
Abstract
Pleiotrophin (PTN) is a member of the family of heparin-binding growth factors that displays mitogenic activities and promotes neurite outgrowth in vitro. In vivo, PTN is widely expressed along pathways of developing axons during the late embryonic and early postnatal period. Although the level of PTN gene expression is very low during adulthood, activation of the gene may occur during recovery from injury and seems to play an important role in tissue regeneration processes. In this study, we investigated whether PTN was involved in the regenerative process of injured peripheral nerves. To refer localization of the fluorescent markers to myelinated axons, we developed a specific computer tool for colocalization of fluorescence images with phase contrast images. Immunohistochemical analysis showed PTN in different types of nonneural cells in distal nerve segments, including Schwann cells, macrophages, and endothelial cells, but not in axons. Schwann cells exhibited PTN immunoreactivity as early as 2 days after injury, whereas PTN-positive macrophages were found 1 week later. Strong PTN immunoreactivity was noted in endothelial cells at all time points. These findings support the idea that PTN participates in the adaptive response to peripheral nerve injury. A better understanding of its contribution may suggest new strategies for enhancing peripheral nerve regeneration.
Keywords
P
The cellular events that follow transection or crush of a peripheral nerve have been extensively documented. To achieve successful nerve repair, axons have to regrow and find their correct target cells. The first step is removal of myelin debris, a process known as Wallerian degeneration and carried out by resident Schwann cells and infiltrating macrophages. The Schwann cells dedifferentiate, proliferate, and align within basal lamina tubes (bands of Büngner), providing a guidance substrate for growing axons. The regrown axons are then myelinated by redifferentiated Schwann cells (Kury et al. 2001).
In this study, we investigated whether PTN was involved in peripheral nerve regeneration after nerve injury. We used immunohistochemical markers to look for PTN in distal portions of mouse sciatic nerves after crush injury, and we examined spatial and temporal relationships between PTN expression, Schwann cells, macrophages, and endothelial cells during Wallerian degeneration and nerve regeneration. To refer localization of the fluorescent markers to myelinated axons, we developed a specific computer tool for colocalization of fluorescence images with phase contrast images. This system allowed us to investigate, on a same cryostat transverse section, both fluorescent markers and myelinated axons. It must be noted that revealing myelinated axons usually requires a technique that is incompatible with immunocytochemistry (e.g., osmium tetroxyde, toluidine blue).
Materials and Methods
Experimental Animals and Surgical Procedures
All experiments were performed in accordance with the European community's council directive of November 24, 1986. Swiss mice (IffaCredo; L'Arbresle, France) weighing 25–30 g were anesthetized using sodium pentobarbital (50 mg/kg intraperitoneally). For each experiment, 5 to 10 animals were used. The right sciatic nerve was exposed and then crushed at the mid-thigh level for 10 sec with a fine forceps. For Western blotting or immunohistochemistry, the mice were killed at different times after the nerve injury, and nerve segments approximately 10 mm in length were removed distal to the crush; matching segments of the contralateral nerve were isolated in the same way.
Western Blot Analysis
For each time studied, three to five nerve segments (each from a different animal) were pooled and homogenized in 10% w/v lysis buffer (20 mM Hepes [pH 7.4]; 1 μg/ml each aprotinin, leupeptin, and pepstatin; 0.1 mM phenylmethylsulphonyl fluoride; and 3 mM EDTA) containing 2 M NaCl. After centrifugation (12,000 × g for 15 min), the supernatant was diluted 5-fold with lysis buffer, and 1 ml was incubated overnight at 4C with heparin-Sepharose CL-6B beads (Pharmacia; Uppsala, Sweden). The beads were washed, and heparin-Sepharose-bound molecules were then eluted with electrophoresis sample buffer as previously described (Milhiet et al. 1998). Following SDS-15% polyacrylamide gel electrophoresis and transfer to Immobilon-P membranes (Milhiet et al. 1998), immunological detection was performed using anti-PTN (C-19) goat polyclonal IgG (Santa Cruz Biotechnology; Santa Cruz, CA) 1/500 and the ECL system (Amersham; les Ullis, France).
Immunocytochemistry
Mice were killed at different times after the operation, and their sciatic nerves were fixed in situ with 4% paraformaldehyde in 0.1 M phosphate buffer. Nerve segments approximately 10 mm in length were removed distal to the crush (or at matching sites of the contralateral nerve), postfixed for 3 hr with the same fixative, immersed in 15% sucrose in 0.1 M phosphate buffer for 2 days, embedded in gelatin 7.5% sucrose 15% 0.1 M phosphate buffer and frozen with dry-ice isopentane. Transverse sections (10 μm in thickness) were obtained using a cryostat microtome. Nonspecific binding sites were blocked by preincubation with 6% bovine serum albumin (BSA) in phosphate-buffered saline (PBS) for 30 min. Sections were then incubated overnight at 4C in primary antibodies diluted with BSA 1%-Triton 0.4%-PBS: anti-PTN (C-19) goat polyclonal IgG 1/50 (Santa Cruz Biotechnology); anti-ED1 mouse monoclonal IgG 1/50 (Serotec, Toronto, Canada); anti-S100 rabbit polyclonal IgG 1/50 (Dako; Glostrup, Denmark). Sections were then rinsed 3-fold with PBS and incubated for 2 hr at room temperature with secondary antibodies (Jackson; Soham, UK): TRITC donkey anti-goat 1/200; FITC donkey anti-mouse 1/200; FITC donkey anti-rabbit 1/200; or TRITC goat anti-rabbit 1/200. Control sections were processed at the same time and in the same way except that PBS was used instead of the primary antibody. No stained structures were seen in the controls. For nuclear staining, after immunolabeling, some sections were incubated in Hoechst 33,258 0.01 μg/ml (Molecular Probes; Eugene, OR) for 10 min. For double labeling, two primary antibodies, raised in different animal species, were mixed and a mixture of the two corresponding secondary antibodies was then used.
Image Acquisition and Processing
Digital Microscopy and Image Analysis Software. Conventional microscopy images were obtained using a C-MOS monochrome imaging device (C-MOS PRO; Matco) adapted to a BH-2 epi-fluorescence optical microscope (Olympus, France) with a C-mount optical adaptor (x0.3). Confocal microscopy observations were performed with a Zeiss LSM410 Confocal Axiovert 135M inverted microscope (Zeiss, Germany), using the 488 nm emission ray of an Ar/Kr laser for both transmission and fluorescence mode. Confocal microscopy images treatments were performed with ImageJ software, and original developments were made with NIH Image software (both developed by Wayne Rasband at the National Institutes of Health [Bethesda, MD] and available at http://rsb.info.nih.gov/nih-image/). Three-dimensional volume rendering was obtained with MacVol software, which is based on the VolPack rendering library of the Stanford Computer Graphics Laboratory (Lacroute and Levoy 1994).
Computer Colocalization Image Processing. The complexity of the histological material used for our study led us to develop a specific computer tool for colocalization of pseudo-color fluorescence images and triple channel fluorescence composite images with phase-contrast images, to facilitate localization of the fluorescent regions. The phase-contrast images were taken in the exact focal plan of the fluorescence images, and the slight blur resulting from the difference in optical pathways was corrected using an unsharp-masking filter (gaussian convolution matrix size 5 × 5, sd = 1). A master image presenting the pseudo-color fluorescence images, the composite image of three channels fluorescence and their combination with phase-contrast images was generated. By selecting a region of interest on any of these representations, all possible combinations could be obtained.
We elected to use horizontal representations containing a region of interest viewed in phase contrast, pseudo-color, and/or composite modes, and the combination of phase-contrast and composite representations.
Results
High levels of PTN were detected in sciatic nerves within the first few days after injury. To locate PTN protein in sciatic nerves, we performed immunochemical studies with anti-PTN antibodies. The specificity of the antibodies used in this study was first tested via Western blotting analysis with purified human recombinant PTN (Figure 1, Lane E, 50 ng), heparin-purified normal sciatic nerve extracts from the nonoperated nerve (Lane A and C), and heparin-purified sciatic nerve extracts from the first (Lane B) and fourth (Lane D) weeks after injury. The anti-PTN antibodies recognized sciatic nerve PTN as a single 18-kDa band (the apparent molecular weight of the unprocessed precursor). High PTN levels were detected within the first week after injury (Lane B), whereas very low levels were found in normal nerves (Lane A and C) and 4 weeks after injury (Lane D).
Two days after injury, numerous PTN-positive cells were observed in the distal portion of the nerve. In transverse sections of the uninjured sciatic nerve (control), after immunostaining, nonneural cell nuclei were visible between myelinated axons. These cells exhibited no PTN immunoreactivity (Figures 2A–2C). Conversely, in the injured nerve distal to the crush, numerous nonneural cells exhibited PTN immunoreactivity in their cytoplasm (Figures 2D–2F), whereas no PTN staining of axons was visible in the same sections. Among these nonneural PTN-positive cells, some had small nuclei and were placed alongside each other (Figures 2G–2I), indicating that they were probably new Schwann cells produced by proliferation. These cells were located in areas with marked axonal disintegration. Some cells had PTN-positive processes, connecting two cells (Figures 2J–2L).
Among PTN-positive nonneural cells, some expressed S-100 protein. Among nonneural cells stained for PTN, we found S-100-positive and S-100-negative cells (Figures 3A–3D,3M–3P). These two types of cells were closely associated. Among PTN-positive, S-100-negative cells, we were able to distinguish endothelial cells derived from blood vessels (Figures 3M–3P) and cells in their immediate proximity, which were probably macrophages (Figures 3M–3P). Most of the small PTN-positive grouped cells were S-100-negative (Figures 3E–3H, 3I–3L) and were located in areas with marked axonal disintegration. S-100-positive cells, negative for PTN, were also seen (Figures 3I–3L) as tongues of cytoplasm into the empty space inside myelin sheath (Figures 3E–3H).
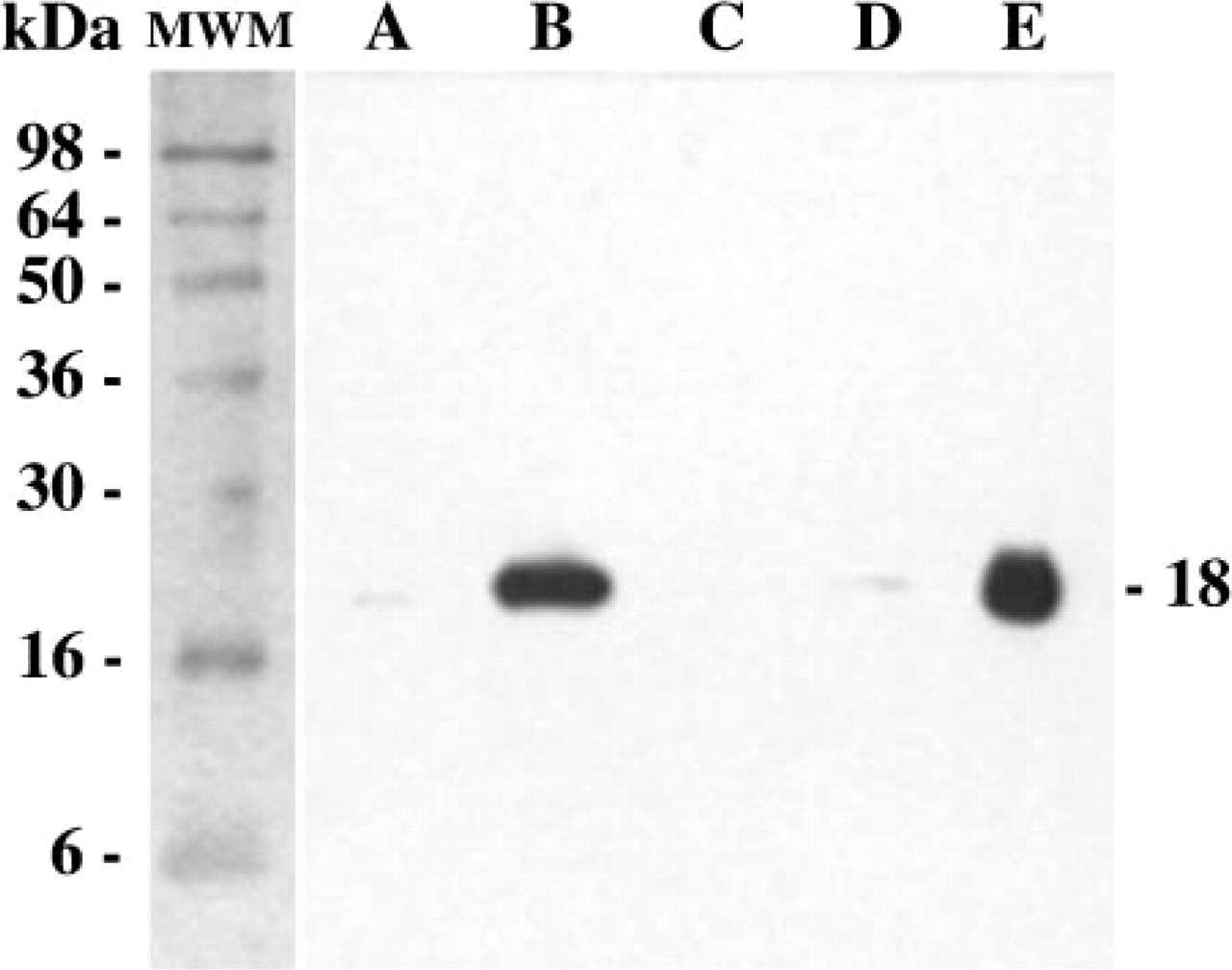
Western blot analysis of sciatic nerve extracts probed with anti-PTN antibodies. Lanes A and C: normal sciatic nerve extracts from the unoperated side. Lane B: sciatic nerve extract 1 week after injury. Lane D: sciatic nerve extract in the fourth week after injury. Lane E: human recombinant PTN, 50 ng. MWM = molecular weight markers.
ED1-positive cells were detected as early as the second day after injury, but their numbers increased dramatically during the second week, with a strong modification of their cellular aspect. Two days after injury, few small cells exhibiting moderate ED1 immunoreactivity were seen (Figure 4A). One week later ED1-positive macrophages invaded the distal portion of the crushed nerve. The morphology of these ED1-positive cells changed to the foamy profile characteristic of phagocytes. These cells were large, with abundant vacuolated cytoplasm, suggesting that these cells are engaged in phagocytosis of myelin debris (Figure 4B). Confocal analysis (Figures 4C–4F) shows marked ED1 staining of the vacuoles.
During the second week, ED1-positive macrophages expressed marked PTN-immunoreactivity or were closely associated with PTN-positive cells. At the end of the second week, the distal stump of the crushed nerve was largely replaced by regenerated myelinated structures that stained for the S-100 marker (Figure 5A). Few PTN-positive Schwann cells were seen (not shown). Numerous ED1-positive and S-100-negative macrophages were visible (Figures 5B–5C). These macrophages were often strongly PTN-positive (Figures 5D–5F); some macrophages were PTN-negative but were closely associated with PTN-positive cells or cellular debris (Figures 5G–5I).
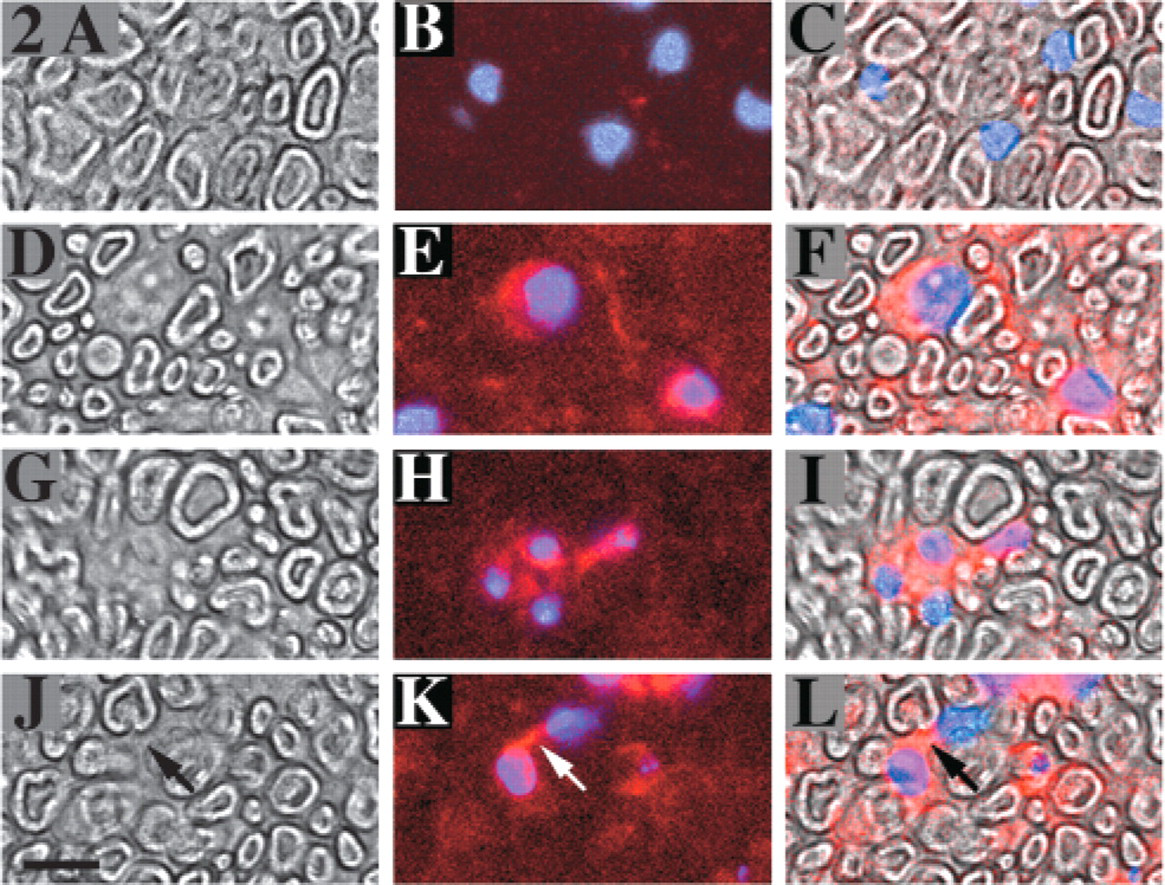
(
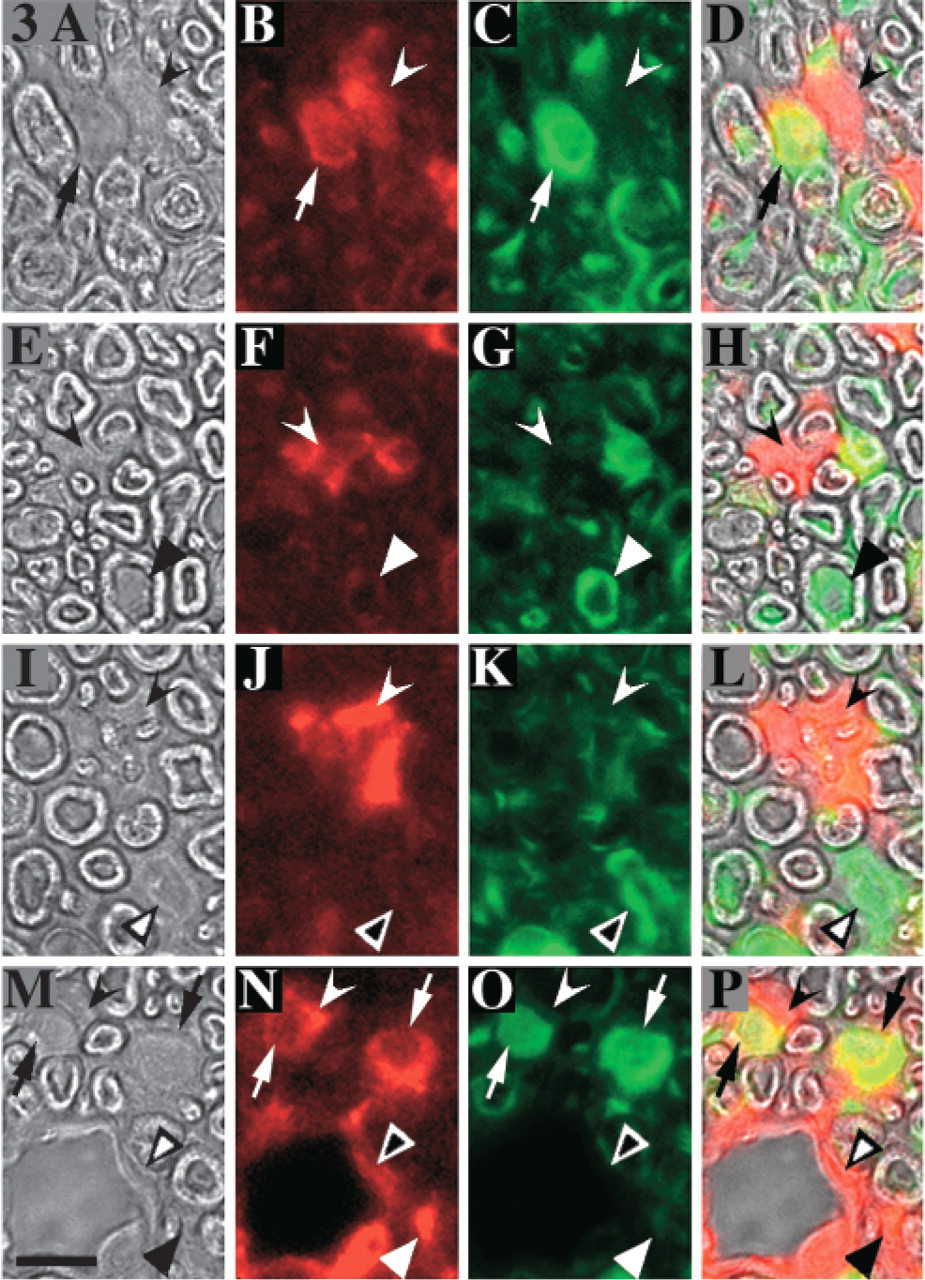
Distal portion of crushed nerve 2 days after injury. The four pictures in each row were obtained from the same region of interest, observed in phase contrast and in fluorescence after a double-staining by anti-PTN (red) and anti-S-100 (green). (
After 3 weeks, no PTN immunoreactivity was detected, except in endothelial cells. In the fourth week after injury, slender regrowing axons with thin myelin sheaths were visible (Figures 6A and 6E). These myelin sheaths faintly expressed S-100 immunoreactivity (Figures 6C and 6D), whereas PTN immunoreactivity was undetectable in axons and myelin structures (Figure 6B). Numerous nonneural cell nuclei were present (Figures 6F and 6G), and endothelial cells at the periphery of the microvasculature were PTN-positive (Figures 6E–6G).
Discussion
The main findings from this study are: (1) high PTN levels in sciatic nerve extracts in the first week after injury, contrasting with very low PTN levels in normal nerves or nerves removed several weeks after injury; (2) the presence of PTN in various types of nonneural cells in the distal portions of nerves after injury, including Schwann cells, macrophages, and endothelial cells, contrasting with absence of PTN in axons; and (3) differences in the timing of PTN expression in these various cell types, with Schwann cells exhibiting PTN immunoreactivity as soon as 2 days after injury, PTN-positive macrophages being seen 1 week later, and endothelial cells showed strong PTN immunoreactivity at all timepoints.
High PTN levels were observed in sciatic nerves within a few days after crush injury, when axon regrowth was induced, whereas PTN levels were low after the axons reached their target. It is worth pointing out that PTN was initially identified as a molecule that induced the neurite outgrowth of fetal neurons (Rauvala 1989). Furthermore, strong PTN expression has been found along the pathways of developing axons during late embryonic and early postnatal periods (Rauvala et al. 1994). The increased PTN protein levels during the first step of peripheral nerve regeneration suggest time-restricted synthesis of PTN within the injured nerve. Using anti–S-100 antibody for Schwann cell identification (Kahn et al. 1983) in the distal portion of the nerve after crush, we found that many of these Schwann cells showed strong PTN immunostaining within the first few days after injury, suggesting PTN synthesis. In keeping with this finding, a previous study showed that PTN transcripts were produced by Schwann cells (Mashour et al. 2001). The strong PTN immunostaining observed in Schwann cells a few days after injury coincided with the time of axonal degeneration in the distal part of the lesioned sciatic nerve in adult rodents (Brown and Watson 1991), indicating that Schwann cells become PTN-positive after loss of axonal contact. In contrast, axons were not immunostained by anti-PTN.
In response to nerve injury, Schwann cells divide and form long chains of cells, known as bands of Büngner, which provide a substrate for axonal regeneration (Bunge 1980). It is well known that denervated Schwann cells increase their production of several neurotrophic molecules, including nerve growth factor (Heumann et al. 1987) or brain-derived neurotrophic factor (Meyer et al. 1992). Axon–Schwann cell interaction mediated by neurotrophic factors may play a pivotal role in peripheral nerve regeneration (Ramon y Cajal 1928; Liu et al. 1995). Increased production of trophic factors by Schwann cells and fibroblasts may substitute for the usual target-derived trophic factors. The signals that induce these complex changes are not known and the interactions among various nonneural cells are incompletely understood, but the data presented in this study suggest a contribution of PTN to these mechanisms.
After nerve injury, the damaged nerve tissue must be eliminated. In particular, myelin debris inhibits axon growth and must therefore be removed. Active Schwann cell division occurs in the distal stump following nerve injury, where Schwann cells act as phagocytes to digest axons and their myelin sheaths (Beuche and Friede 1984; Nishio et al. 2002). Several nonneurotrophin growth factors may potentially participate in the proliferative phase of peripheral nerve repair; examples include fibroblast growth factor II (Timmer et al. 2003), insulin-like growth factor (Kanje 1991), or vascular endothelial growth factor (Zochodne 2000). Some of these molecules, together with PTN, may act as autocrine mitogens for Schwann cells or as chemoattractants for macrophages (Tofaris et al. 2002) and contribute to the neovascularization that is key to successful nerve regeneration. Although the chain of early events resulting in myelin clearance during Wallerian degeneration is performed by Schwann cells alone, these cells must subsequently receive help from hematogenous macrophages (Hirata et al. 1999). In our study, we used the monoclonal antibody ED1 as a marker for macrophages. This antigen is expressed on the membrane of phagolysosomes, as well as on the cell surface of macrophages and the level of ED1 expression in a single cell can be correlated to phagocytic activity of the cell (Damoiseaux et al. 1994). During the second week after nerve injury in our study, numerous macrophages, strongly stained for ED1, invaded the distal portion of the crushed nerve, where they probably eliminated myelin debris via phagocytosis. These macrophages were usually strongly positive for PTN; some were PTN-negative but were associated with PTN-positive cells or cellular debris. It is possible that myelin debris was released by Schwann cells, then phagocytized by macrophages as previously reported by Hirata and collaborators (Hirata et al. 1999). Interestingly, in the central nervous system, a very striking increase in the levels of expression of the PTN gene was found in microglia and macrophages after acute ischemic brain injury (Yeh et al. 1998; Poulsen et al. 2000). In spite of these results, strong PTN immunoreactivity in macrophages could reflect phagocytosis of PTN-expressing cell debris and not only de novo synthesis of PTN.
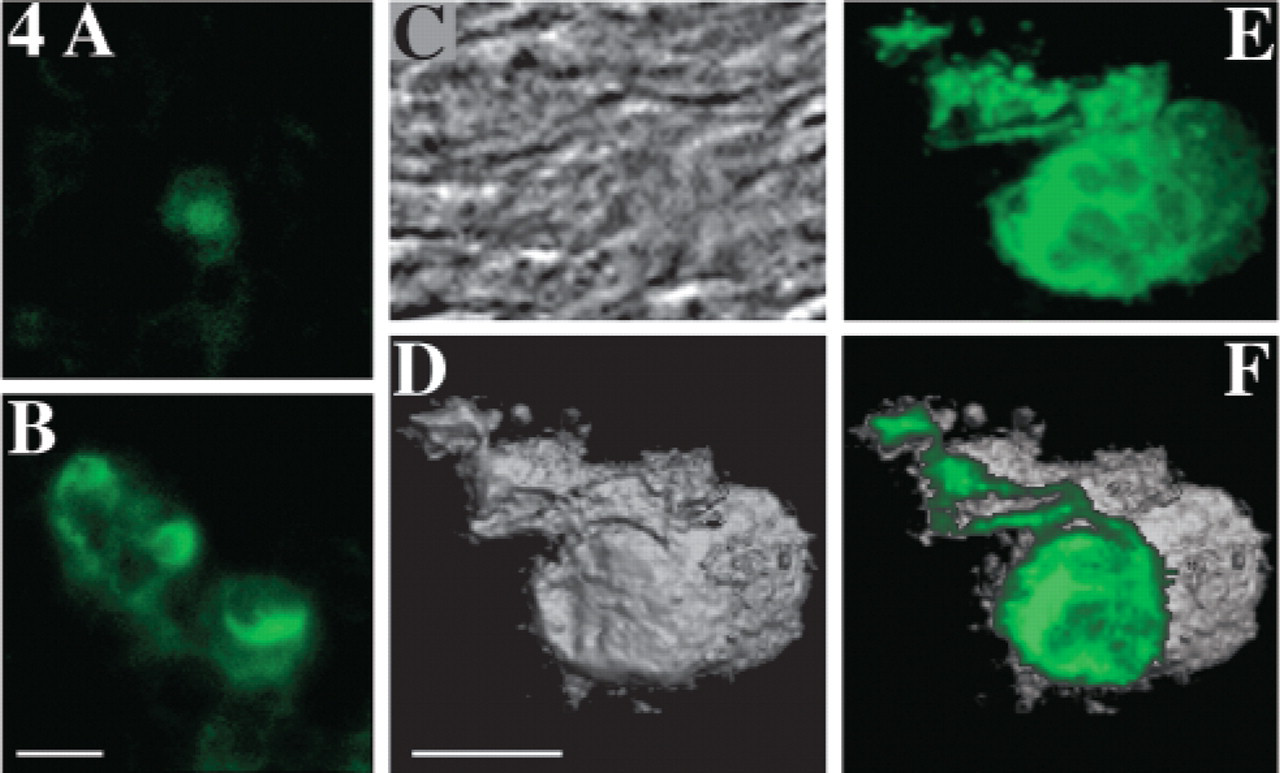
Morphology of ED1-positive cells. (
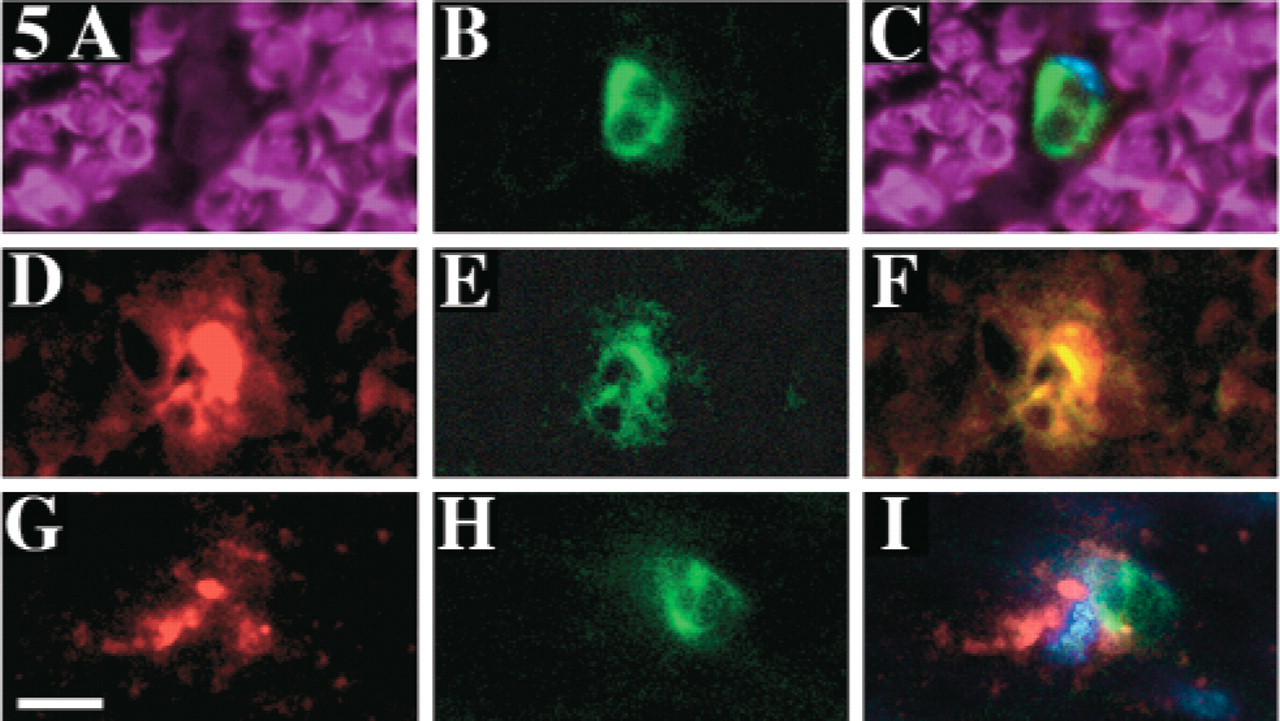
Distal portion of crushed nerves in the second week after injury. The three pictures in each row were obtained from the same region of interest after double-labeling with anti-S-100 (magenta), anti-PTN (red), and anti-ED1 (green). Hoechst 33,258 (blue) staining was made in
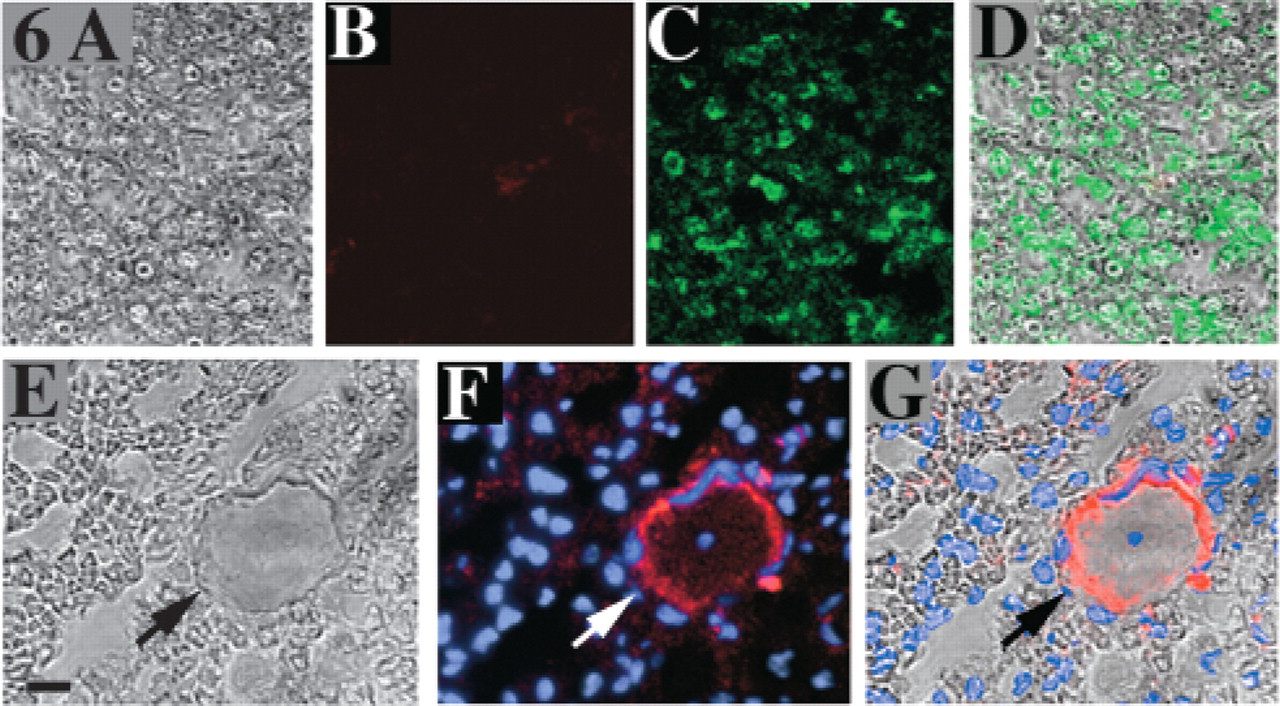
Distal portion of crushed nerves after 3 weeks. For each row, the images were obtained from a same region of interest in phase-contrast and in fluorescence. The first row shows double-staining with anti-PTN (
These observations raise questions about the function of PTN in macrophages and its potential role in recruiting monocytes/macrophages toward the regenerative tissue. However, the demonstration of a direct role of PTN in recruiting monocytes/macrophages by chemotaxis or haptotaxis requires further in vitro investigations. In addition, macrophages contribute to the secretion of mitogenic factors promoting the proliferation of both Schwann cells and fibroblasts (Hirata et al. 1999).
Another role of PTN is to stimulate proliferation of endothelial cells (Courty et al. 1991; Fang et al. 1992). Compelling evidence that PTN contributes to regulate angiogenesis has been reported (Laaroubi et al. 1994; Delbe et al. 1999). In our study, we detected strong PTN immunoreactivity in endothelial cells from the first to the fourth week after injury. PTN expression by endothelial cells has been reported previously. For example, PTN mRNA and protein have been detected in endometrial capillary endothelial cells, and upregulation of PTN mRNA has been found during the revascularization phase of the endometrial cycle (Milhiet et al. 1998). Yeh and collaborators (Yeh et al. 1998) have also shown increased PTN expression in the endothelial cells of the newly formed vessels after ischemic brain injury. More recently, we found that PTN and the corresponding peptides 1–21 and 121–136 stimulated endothelial cell proliferation and migration (Papadimitriou et al. 2001). Promoting neovascularization may be an important function of PTN after brain or nerve injury.
Taken together, these findings indicate that PTN participates in the adaptive response to peripheral nerve injury, at least in part via partial replication of the molecular and cellular mechanisms that operate during development.
In conclusion, our results suggest that PTN may contribute to nerve regeneration, in addition to its already-described roles in neuronal survival, cell proliferation, and angiogenesis. However, its exact implication has yet to be defined. A better understanding of its contribution may generate new strategies for enhancing peripheral nerve regeneration.
Footnotes
Acknowledgements
This work was supported by grants from Ministère de l'Education Nationale and Centre National de la Recherche Scientifique (CNRS, a publicly funded research institute).