
Abstract
Adrenomedullin (AM) is a potent vasodilator peptide present in the lung of mammals where it is expressed mainly in the columnar epithelium and alveolar macrophages. AM increases the secretion of phosphatidylcholine by type II pneumocytes, which suggests a role as an autocrine modulator of surfactant secretion. In this study we show the expression of an AM-like protein in the lung of the pigeon, Columba livia. Using an antibody against its human ortholog, AM-like immunoreactivity was found to be associated with membranous structures of the multivesicular bodies of type II pneumocytes. We also studied the differential expression of AM-like peptide in the lung of pigeons exposed to polluted city air vs cleaner countryside conditions and found that AM-like expression was higher in city animals. Similar results were obtained in an experimental study in which pigeons were exposed to increasing concentrations of a single pollutant, ozone. Taken together, our findings support the implication of AM in the response of type II pneumocytes to air pollutants.
S
In mammalian lung, AM is expressed in the columnar epithelium of bronchi and bronchioli, in some glands, endothelial cells, alveolar macrophages, neurons of the pulmonary parasympathetic nervous system, smooth muscle cells, and chondrocytes (Martínez et al. 1995). The AM receptor constituted by calcitonin receptor-like receptor (CRLR) and receptor activity modifying protein (RAMP2) is also abundantly expressed in mammalian lung (Martínez et al. 1997; Ono et al. 2000). With relation to its physiological role in lung, AM contributes to pulmonary vasodilatation and inhibition of bronchoconstriction (Yang et al. 1996). Increased levels of circulating AM have been reported in several respiratory pathologies such as asthma (Kanazawa et al. 1996; Kohno et al. 1996; Ceyhan et al. 2001).
It is also known that circulating AM is removed when it passes through the pulmonary vascular bed (Yoshibayashi et al. 1997), and an anti-inflammatory role has been suggested for AM in this organ because this peptide inhibits alveolar macrophage release of neutrophil chemoattractants in response to LPS (Kamoi et al. 1995).
AM increases the secretion of phosphatidylcholine, the predominant lipid component of pulmonary surfactant, in type II pneumocytes in culture (Okumura et al. 2000). Pulmonary surfactant is a complex mixture of lipids and proteins that reduces the surface tension in the alveolar air-liquid interface (Chander and Fisher 1990). There are two main surfactant reservoirs: the intracellular surfactant contained in the lamellar bodies of type II pneumocytes and the extracellular surfactant that lines the alveolar surface (Wright and Clements 1987; Risco et al. 1994; Haller et al. 1998). Defects in the synthesis and excretion of surfactant, as well as alterations in its components, result in a disturbance of the alveolar gas exchange rate, as has been reported in certain pathologies such as the respiratory distress syndrome (Hallman et al. 1982; van Golde et al. 1988).
In spite of the large amount of references related to the presence of AM in lung, little is known about the expression of this peptide in the lung of non-mammalian species. Moreover, although the function of AM in lung is well known, its physiological relationship with the surfactant system and the relevance of AM secretion in response to stress situations, such as the prolonged exposure to air pollutants, is still poorly understood. Air pollutants have been recognized to influence the structure and function of the surfactant system, resulting in increased synthesis of surfactant components but, at the same time, they reduce surfactant secretion (Müller et al. 1998). In avian species, it has been shown that air pollution causes a reduction in the number of lamellar bodies present in type II pneumocytes (Lorz and López 1997).
In the present study we focus on the distribution of AM-like peptide immunoreactivity in the lungs of animals raised in two different habitats: a polluted city and a non-contaminated village. In addition, to better understand the contribution of individual pollutants, we have designed an in vivo experimental model that allows for controlled exposure to different air pollutants. Using this model, we have studied the effects of ozone on AM expression. The animal species chosen was the common pigeon (Columba livia), because the clustered localization of type II pneumocytes covering the parabronchial surfaces in birds (López 1995; Klika et al. 1999) makes it easy to perform studies related to their physiology.
In this study we demonstrate the presence of an AM-like peptide in avian lung and describe the effects of air pollution on the expression of this peptide in type II pneumocytes.
Materials and Methods
Animal Model
Eighteen specimens of Columba livia domestica were used for this study; six were collected from a farm located 50 km away from the closest industrial city, Guadalajara, Spain (referred to in the text as “countryside pigeons”). Another subset of six pigeons was captured in the city of Madrid, Spain (“city pigeons”). The six remaining specimens were used in the experimental model for ozone and were purchased from a farm in Valencia, far from the nearest city. All specimens were checked by a veterinary service before performing the experiment and were found to be healthy.
All protocols were approved by the Ethics and Animal Care Committee of the Universidad Autónoma de Madrid. Atmospheric levels of noxious agents in the city of Madrid during the month when the specimens for this comparative study were captured are shown in Table 1.
Ozone Exposure System
Because a polluted atmosphere consists of a mixture of air pollutants in different concentrations, the comparative study provides evidence of the effects of chronic exposure to pollution levels such as those found in an industrialized city. To further understand the effects of individual contaminants on the expression of AM-like peptide in the lung of the pigeon, we developed a model that allowed for the study of controlled exposures to single pollutants. For this model, we chose the open-top chambers that had been successfully used in studies of ozone exposure in vegetables (Gimeno et al. 1989). The ozone exposure system comprised three open-top chambers, each consisting of a frame surrounded by clear polyethylene plastic and perforated at the bottom to allow air circulation (Heagle et al. 1989). Three different treatments were used in each chamber: non-filtered air (NF), air enriched with 60 μg/m3 O3 (30 ppb), and air enriched with 200 μg/m3 O3 (100 ppb). Ozone was pumped daily into the chambers from 8:00 am to 4:00 pm during 5 consecutive days, representing a typical exposure to ozone for an urban environment. The ozone concentration inside the chambers was monitored continuously with an EPA-approved monitor. A representative daily average of the ozone concentrations in the chambers is depicted in Figure 1. Two pigeons per treatment were used. The animals were placed in cages in the middle of the chamber and were fed cracked corn and sterile water. After 5 days exposure, the lungs were processed as explained below.
For this study, the concentrations of ozone always remained within the range in which they could be found in a polluted city atmosphere. Animals breathing NF air were used as controls.
Average concentration of common pollutants in the city of Madrid in the month when the pigeons were captured and the average for the same year a
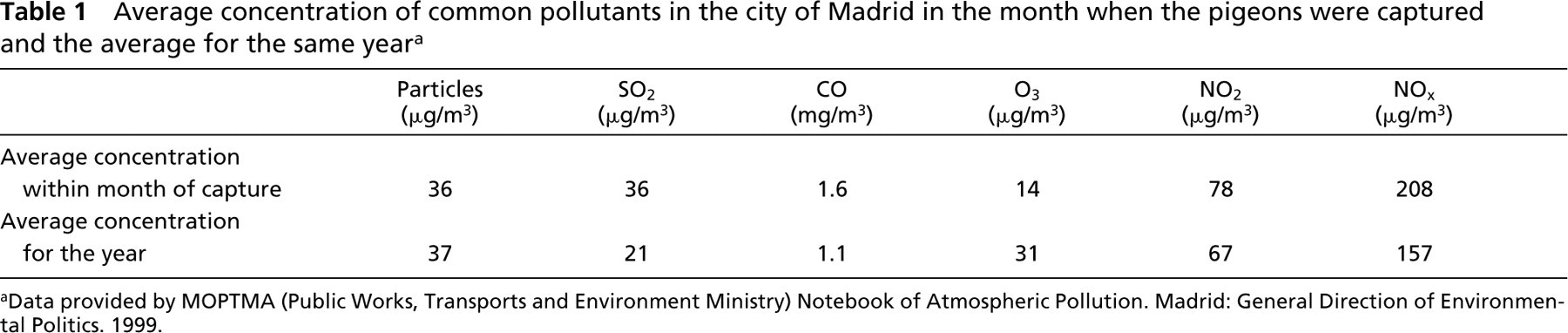
aData provided by MOPTMA (Public Works, Transports and Environment Ministry) Notebook of Atmospheric Pollution. Madrid: General Direction of Environmental Politics. 1999.
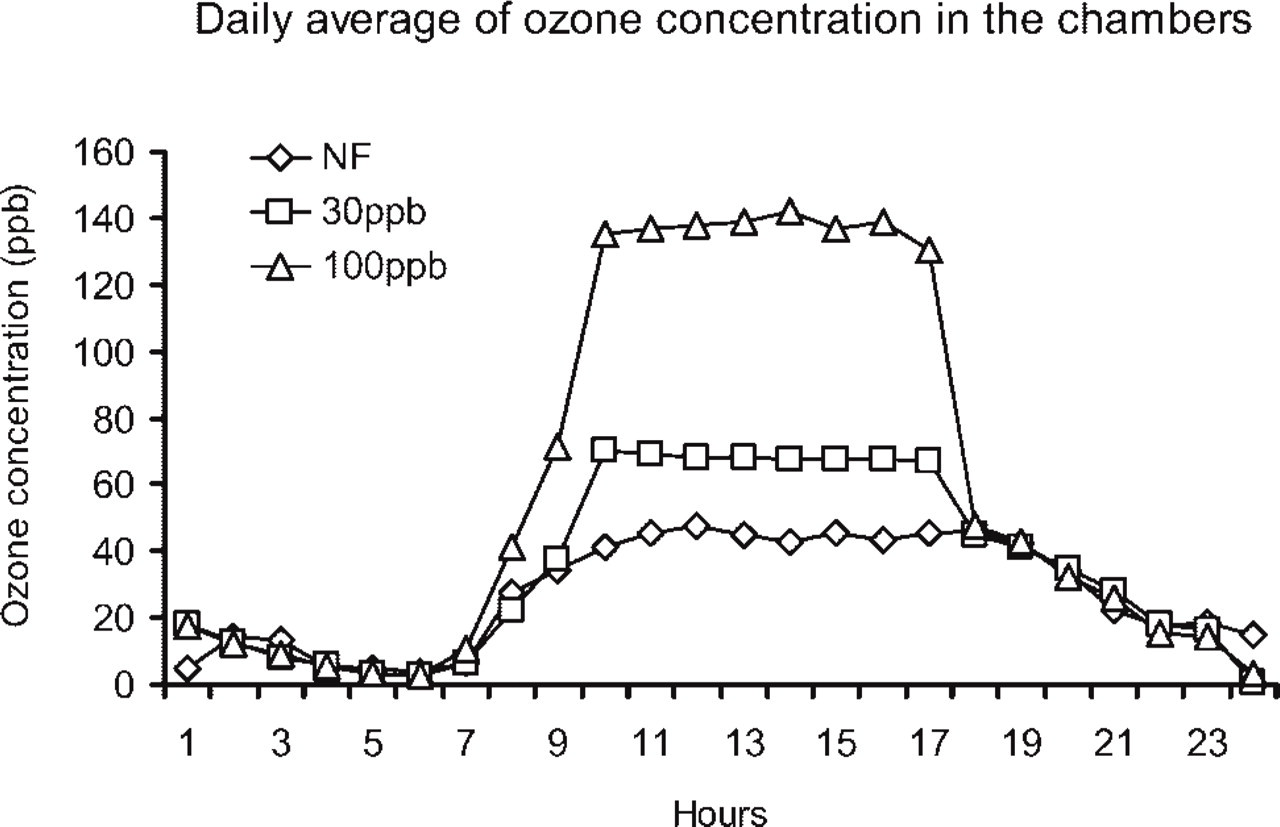
Representative example of the daily fluctuations in ozone concentration inside the three chambers used to expose the pigeons. Ozone was added to the chambers during 8 hr each day, for a period of 5 consecutive days. One chamber contained non-filtered air (NF), and the graph shows the average ozone concentration present in the atmosphere during the experiment. The air in the other two chambers was enriched with 60 μg/m3 O3 (30 ppb) and 200 μg/m3 O3 (100 ppb), respectively.
Western Blotting
Tissue samples destined for Western blot analysis were frozen in liquid nitrogen immediately after dissection and maintained at − 80C. Extracts were prepared by thawing the tissue on ice and homogenizing the samples in ice-cold homogenization buffer: 2x Tricine buffer with 8% SDS (Novex; San Diego, CA) containing 1 mM final concentration of the protease inhibitors pefablock (Centerchem; Stanford, CT), bestatin, and phosphoramidon (Sigma; St Louis, MO). The homogenate was centrifuged at 100,000 × g for 30 min, and the supernatant was collected. Homogenate supernatant protein content was measured using a BCA kit (BioRad Labs; Richmond, CA). Protein extracts were diluted to an approximate protein concentration of 1 μg/μl and were electrophoretically fractionated on a 12% Tricine SDS-PAGE gel (Novex) under reducing conditions (5% β-mercaptoethanol). Proteins were transferred to a polyvinyldifluoride membrane (Immobilon PVDF; Millipore, Billerica, MA). The membrane was blocked overnight in 5% non-fat milk-PBS, incubated for 1 hr in a 1:1000 dilution of rabbit anti-AM (Martínez et al. 1995), and washed three times in 0.1% Igepal (Rhodia; Cranbury, NJ). The membrane was further exposed to biotinylated goat anti-rabbit immunoglobulins (1:200 dilution; Dakopatts, Glostrup, Denmark) for 1 hr and then exposed to avidin-biotinylated peroxidase complex (1:500; Dakopatts) for an additional hour. Peroxidase activity was revealed with the ECL + Plus chemiluminiscence kit (Amersham; Arlington Heights, IL) following manufacturer's instructions. The specificity control consisted of duplicate membranes incubated in antigen-preabsorbed (10 nmol/ml) antiserum.
Immunohistochemistry in Paraffin Sections
Pigeons were anesthetized with a mild dose of chloroform and then decapitated before proceeding to the dissection. The lungs were fixed by immersion in 4% buffered formalin for 24 hr and then embedded in paraffin and sectioned following routine procedures. The avidin-biotin histochemical staining procedure (Hsu et al. 1981) was used to localize AM in the lungs from city and countryside pigeons and in the experimental model for ozone. After deparaffinizing in xylene, tissue sections were rehydrated, and antigen retrieval was performed with a citrate buffer solution (0.01 M citric acid, pH 6) for 20–30 min in a microwave oven (maximum power). Briefly, slides were blocked with normal swine serum (1:20 dilution) and incubated overnight at 4C with the primary serum: rabbit anti-AM (1:500 dilution). Developing was performed using DAB (50 mg/100 ml) (Sigma) and H2O2 as a substrate. The sections were lightly counterstained with Harris hematoxylin, dehydrated, and mounted for analysis. Rat pancreas was used as a positive control for AM immunostaining. Substitution of the primary antibody by a prebleed serum was used as the negative control. To ensure the specificity of the immunostaining, liquid phase and solid phase absorption controls were performed with the synthetic antigen at a 10 nmol/ml concentration.
For the experiments in which we quantified the intensity of the immunostaining, reagents for avidin-biotin complexes were obtained from the Alkaline Phosphatase Substrate Kit (Vector Labs; Burlingame, CA), and Fast Red TR/Naphthol (Sigma) was used as the substrate. Experimental conditions for the immunolabeling, including batches of reagents and time of Red Naphthol development, were kept constant across all experiments.
Quantification of the Immunohistochemical Reaction
Three equivalent sections of the lung from each experimental pigeon were randomly selected and photographed with a Zeiss III photomicroscope (Zeiss; Oberkochen, Germany), using the same light intensity. Pictures were scanned with a minimal resolution of 300 ppi, and the immunostaining intensities were quantified by computerized densitometric analysis using Photoshop software (version 5.5; Adobe Systems, San Jose, CA) as described (Lehr et al. 1999; Zudaire et al. 2004). Color images were transformed into black and white, setting the gray level of all pixels between 0 and 255. The integrated optical density (IOD) was measured in each experimental group.
Electron Microscopy
Small pieces of 1 mm3 of the lungs from countryside and city pigeons were fixed in 4% paraformaldehyde plus 0.1% glutaraldehyde in PBS at 4C for 4 hr. They were then washed overnight in 0.25 M sucrose in PBS at 4C. The samples were all dehydrated and embedded in epoxy resin, TAAB-812 (TAAB Labs; Aldermaston, Berkshire, UK). The single immunogold staining technique was applied to ultrathin sections (60–80 nm) as follows: after an incubation in 3% H2O2 for 5 min at room temperature followed by three washes in bidistilled water, the sections were exposed to normal goat serum for 30 min. The primary antibody for AM, diluted 1:100 in 0.1% BSA-TBS, was added overnight at 4C. The sections were then rinsed in 1% BSA-TBS and 0.5% Tween-20 prior to incubation with the secondary antibody, goat anti-rabbit IgG 20-nm gold colloidal particles (E-Y Labs; San Mateo, CA) for 45 min (1:30 dilution). They were then rinsed and double stained with 5% aqueous uranyl acetate (15 min) and lead citrate (5 min) and examined in a JEOL JEM-1010 electron microscope (JEOL; Tokyo, Japan). Electronographies were obtained with a Multiscan GATAN camera and digitalized with Digital Micrograph 3.1 software (Gatan; Pleasanton, CA).
Statistical Analysis
Staining intensities for AM were expressed as mean IOD ± SEM. Differences between countryside and city pigeons and among the three experimental groups exposed to ozone were compared using Student's t-test. Statistically significant differences were indicated by p<0.05. Data were analyzed with SPSS 6.1.3 (SPSS; Chicago, IL).
Results
AM-like Immunoreactivity in the Lung of Columba livia
The expression of AM in the lung of the pigeon (Columba livia) has been studied by both Western blot and immunohistochemical methods.
A clear AM-like immunoreactive band with a molecular weight of 18 kDa, which presumably represents the AM-like precursor, was detected in lysates from all tested organs of the pigeon, with the lung showing the strongest immunoreactivity (Figure 2). This band was no longer visible after preabsorption of the antiserum with its antigen, demonstrating specificity of detection (data not shown). The antibody did not cross-react with any other proteins in the lysates.
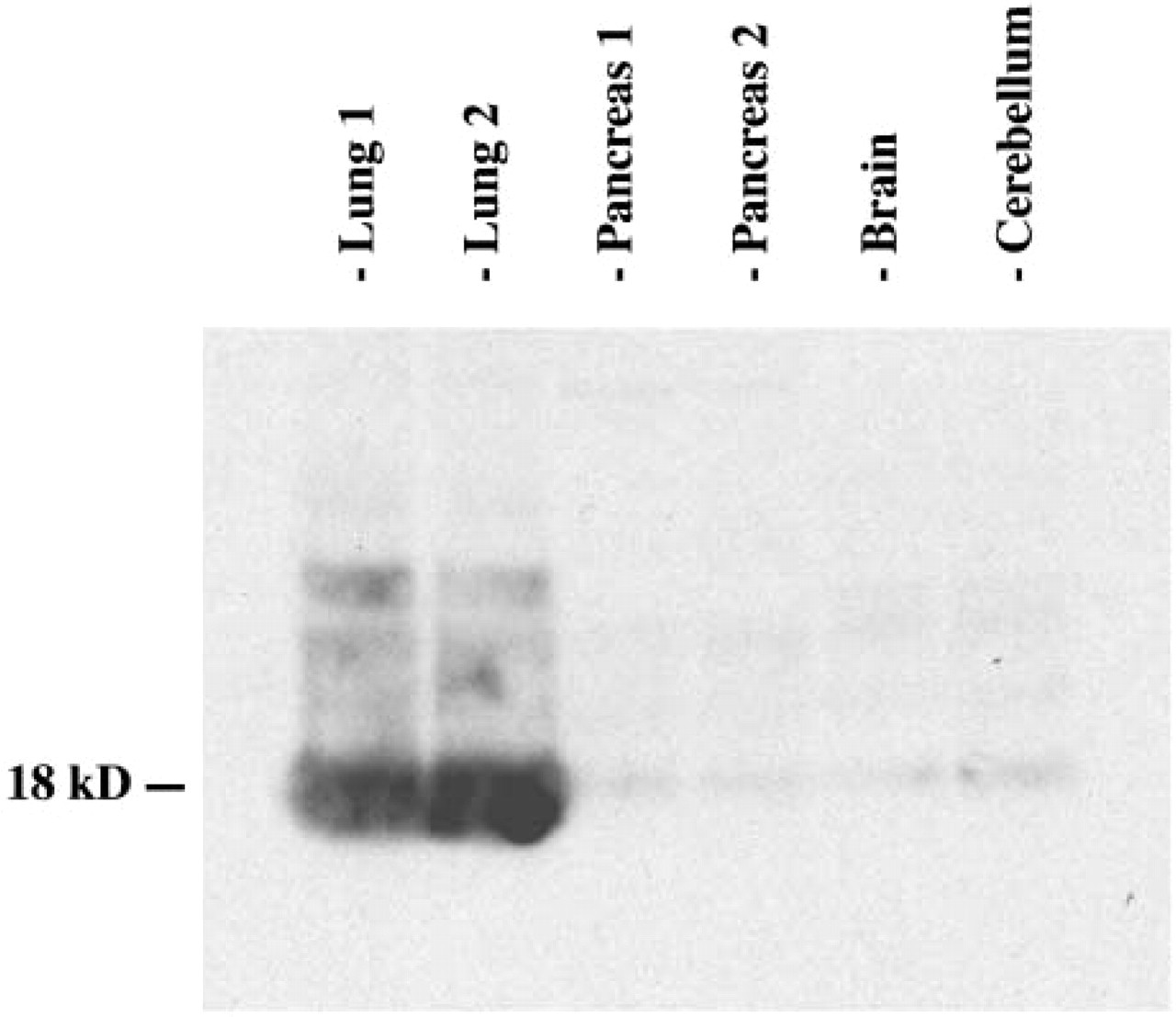
Western blotting analysis, showing the expression of adrenomedullin (AM) in pigeon tissue extracts. For lung and pancreas, extracts from two different animals are included to demonstrate reproducibility. Molecular weight of the immunoreactive band is indicated on the left.
To identify the cell type responsible for the production of AM in avian lung, immunohistochemical methods were used. The histology of avian lung, having type I pneumocytes restricted to capillary airways and type II pneumocytes covering the atrial and parabronchial surfaces, makes it easy to localize both cell types in paraffin sections (for a detailed description of the histology of avian lung, see López 1995). As shown in Figure 3, AM-like immunoreactivity was found mainly in the cytoplasm of type II pneumocytes (Figures 3A, 3E, and 3F), but there was also immunoreactivity in mesobronchial cells (Figure 3B), chondrocytes, and atrial macrophages (results not shown). The specificity of the staining was proven by absorption controls, which resulted in a total quenching of the immunolabeling (Figure 3C).
Electron microscopy immunostaining was applied to investigate the subcellular localization of the AM-like peptide: colloidal gold particles were located mainly in lamellar structures of the multivesicular system or associated with lamellar bodies (Figure 3D) and occasionally in vacuoles of type II pneumocytes (data not shown). No gold particles were found outside this location with the exception of pulmonary macrophages, where some gold particles were also found in the cytoplasm (data not shown).
AM-like Peptide Production in Response to Air Pollutants
To further understand the role of AM in pulmonary type II cells, we designed two sets of experiments, seeking for differences in the production and/or distribution of AM after exposure to air pollutants. The first approach consisted of a comparative study between pigeons raised in the polluted atmosphere of a large city and their counterparts raised in the countryside. When comparing the intensity of the immunostaining in these two sets of animals (Figures 3E and 3F), the immunolabeling for AM was stronger in city pigeons than in countryside pigeons. Densitometric analysis showed that the AM IOD ratio in countryside pigeons was significantly lower than the ratio found in the group of city pigeons (Figure 4).
AM-like Peptide Production in Response to Ozone
In agreement with the previous results in the comparative study, the intensity of AM-like immunostaining was significantly higher in the cytoplasm of type II pneumocytes from animals exposed to higher ozone concentrations (Figure 5). These differences are statistically significant between the control group and the two groups exposed to ozone (p<0.05), with no perceptible difference in the intensity of the immunostaining between the two subsets of animals exposed to ozone.
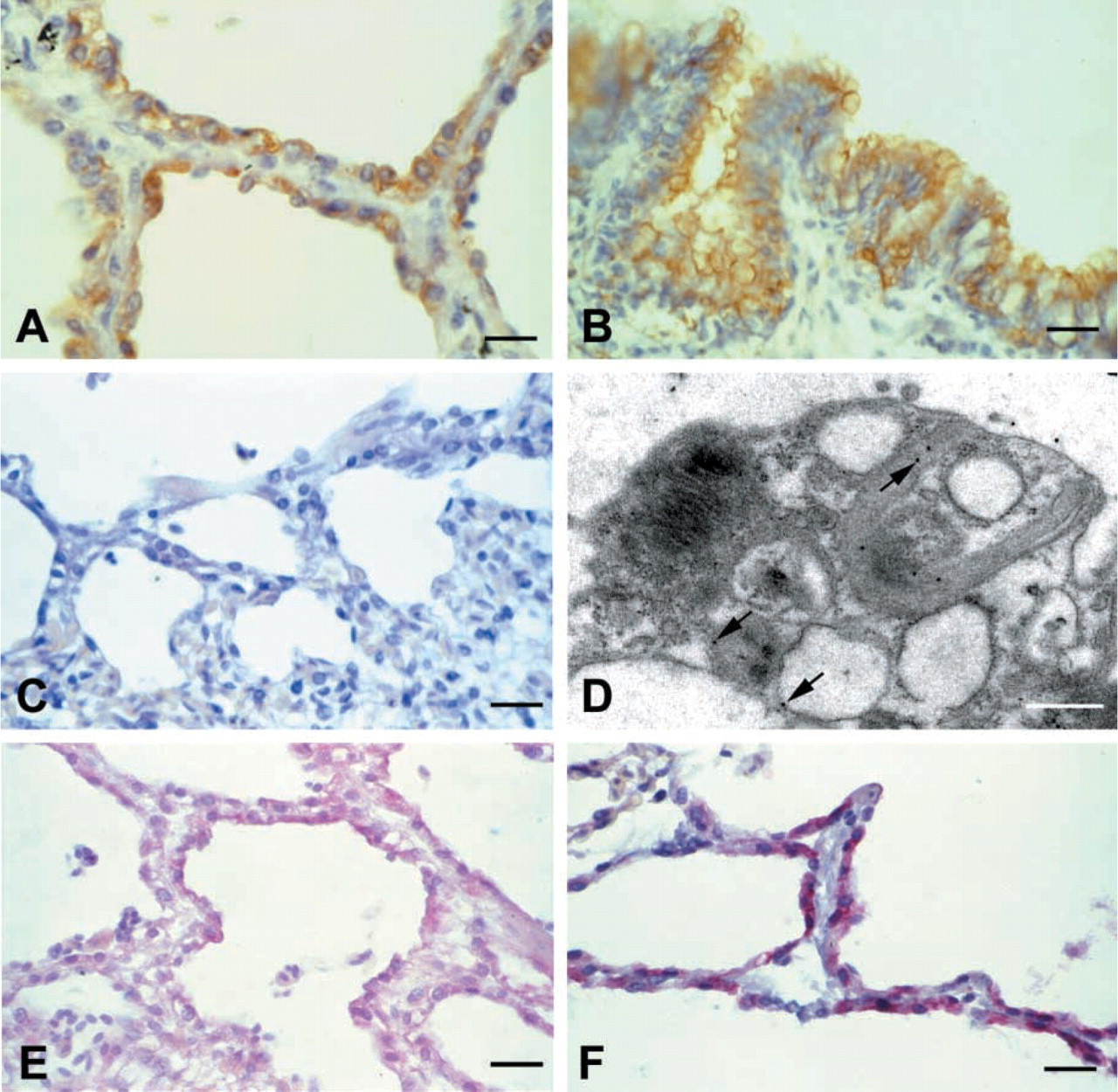
Distribution of AM immunoreactivity in the pigeon lung as seen by immunohistochemical methods.
Discussion
This study presents the first immunohistochemical description of an AM-like protein in the pulmonary surfactant system of a non-mammalian vertebrate species. The antibody used was raised against the human peptide but has been used successfully in previous studies in non-mammalian species including birds (López et al. 1999a,b), supporting that AM is well preserved throughout the evolution of vertebrates. The specificity of the immunostaining has been demonstrated by Western blots and absorption controls.
We have shown that AM-like immunoreactivity is expressed in the lung of Columba livia mainly by type II pneumocytes, atrial macrophages, and mesobronchial epithelial cells. This distribution is similar to the pattern described for mammals (Martínez et al. 1995), with the exception of type II pneumocytes. In rat type II pneumocytes, only AM mRNA has been detected (Okumura et al. 2000). The presence of the peptide has not been demonstrated by immunocytochemical methods, probably because this peptide is rapidly secreted, making the concentration of AM in the cytoplasm too low to allow for detection with these methods. In pigeons, however, the amount of intracellular AM-like peptide seems to be sufficient for immunohistochemical detection. If confirmed in other avian species, this difference could be relevant from a phylogenetic point of view.
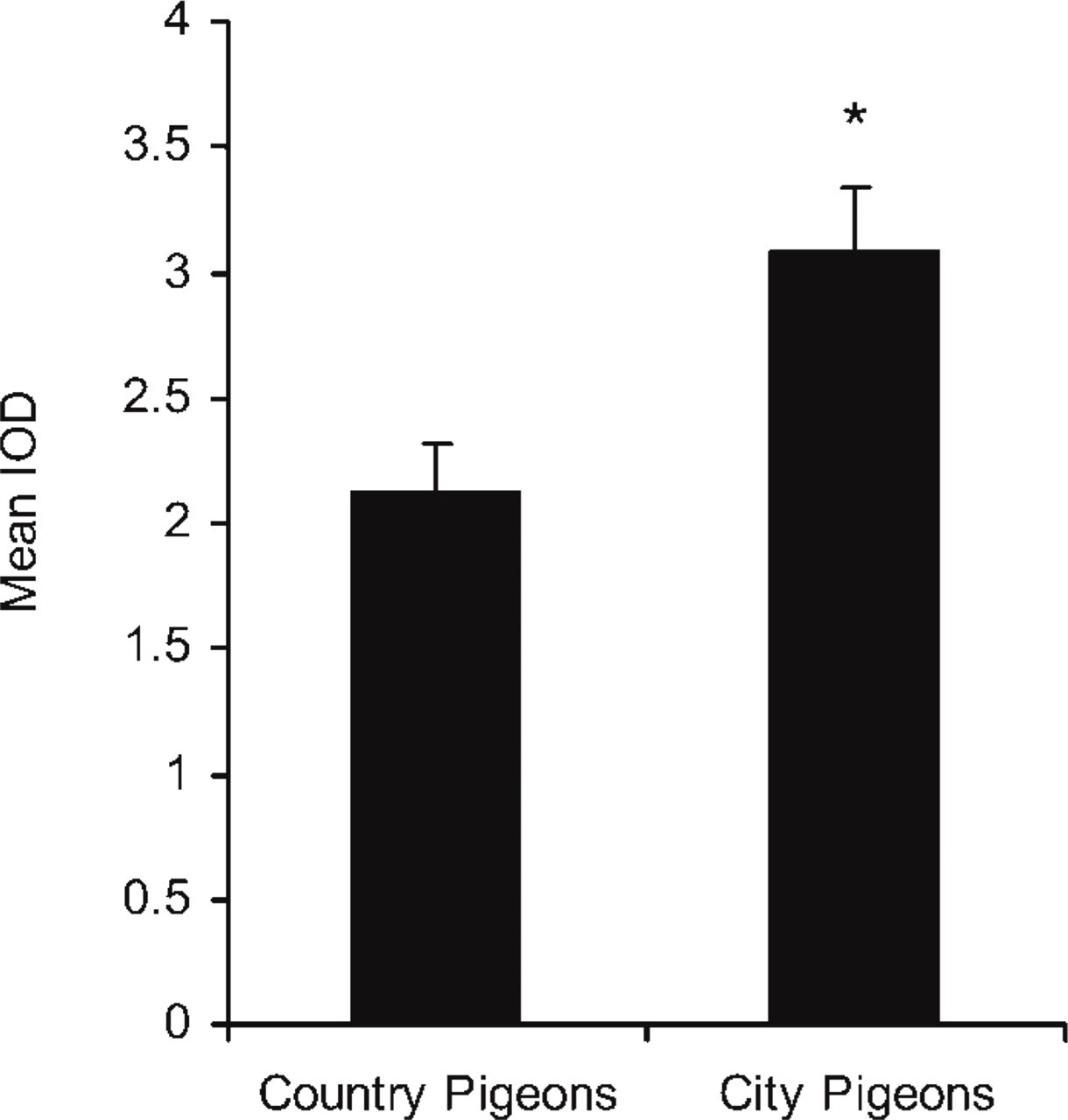
Quantitative analysis of the expression of AM in countryside and city pigeons. The relative immunoreactivity for AM in countryside and city pigeons was measured by densitometric analysis integrated optical density (IOD). (IOD ratio: 2.12 ± 0.2 vs 3.09 ± 0.28; ∗ p<0.05, n= 6).
At the subcellular level, we have detected the presence of AM-like immunoreactivity in membrane structures of the multivesicular system and in lamellar bodies of type II pneumocytes. This observation implies that AM may be stored and secreted together with the components of pulmonary surfactant, as do other molecules such as vitamin E (Rüstow et al. 1993) or lysozyme (Beers et al. 1994). We have also detected immunogold particles linked to the AM antibody in vacuoles of type II pneumocytes. This fact suggests that AM could be secreted to the extracellular milieu by an alternative mechanism independent of the excretion of lamellar bodies. Its presence in atrial macrophages and mesobronchial cells suggests a role as a protective peptide, participating in the prevention of local infection, as some authors have previously proposed (Martínez et al. 1995,1997; Allaker et al. 1999). In this sense, the simultaneous secretion of AM and pulmonary surfactant may add an important physiological value to the surfactant components, namely, antimicrobial activity. Such localization might also point to a possible role in the regulation of surfactant secretion. It has been previously demonstrated that AM increases phosphatidylcholine secretion (the main component of pulmonary surfactant) in rat type II pneumocytes (Okumura et al. 2000). These results, together with the subcellular localization of AM in lamellar bodies, suggest a possible role for this peptide as an autocrine/paracrine modulator of surfactant secretion.
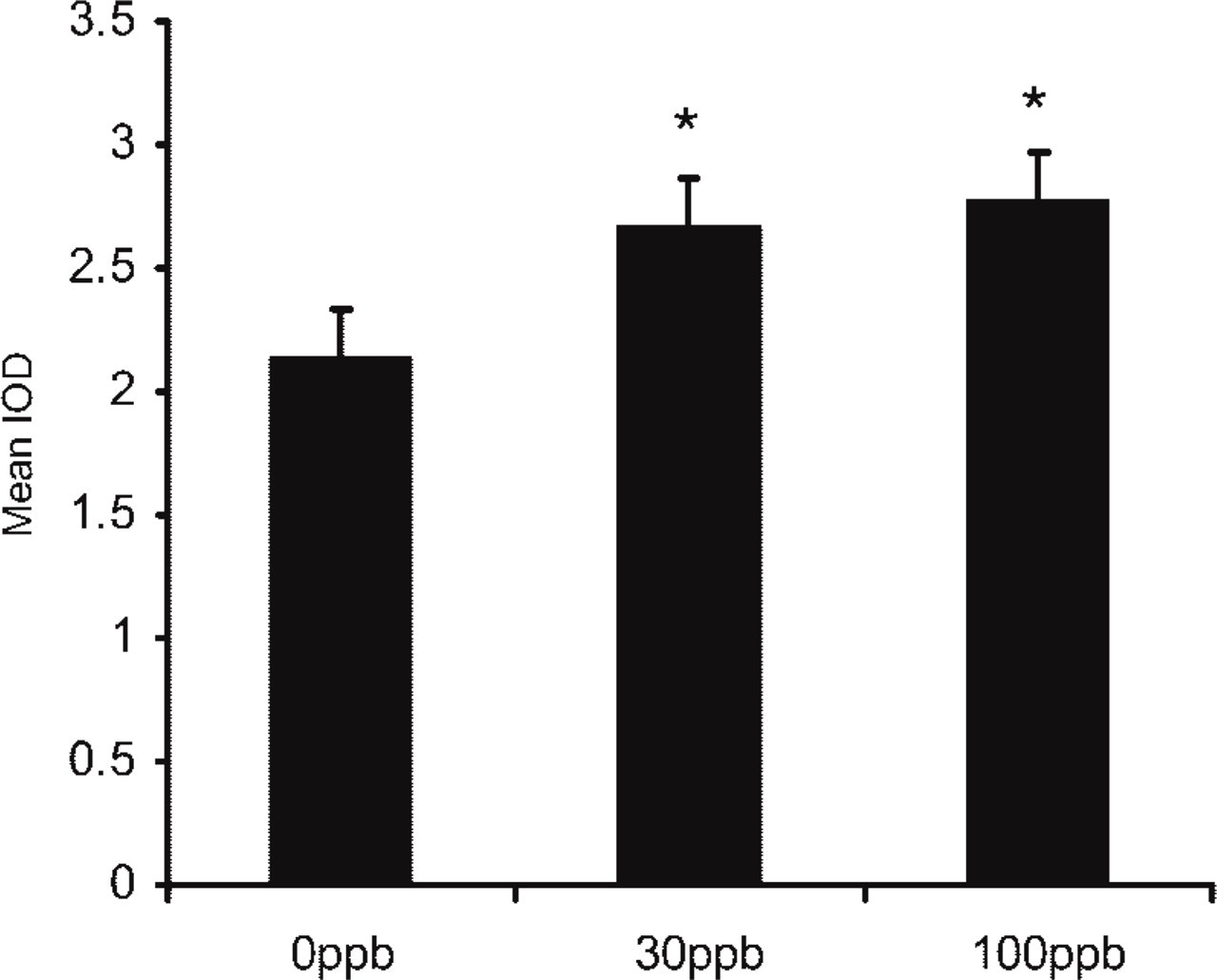
Quantitative analysis of the expression of AM in pigeons exposed to different ozone concentrations for 5 days. The IOD ratio in the non-exposed group was 2.14 ± 0.03 vs 2.66 ± 0.06 and 2.77 ± 0.07 in the exposed groups (30 ppb and 100 ppb ozone, respectively) (∗ p<0.05, n = 2).
Our results show that AM-like immunoreactivity is significantly elevated after either chronic or acute exposure to air pollutants. This observation suggests that AM expression may be activated by lung injury. It has been shown previously that pulmonary AM levels are significantly elevated in response to stress situations such as hypoxia and acute restrain (Khan et al. 1999; Matsui et al. 2004). Under such conditions, pulmonary surfactant components present alterations in their synthesis and excretion rates (Hallman et al. 1982; van Golde et al. 1988). Previous studies showed that the relative amount of lamellar bodies is reduced in city pigeons when compared with countryside birds (Lorz and López 1997). These results, together with our recent findings of a relative increase in AM-like peptide levels in type II pneumocytes of city pigeons, allow us to hypothesize that AM synthesis might be activated to maintain constant levels of this peptide in the atrial space.
The relaxation of the vascular bed is another physiological role attributed to AM (Nossaman et al. 1995). An increase in AM production in response to air pollutants, as well as the production of other vasodilatory peptides, would allow some nutrients to more easily reach the damaged cells.
As we pointed out previously, studying the effects of combined exposures of pollutants is essential for health effect predictions but describing the effects of single pollutants on the pulmonary surfactant system is necessary as well. For this purpose, we chose an experimental model to study the effects of a common oxidant, ozone, present in urban atmosphere. In the majority of studies involving ozone as a single atmospheric pollutant, the concentration used is much higher than that found in polluted air (Balis et al. 1991; Putman et al. 1995,1997; Currie 1998). The concentrations of ozone analyzed in our study were situated within the range of “permitted” levels established by the European Community Legislation and provide a better understanding of the possible noxious effects to pulmonary cells of a polluted environment. Under these experimental conditions we could detect statistically significant changes in AM-like immunoreactivity following short-term exposure to ozone concentrations as low as 30 ppb. In agreement with the comparative study between countryside and city pigeons, AM-like peptide expression is augmented after exposure to a single air pollutant. As some studies have reported, there is an increase in AM secretion after oxidative stress in endothelial cells (Chun et al. 2000; Saito et al. 2001), suggesting that AM could be a protective peptide when an oxidative injury occurs. This antioxidant effect, if proven in type II pneumocytes, would explain the augmented AM-like immunoreactivity after ozone exposure.
To summarize, our study demonstrates the presence of an AM-like peptide in type II pneumocytes of the avian lung. Long-term exposure to poor air quality, even at levels within current standards, results in increased AM-like immunoreactivity in type II pneumocytes. AM increased expression and secretion in response to air pollutants might suggest a protective role for this peptide in the pulmonary epithelium and a potential contribution to surfactant secretion regulation.
Footnotes
Acknowledgements
Supported partially by grants from Comunidad Autónoma de Madrid and Caja de Madrid Foundation to NC.
We are grateful to the Atmospheric Pollution Department of the Madrid Council and to the Electron Microscopy Service of the Universidad Autónoma de Madrid for their help. We wish to thank Dr. M.J. Sanz and Centro de Estudios Ambientales del Mediterráneo (Valencia, Spain) for letting us use the open top chambers located in their installations.
This study is dedicated to the memory of the late Prof. José López Diez del Corral, under whose supervision it was performed at the Department of Biology, Universidad Autónoma de Madrid.