
Abstract
Adrenomedullin (AM) is a multifunctional regulatory peptide that stimulates cyclic AMP production in many target tissues and is highly expressed in the lung. Analysis of the distribution of the recently cloned AM receptor (AM-R) by non-radioactive in situ hybridization revealed abundant expression in the basal cells of the airway epithelium and Type II pneumocytes. The expression of AM-R in the two cell types involved in epithelial regeneration of the lung suggests that AM may be relevant in such functions as organ development, wound repair, and epithelial turnover. AM-Rs are also synthesized in vivo and in vitro by a variety of tumor cells that also express the ligand, suggesting the existence of an autocrine loop that may be involved in tumor growth stimulation. The present findings suggest that the AM/AM-R regulatory system plays a major role in respiratory physiology and lung carcinogenesis and that new functions for AM remain to be identified.
A
The biological effects of AM are mediated by elevating cellular cAMP levels through a G-protein-coupled mechanism (Ishizaka et al. 1994). A specific receptor for AM (AM-R) has recently been cloned and sequenced (Kapas et al. 1995). It contains seven transmembrane domains and has a general structural resemblance to other members of the G-protein-linked receptor superfamily. The receptor binds AM at nanomolar concentrations and does not recognize CGRP, although AM appears to be able to activate CGRP receptors with lower affinity (Eguchi et al. 1994).
Expression of the AM-R is particularly high in lung, in which Northern blot analysis reveals several mRNA species, of which the most prominent is a 1.8-KB transcript (Kapas et al. 1995). We have used in situ hybridization to determine the distribution of AM-R in human lung for both normal and malignant settings, as a first step in the understanding of AM/AM-R functions in the lung.
Materials and Methods
Tissues
Ten normal lung specimens from patients who died without pulmonary involvement were provided by the Department of Pharmacology, University of Pittsburgh (protocol M1259). Eighteen additional formalin-fixed, paraffin-embedded blocks containing representative cases of lung tumors (Table 1) were obtained from the files of the NCI-Navy Medical Oncology Branch, National Naval Medical Center, Bethesda (protocol 83-15).
Results obtained by in situ hybridization in surgical specimens representing the different major neoplasias of the lung
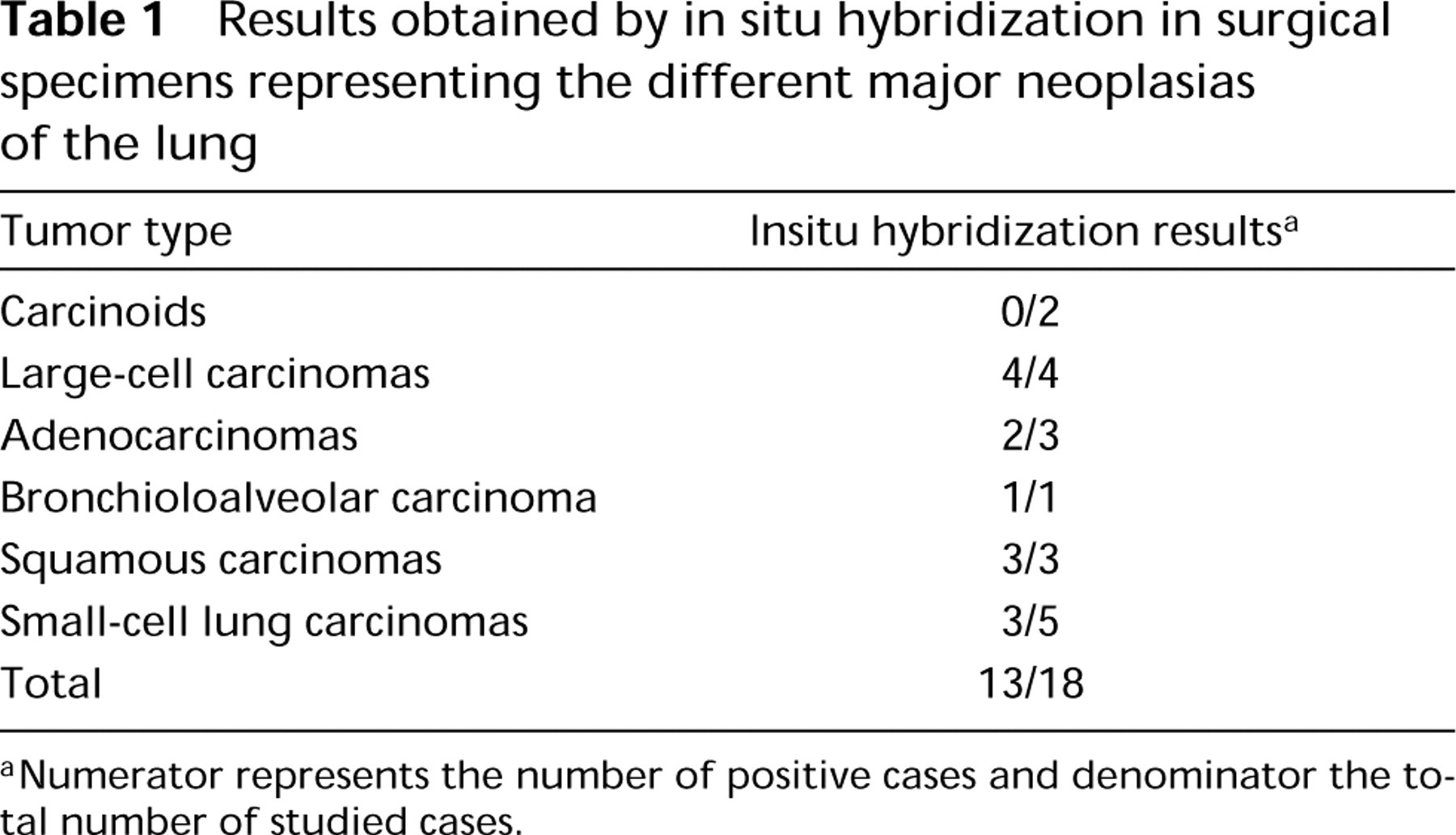
aNumerator represents the number of positive cases and denominator the total number of studied cases.
Cell Culture
Twenty-six lung tumor cell lines were used in this study: NCI-H727, H679, H835, UMC11 (carcinoids), H23, H726, H2087, H1373, H2228, H920 (adenocarcinomas), H1404, H820, A549 (bronchioalveolar carcinomas), H520 (squamous carcinoma), H82, H446, N417, H187, H345, H209, H510, H889, H774, H123, H735, and N592 (small-cell lung carcinomas). Cells were grown in serum-free, hormonefree medium, as previously described (Siegfried et al. 1994). mRNA was extracted from fresh cell pellets using the isothiocyanate-cesium chloride method (Glisin et al. 1974). Polyadenylated RNA from normal human lung was purchased from Clontech Laboratories (Palo Alto, CA).
RT-PCR
Three oligonucleotides were selected via DNASTAR software analysis of rat AM-R cDNA (Table 2) and were used as primers/probe for PCR and Southern blot experiments. An expected PCR product of 471 BP was indicative of AM-R message expression. Respective oligonucleotides were synthesized by Midland Certified Reagents (Midland, TX).
Reverse transcription was performed using the Super-Script Preamplification System (Life Technologies; Gaithersburg, MD) per the manufacturers protocol. A Perkin-Elmer 9600 thermocycler was used in amplifying the gene products in the samples. All buffers, enzymes and nucleotides used were obtained from Applied Biosystems (Perkin-Elmer Cetus; Norwalk, CT). PCR products were analyzed electro-phoretically using 1% agarose gels, and after ethidium bromide staining were visualized under UV light, followed by Southern analysis. Two PCR products (total human lung and cell line NCI-H720) obtained using the same primers have been previously cloned and sequenced, obtaining a high homology with the original rat clone (Miller et al. 1996).
Sequences of the oligonucleotides synthesized for this study

Southern Blot Analysis
Gels were denatured in 1.5 M NaCl/0.6 M NaOH and 1.5 M NaCl/2 M Tris and DNA transferred overnight onto 0.2-μm nitrocellulose filters in 20 X SSC by standard capillary flow techniques. DNA was crosslinked to the filter at 80C under vacuum and incubated in hybridization buffer. The anti-sense nested probe was 32P-end-labeled by standard procedures. Hybridization with the probe was done overnight at 42C. Room temperature stringency washes were performed using 5 X SSC/0.1 % SDS and 1 X SSC/0.1 SDS. Filters were air-dried and 32P signal resolved by autoradiography at −80C, using Kodak XAR5 film.
In Situ Hybridization
Detection of the AM-R mRNA was performed using in situ hybridization as previously described (Martínez et al. 1996). The full-length cDNA was ligated into the expression vector pcDNA1 (Kapas et al. 1995) and used to generate riboprobes. The plasmid was linearized with EcoRV and BamHI and was used as a template to synthesize digoxigenin-labeled sense and anti-sense RNA transcripts. Hybridization was performed in a moist chamber at 46C for 20 hr in a 20-μl volume containing 2.5 ng of probe/μl. After stringency washes, visualization of hybridized probes was performed using a digoxigenin detection kit (Boehringer Mannheim; Indianapolis, IN). Controls included the use of the sense probe and digestion with RNAse before hybridization.
Immunocytochemistry
In selected slides, immunocytochemistry for AM was performed after in situ hybridization for the receptor. As previously characterized rabbit anti-human AM antibody (Martínez et al. 1995) was used together with the Vectastain ABC kit (Vector Laboratories; Burlingame, CA) for ligand localization.
Results
Normal Lung
In the upper airways, AM-R mRNA was localized in the cytoplasm of the basal cells of the epithelium (Figure 1A). Control slides, treated with the sense probe, showed some nonspecific staining in discrete cells of the connective tissue (Figure 1B) but clearly demonstrated the specificity of the staining with the anti-sense probe in the other structures. Surprisingly, the smooth muscle bundles of both the airway walls and the blood vessels did not show any hybridization signal. Immunocytochemical staining for AM after in situ hybridization showed the previously described pattern (Martínez et al. 1995), being especially strong in the apical border of the epithelium and in the airway smooth muscle (Figures 1C and 1D). Other locations in which the AM-R was highly expressed included the cytoplasm of the serous cells of the bronchial glands and some inflamatory cells in their neighborhood (Figure 2). In the peripheral lung, AM-R was expressed by the Type II pneumocytes, which are characterized by their prominent cytoplasm in the alveolar lumen (Figure 3).
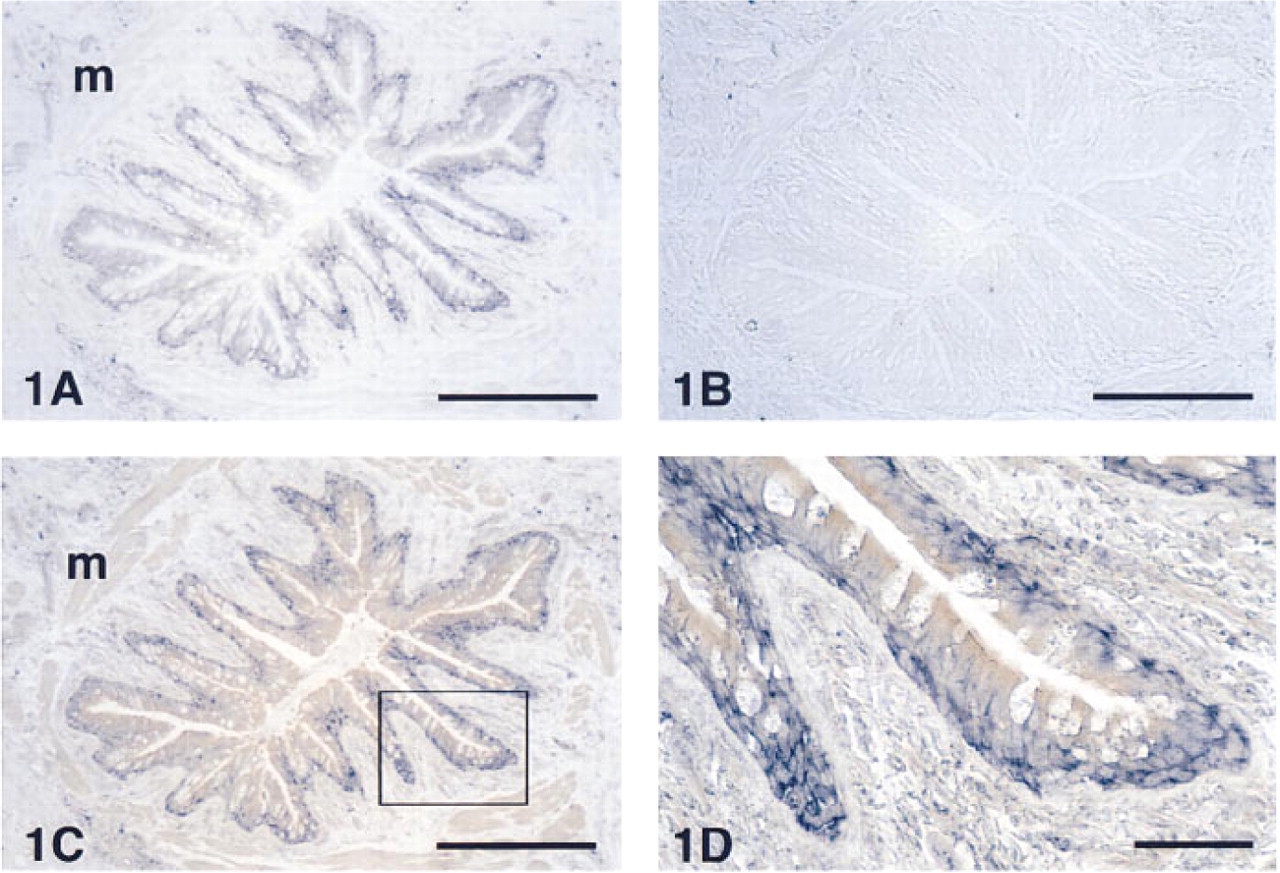
Localization of AM-R in a normal human bronchiolus after hybridization with anti-sense
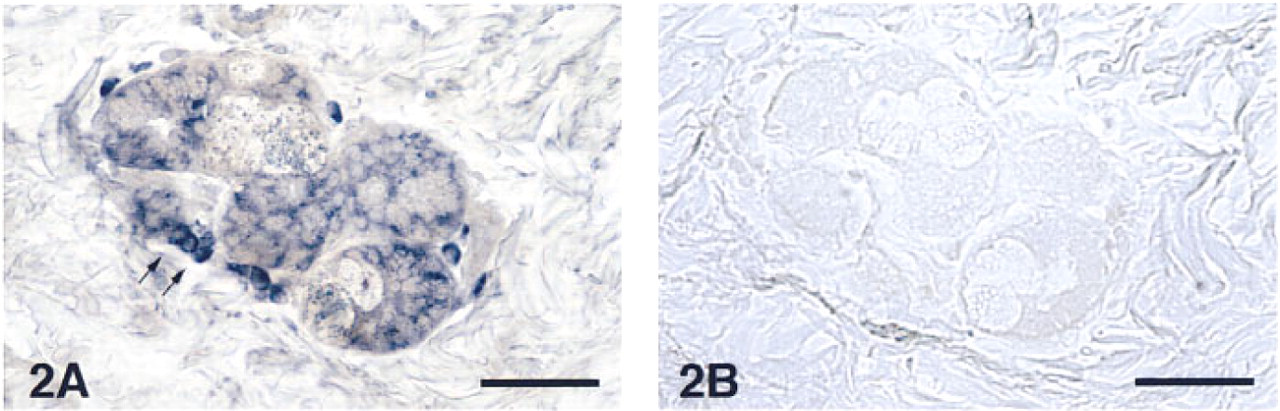
Section of a bronchial gland in a healthy human lung displaying an intense signal for AM-R expression in the serous cells and in some associated inflammatory cells (arrows). Bar = 40 μm.
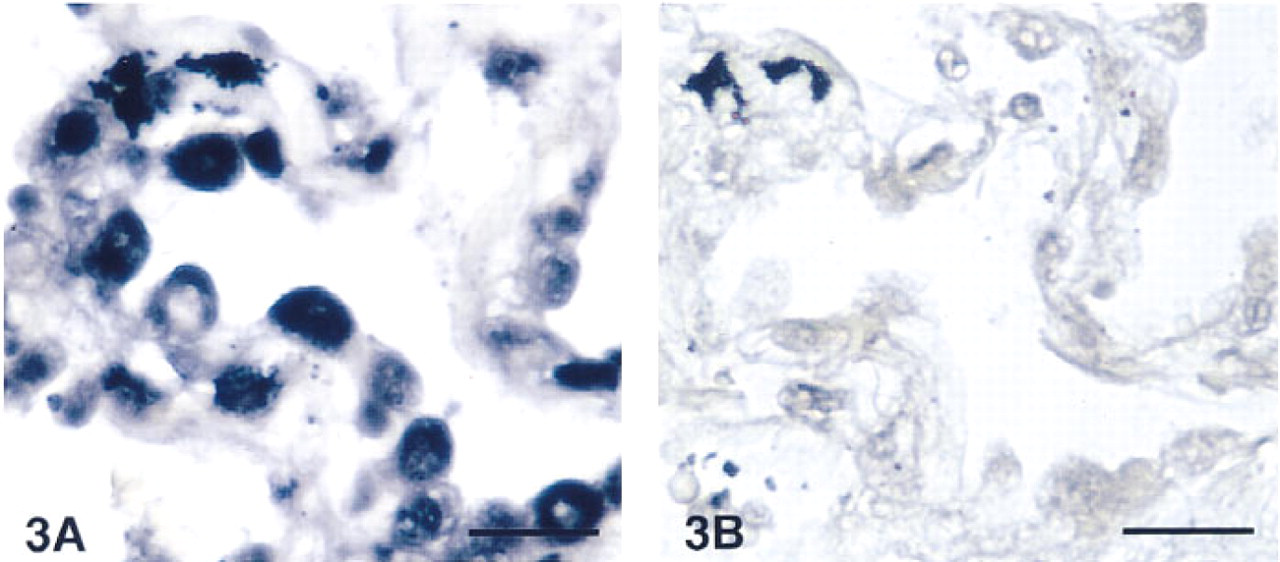
Detail of the alveolar region in a normal human lung. Type II pneumocytes were strongly stained with the anti-sense probe for the AM-R
Tumors
We studied 18 tumors of pulmonary origin with different diagnoses (Table 1). Of the 18 studied, 13 tumors (72%) were positive for AM-R by in situ hybridization. Some examples of these are shown in Figures 4 and 5. Analysis of the production of AM-R mRNA in tumor cell lines by RT-PCR revealed that 15 of the 26 cell lines (58%) expressed detectable amounts of mRNA, as demonstrated by Southern blot (Figure 6).
Discussion
We have shown, using in situ hybridization techniques, that AM-R is highly expressed in the lung, consistent with the results of the Northern blot data presented in the original description of the AM-R (Kapas et al. 1995). Taking into consideration the similarity between members of the G-protein-linked receptor superfamily, we used high-stringency conditions during hybridization (46C instead of the usual 42C) and subsequent washes to exclude possible cross-hybridization artifacts.
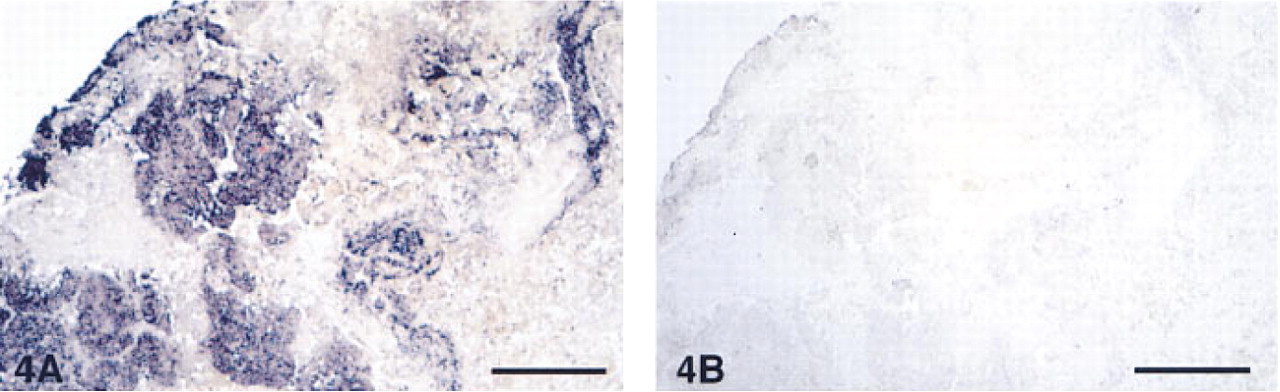
Panoramic view of a squamous carcinoma showing intense staining in the tumoral cells and not in the stroma. Bar = 200 μm.
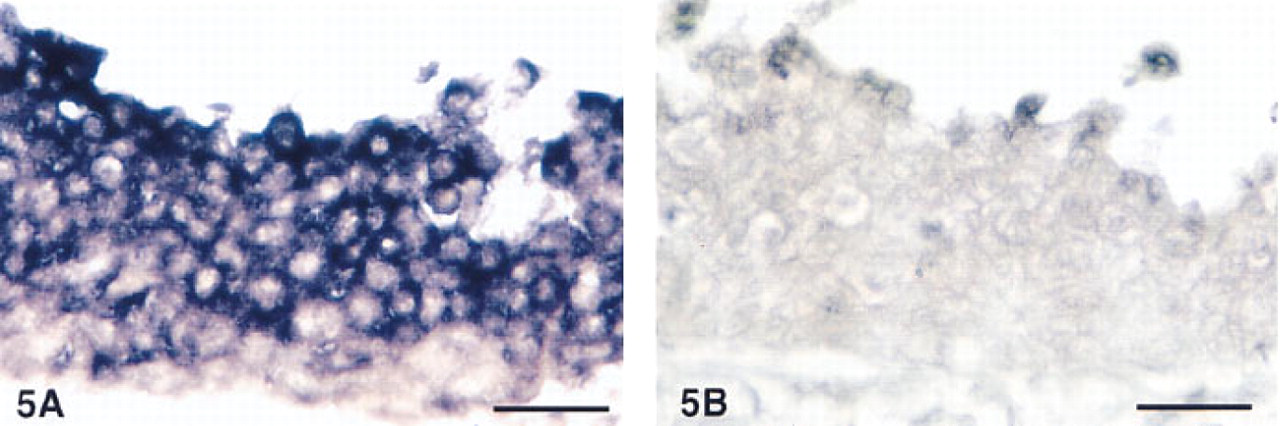
Detail of the tumor cells of a large cell carcinoma. The hybridization signal was restricted to the cytoplasm after hybridization with the anti-sense probe
We have previously described the distribution of AM in the normal and malignant human lung. AM is produced by the airway epithelium, bronchial glands, neurons, endothelial cells, chondrocytes, macrophages, and smooth muscle cells, and is present in many tumors (Martínez et al. 1995). This distribution coincides with the expression pattern of peptidyl-glycine α-amidating mono-oxygenase, the enzyme complex responsible for amidation of regulatory peptides (Saldise et al. 1996). It has been shown that AM requires this post-translational modification to bind efficiently to the receptor (Eguchi et al. 1994).
The expression of AM-R in basal cells and Type II pneumocytes, two cell types involved in regeneration of the epithelial layers of the lung (Keenan 1987; Plopper and Dungworth 1987), as well as in a variety of tumor cells, points to a role for AM as a growth factor. Therefore, it is possible that AM participates in organ development, normal turnover of the epithelium, wound repair, and tumor progression. Further evidence for this function was obtained in tumor cell lines, in which the application of a monoclonal antibody against AM drastically and reversibly reduced cell growth (Miller et al. 1996). Consistent with our findings, AM has been identified as a mitogenic factor for the mouse fibroblast cell line Swiss 3T3, which mediates its trophic actions through an AM-specific receptor system insensitive to CGRP (Withers et al. 1996).
The specific distribution of AM and its receptor in the airway epithelium suggests a new hormonal mechanism of action that might be important in the rapid response to tissue injury. AM is stored in the apical end of the ciliated cells and is secreted into the lumen of the organ, where it acts as an antimicrobial agent (Tom Walsh, Pediatric Oncology Branch, NCI, personal communication). Ligand-specific receptors, on the other hand, are located in the basal region of the epithelium and are separated from this source of AM by the intact epithelial layer. If an epithelial injury occurs (infection, chemical or mechanical damage), cell boundaries are disrupted and the luminal pool of AM becomes available for receptor binding. Given the mitogenic potential of AM, this results in a rapid proliferative response which eventually heals the wound, restoring epithelial integrity and re-separating the ligand from the receptor. The term “traumakines” would be appropriate for peptides displaying such behavior, with AM as the first identified member of this family. During normal growth conditions (epithelial turnover), the receptors could receive the ligand from mesenchymal components, such as smooth muscle cells, or from the interstitial fluid derived from the plasma, a recognized source of AM (Ichiki et al. 1994).
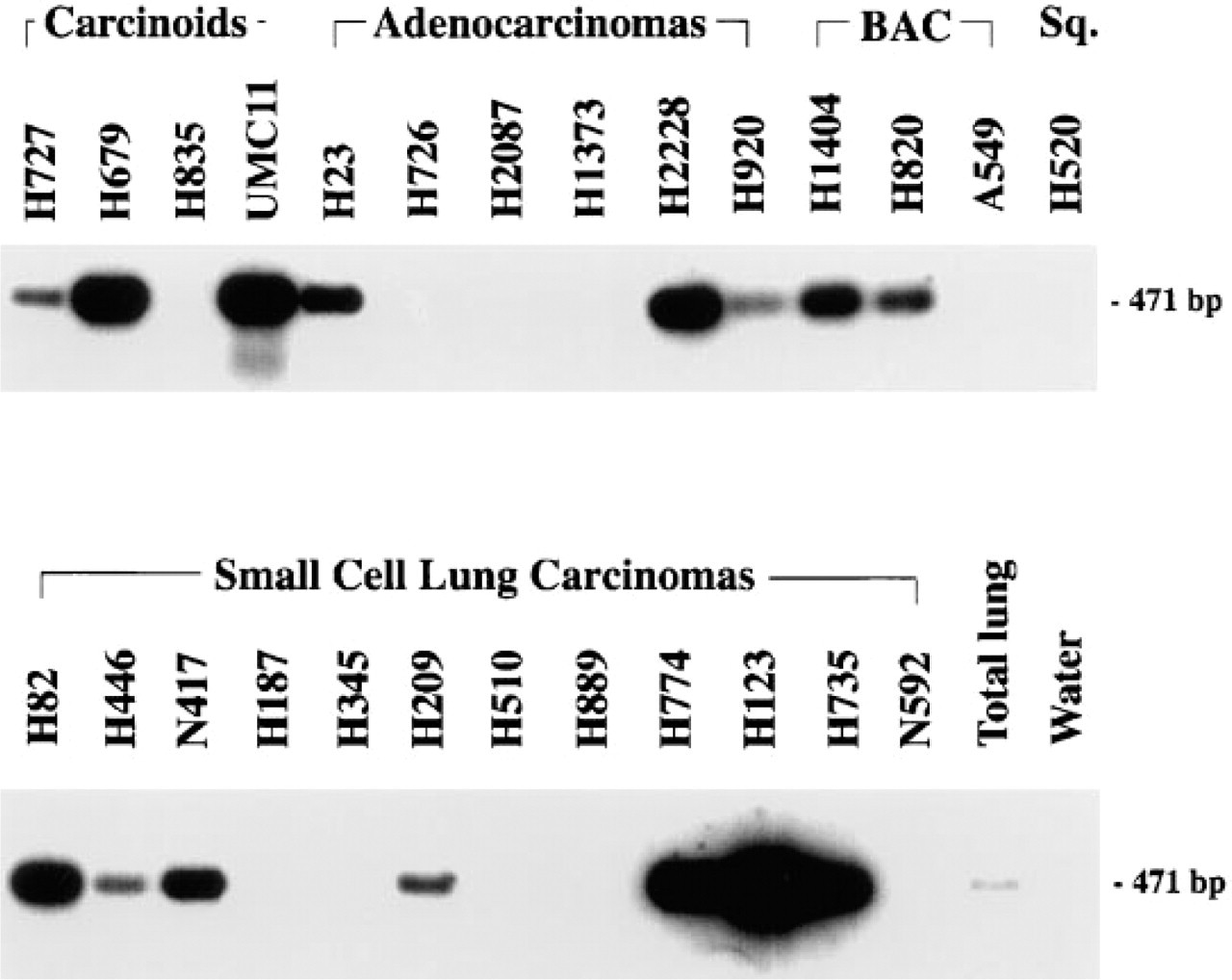
Southern blot analysis of 26 tumor cell lines representing the five major malignancies of the lung. Amplification of mRNA from normal human lung by RT-PCR using AM-R-specific primers yielded the expected 471-BP product.
The co-expression of both the ligand and the receptor in tumors suggests that AM and its receptor are implicated in an autocrine mechanism, that may regulate cell growth (Miller et al. 1996). Several such autocrine growth factors have been identified (Cuttitta et al. 1993) and their use as potential targets for pharmacological intervention has been proposed (Mulshine et al. 1991).
An unexpected finding was the absence of a hybridization signal in the smooth muscle cells of the blood vessels and the bronchial walls (Figures 1A and 1C). Smooth muscle cells are believed to express AM-R that mediate the vasodilatatory and bronchorelaxant responses to AM. There are several reports (e.g., Sugo et al. 1995; Eguchi et al. 1994) that describe the presence of AM-R in smooth muscle cells. However, these studies employed systems in which the CGRP receptor was also present. It is conceivable that some of these reported responses could have been mediated through a less selective receptor for AM. Pertinent to our findings, AM-R was isolated from a pulmonary library and was found to be maximally expressed in the lung (Kapas et al. 1995). It is possible that this receptor represents a pulmonary subtype and that additional receptor isoforms for AM, including a specific vascular subtype, remain to be identified.
In conclusion, our data clearly demonstrate the expression of AM/AM-R in normal and malignant lung. The regional distribution of these molecules, along with previously reported data on their trophic action, implicates this ligand/receptor relationship as a possible growth regulatory mechanism involved with normal epithelial turnover, wound repair, and carcinogenesis.
Footnotes
Acknowledgements
Supported by the United States Department of Energy and the Los Alamos National Laboratory Flow Cytometry Resource (NIH grants p41-RR01315, RR06758, and R01 RR07855).