cycle (anagen VI, day 12 after hair growth induction by depilation) were immunolabeled with rhodamine (red) to detect TrkA, proNGF, and p75 protein or with fluorescein (green) to detect NGF as described in Materials and Methods. Cell nuclei have been counterstained with DAPI (blue). Schematic drawings indicate the most representative staining pattern as scrutinized from 10 microscopic fields in five different mice per staining. Black boxes in schematic drawings indicate localization and proportion of the corresponding photomicrographs labeled with the identical upper case letter in the lower left corner. Orange staining indicates overlay of NGF and TrkA staining. (
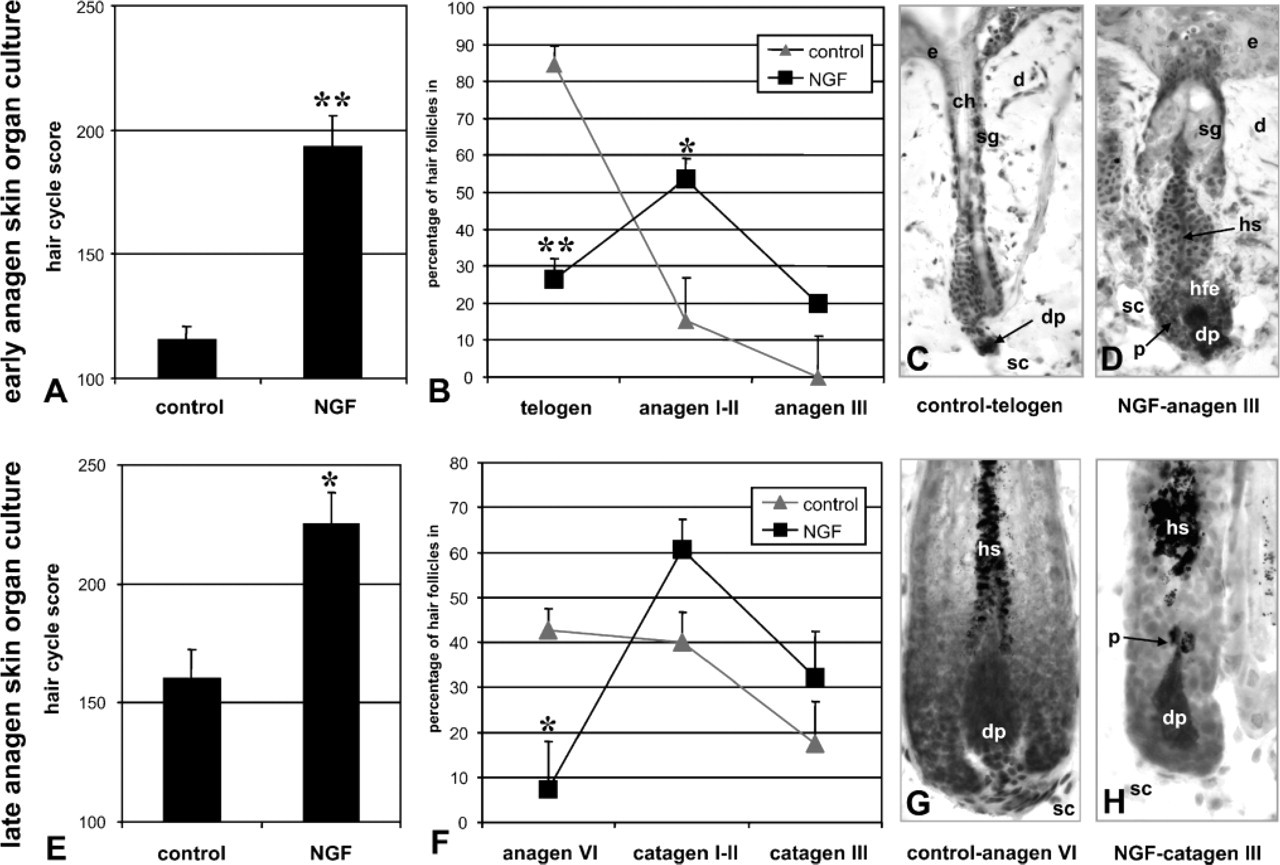
Organ cultured early and late anagen skin responds to nerve growth factor (NGF) treatment. Organ cultured early anagen skin (skin biopsies taken from back skin on day 3 after depilation) exhibits a low hair cycle score (
These complex NGF signaling pathways are not fully understood (Barker 2004), though they carry great implications for the understanding and therapeutic intervention of skin diseases marked by a misbalance between growth and regression, proliferation, and apoptosis of neurotrophin receptor–carrying cells. Here we employ immunohistochemically based histomorphometry and the physiologically occurring tissue remodeling processes during the hair growth cycle to show for the first time that NGF, its precursors proNGF, and their receptors are expressed differentially and functionally distinct in epithelial and mesenchymal cell populations.
This study confirms how useful the cyclic growth, regression and remodeling of the hair follicle—a prototypic, uniquely accessible, and easily manipulated neuroectodermal-mesodermal interaction system (Stenn and Paus 2001)—is as a general model for further dissecting the functions of neurotrophin receptor-mediated signaling in complex peripheral tissue interactions systems under physiologically relevant conditions (Peters et al. 2003; Botchkarev et al. 2004). Even more so, tissue culture models such as the skin organ culture model employed in our study invite testing of novel NGF preparations exclusively containing NGF or proNGF and provide an easy read-out system for new therapeutic approaches to improve treatment of hair growth and other skin disorders such as wound healing.
Our findings have important implications for the future design of neurotrophin research to further explore their role in tissue homeostasis. Not only do they emphasize the necessity to develop specific receptor ligands (e.g., as a novel class of hair growth modulators), but they also emphasize the importance of further research into NGF processing and downstream signaling events specifically in the study of keratinocyte and hair follicle growth control and in a broader sense, wherever NGF signaling is involved in proliferation and apoptosis of epithelial tissues.
Footnotes
Acknowledgements
This study was supported in parts by grants from the Charité University Medicine Berlin, Germany, and the German Research Foundation (DFG Pe 890/1–3, Pa 345/6–4) to E.M.J.P. and R.P.
Many thanks go to Dr. Vladimir Botchkarev, Department of Dermatology, Boston University School of Medicine, Massachusetts, who first established the immunofluorescence detection method for NGF, TrkA, and p75, and generously taught this technique to Dr. Peters. The excellent technical assistance of Ruth Pliet and Evelyn Hagen are gratefully appreciated.