
Abstract
Microdissected rat proximal straight tubules (PST) and inner medullary collecting ducts (IMCD) highly produce urea from L-arginine, supporting the expression of the mitochondrial arginase II. However, IMCD contain a very low density of mitochondria compared with PST. Recently, arginase II has been localized by immunohistochemistry in rat PST but not IMCD. This study was designed to verify whether rat IMCD express arginase II and to identify its subcellular localization. We developed an antibody raised against arginase II that allowed the detection of a band of 38 kDa corresponding to arginase II on immunoblots. In male and female rat kidneys, Western blot analyses revealed that arginase II was highly expressed in the inner medulla (IM), the outer stripe of the outer medulla (osOM), and the deep cortex. Immunocytochemistry demonstrated that arginase II was homogeneously expressed in IMCD. Proteins of the cytosolic and mitochondrial fractions extracted from osOM and IM and analyzed by Western blot showed that 86% of arginase II was associated with mitochondria. The molecular weight of arginase II was similar in the cytosolic and mitochondrial fractions. Immunoelectron microscopy confirmed the presence of arginase II in the mitochondria of IMCD. In conclusion, arginase II is expressed in mitochondria of male and female rat IMCD.
A
Arginase II gene is expressed in kidneys of various mammalian species. Significant amounts of arginase II mRNA and protein were detected in whole human (Morris et al. 1997), rat, and mouse (Gotoh et al. 1996; Iyer et al. 1998; Shi et al. 1998; Ozaki et al. 1999; Levillain et al. 2005) kidneys, whereas arginase II activity was measured in rat (Skrzypek-Osiecka et al. 1983) and mouse (Kochakian and Stettner 1948; Bradley 1973; Manteuffel-Cymborowska et al. 1995) kidneys. In almost all the species studied including rodents, carnivores, omnivores, and lagomorphs, the renal localization of arginase II exhibits similarity and differences (Robinson and Schmidt-Nielsen 1963; Levillain and Hus-Citharel 1996; Levillain et al. 2005). The precise localization of arginase II in the mammalian kidney has been achieved by different experimental approaches. Several years ago we indirectly localized arginase II activity by measuring the production of 14C-urea from viable microdissected tubules incubated in the presence of
Transcription of arginase II mRNA leads to a single polypeptide of 354 amino acid residues including a putative NH2-terminal presequence for mitochondrial import (Shi et al. 1998). This arginase II precursor has a molecular weight of ~40 kDa, whereas the mature form of arginase II, which is localized within the mitochondria, exhibited a molecular weight of ~38 kDa (Gotoh et al. 1996; Levillain et al. 2005). The mitochondrial localization of arginase II in the murine OSPST has recently been demonstrated by using a specific and purified antibody raised against arginase II (Levillain et al. 2005). Interestingly, the relative abundance of mitochondria in the rat PST is clearly several times higher than in IMCD (Pfaller 1982). Regarding the high rate of
The present paper was designed to determine whether the male and female rat IMCD express the arginase II gene and to identify the subcellular localization of the mature arginase II in this nephron segment. To achieve these goals, we developed a specific antibody raised against arginase II that differs from the antibody used by Miyanaka et al. (1998). Soluble proteins were extracted from the main renal zones dissected from the male and female rat kidneys and were analyzed on immunoblots to detect arginase II. Immunofluorescence and immunoelectron microscopy were performed to localize arginase II at the tubular and subcellular levels in dissected inner medulla (IM). In addition, to estimate the relative abundance of arginase II in the cytosol and mitochondria, the soluble proteins of each subcellular compartment were extracted from dissected outer stripe of the outer medulla (osOM) and IM, and analyzed on immunoblots. This experiment also allowed verification of the molecular weight of arginase II in each subcellular compartment.
Our results clearly show that in the male and female rat kidneys our antibody against arginase II allowed the detection on immunoblots of a single peptide of 38 kDa that corresponds to the expected molecular weight of arginase II. The level of arginase II was extremely abundant in all cells of the male and female IMCD and was almost exclusively localized in their mitochondria.
Materials and Methods
Animals
Six-week-old male (236 ± 4 g body weight, BW) and female (168 ± 6 g BW) Sprague Dawley rats from Iffa Credo (L'Arbresle sur Orge, France) had free access to tap water and standard laboratory diet (Souffirat; 20% protein, Genthon, France). Eight-week-old female Sprague Dawley rats (211 ± 3 g BW) were used for subcellular fractionation. Animals were housed in a room maintained at 20C with a 12:12-hr light/dark cycle. Rats were anesthetized intraperitoneally using 0.1 ml/100 g BW pentobarbital sodium (Nembutal 6%; Clin Midy, Paris, France). The experiments were approved by the local commitee for animal experiments.
Arginase II Antibody
A peptide-derived polyclonal antibody raised against arginase II was prepared by CovalAb (Lyon, France) and used for immunoblotting, immunohistochemistry, and immunoelectron microscopy. To obtain this polyclonal antibody, the conserved amino acid sequence (IASSFGQTREGGHIEC) of the human (Swiss-Prot P78540), mouse (Swiss-Prot O08691), and rat (Swiss-Prot O08701) arginase II was used to produce a synthetic peptide that differs from that of Miyanaka et al. (1998). The synthetic peptide was analyzed by HPLC and conjugated to keyhole limpet hemocyanin via covalent linkage to cysteine. One female New Zealand rabbit was immunized three times with the conjugate using a complete Freund's adjuvant. Arginase II antibody titer was determined by ELISA and immunopurified using a column on which the synthetic peptide was immobilized.
Dissection of the Main Renal Zones
The right and left kidneys of male and female rats were rapidly removed, decapsulated, and cut along the cortico-papillary axis to dissect the six main renal zones [superficial cortex (Cs), deep cortex (Cd), outer stripe of the outer medulla (osOM), inner stripe of the outer medulla (isOM), inner medulla (IM), and papilla (Pap)] at 4C under a stereomicroscope. The dissected tissue was immediately placed in a sterilized Eppendorf tube, frozen, and conserved in liquid nitrogen.
Protein Extraction and Western Blot Analyses
Each frozen renal zone was mixed at 4C with a Turrax homogenizer in the proportion of 100 mg frozen tissue per 2 ml of lysing buffer (Laemmli 1970) containing 1 mM of protease inhibitor cocktail, 1 mM phenylmethylsulfonyl fluoride, and 1 mM benzamidine. The homogenate was centrifuged at 10,000 x g for 30 min at 4C. The concentration of the soluble proteins was determined using the Bradford protein assay (Bradford 1976).
Fifty μg of soluble proteins from each renal zone were subjected to 10% SDS-PAGE and then transferred to a polyvinylidene difluoride membrane (0.45 μm; Immobilon-P, Millipore, St. Quentin en Yvelines, France) as previously described (Levillain et al. 2005). Proteins were visualized on the membrane with Ponceau S solution to control protein loading and transfer. Membranes were washed twice in TBST (20 mM Tris, pH 7.6, 137 mM NaCl, 0.1% Tween 20) and immersed twice in a blocking solution consisting of 5% fat-free milk powder in TBST for 30 min. Membranes were incubated separately with the three following rabbit polyclonal antibodies raised against arginase II (see above, dilution 1:1,000), the rat-aldose reductase (AR; dilution 1:3,000), and the rat cysteine sulfinate decarboxylase (CSD; dilution 1:1,000) diluted in 5% fat-free milk-TBST. Immunoblots were washed three times for 10 min in TBST and incubated for 60 min with a goat anti-rabbit IgG peroxidase-conjugated diluted 1:10,000 in 5% fat-free milk-TBST. Immunoblots were washed three times for 10 min in TBST, and antibody binding was revealed using chemiluminescence (ECL) Western Blotting Kit. ECL detection was performed using Kodak X-MAT film. Low-exposure film was scanned, and the intensity optical densitometry (IOD) of the bands was estimated using the ImagerMaster Total Lab v2.01 program (Pharmacia; Orsay, France).
Subcellular Fractionation for Immunoblot Analyses
The kidneys of four female rats were dissected at 4C under a stereomicroscope to collect their osOM and IM. The dissected tissue was rapidly immersed in a buffer (250 mM sucrose, 10 mM Tris-HCl, pH 7.6, and 1 mM Na2 EDTA) prealably maintained at 4C. The tissue was homogenized using a Potter-Elvehjem glass homogenizer PTFE pestle (Bellco Glass Inc.; Vineland, NJ) and centrifuged at 600 x g for 10 min to remove cell debris and the nuclear fraction. The pellet (P1) was discarded whereas the supernatant (S1) was centrifuged at 9000 x g for 10 min. The pellet (P2) that contained mitochondria was resuspended once in the same buffer and centrifuged again at 9000 x g for 10 min. The supernatant (S2) was centrifuged at 128,000 x g for 45 min (Optima TLA 100.2 Ultracentrifuge; Beckman) to obtain proteins of the cytosolic fraction in the third supernatant. Protein concentrations were determined using the Bradford protein assay (Bradford 1976).
Forty-μg proteins from the cytosolic and mitochondrial fractions were subjected to 10% SDS-PAGE and transferred for Western blot analyses as described above. Membranes were incubated with the rabbit polyclonal antibodies raised against the α-subunit F1-ATP synthase (dilution 1:5000) (Moradi-Ameli and Godinot 1983), core 2 (mitochondrial complex III core protein 2, dilution 1:20,000) (Legros et al. 2001), arginase II (see above), and the mouse monoclonal antibody raised against the iron sulfur protein (ISP, dilution 1:80,000) (Legros et al. 2001) diluted in 5% fat-free milk-TBST. For the next steps, immunoblots were incubated with a goat anti-rabbit IgG and a goat anti-mouse IgG peroxi-dase-conjugated diluted each 1:10,000 in 5% fat-free milk-TBST and treated as described above.
Indirect Immunofluorescence
Kidneys were rapidly removed, cut along the cortico-papillary axis, dipped in Bouin fixative, and embedded in paraffin. Tissue sections of 4-7 μm were collected on glass slides (ChemMate; DAKO, Trappes, France), deparaffinized three times in methylcyclohexane (toluene substitute) for 5 min at room temperature (RT), step-rehydrated in 100% ethanol for 5 min, twice in 95% ethanol for 5 min, rinsed in distilled water, and then in phosphate-buffered saline (PBS: 100 mM phosphate buffer, 150 mM NaCl, pH 7.4). Nonspecific sites were coated with PBS supplemented with 0.3% Triton X-100 and 1% BSA (PBST-BSA) for 120 min at RT and incubated overnight at 4C with our purified rabbit polyclonal antibody raised against arginase II (dilution 1:100) in PBST. The slides were rinsed three times in PBST for 5 min and incubated for 120 min at RT with goat anti-rabbit IgG Alexa Fluor 546-conjugated (dilution 1:1,000) (Interchim; Montluçon, France). Slides were washed twice for 5-10 min in PBST, one time in PBS for 5-10 min, and incubated with 4′,6-diamidine-2-phenylindole dihydrochloride (DAPI, 0.1 μg/ml) in PBS for 30 min at RT. Slides were washed three times in PBS and mounted with Fluoprep (bioMérieux; Marcy l'Etoile, France). Tissue sections were examined with a fluorescent microscope.
Immunoelectron Microscopy
Dissection of IM from male and female rat kidney was performed at 4C. The IM was cut into small pieces (<1 mm3) and fixed for 60 min at 4C in PLP composed of 10 mM metaperiodate, 75 mM lysine, and 2% paraformaldehyde in 37 mM Sorensen's phosphate buffer complemented with 0.2% glutaraldehyde. Pieces of tissues were washed three times in 50 mM Sorensen's phosphate buffer, 200 mM sucrose, and 100 mM lysine monochlorhydrate for 20 min at 4C, dehydrated through a series of washes with 30, 50, 70, and 95% ethanol maintained prealably at 4C for 5 min, and infiltrated with London Resin White (LRWhite; Euromedex, Mundolsheim, France) using 1:1 LRWhite and 4C absolute ethanol for 60 min followed by pure LRWhite at 4C for three periods of 60 min and, finally, embedded in pure LRWhite in gelatin capsules for polymerization at 50C for 48 hr.
Ultrathin sections (~80-nm thick) were cut on a RMC MTX ultramicrotome (Elexience; Verrières-le-Buisson, France), mounted on 200-mesh nickel grids coated with polylysine (dilution 1:1,000), and stabilized for 1 day at RT. Immunogold labeling was performed by flotation of grids on drops of reactive media. Nonspecific sites were coated with 1% BSA and 1% normal goat serum in 50 mM Tris-HCl, pH 7.4 for 60 min at RT. Thereafter, incubation was carried out overnight at 4C in a wet chamber with our purified rabbit polyclonal antibody raised against arginase II (dilution 1:100 or 1:150) in 1% BSA, 50 mM Tris-HCl, pH 7.4. Tissue sections were successively washed three times in 50 mM Tris-HCl, pH 7.4 and pH 8.2 at RT, incubated in a wet chamber for 45 min at RT in 1% BSA, 50 mM Tris-HCl, pH 8.2 for 20 min at RT, labeled with a goat anti-rabbit IgG gold-conjugated (20 nm; tebu-bio SA; Le Perray en Yvelines, France) diluted 1:25 in 1% BSA 50 mM Tris-HCl pH 8.2, successively washed three times in 50 mM Tris-HCl pH 8.2 and pH 7.4, and three times in filtrated distilled water. Sections were contrasted with aqueous saturated 3% uranyl acetate for 45 min in darkness and lead citrate for 5 min and observed on a transmission electron microscope (Jeol 1200EX; Tokyo, Japan) equiped with a MegaViewII digital camera and AnalySIS software (Eloise; Roissy Charles de Gaulle, France).
Chemicals
Salts and most chemicals, phenylmethylsulfonylfluoride, benzamidine, Ponceau S solution, secondary anti-IgG antibodies, and Kodak X-MAT films were purchased from Sigma (St. Quentin Fallavier, France). Protease inhibitor cocktail (DAPI) was purchased from Boehringer Mannhein (Strasbourg, France). ECL Western Blotting Kits and ImagerMaster Total Lab v2.01 program were purchased from Amersham (Buckinghamshire, UK and Orsay, France).
Results
Renal Distribution of Arginase II in Female and Male Rats
CSD and AR were used as specific markers of renal zones. In rat kidneys, the expression of CSD gene is restricted to CPST and OSPST (Reymond et al. 2000) whereas AR gene is expressed in the medullary and papillary collecting ducts (Sands et al. 1989). Our purified and specific antibody raised against the synthetic peptide corresponding to amino acids 318-332 of arginase II revealed a single band of 38 kDa that corresponds to the expected size of the mature arginase II (Swiss-Prot: rat O08701). Although the proximal tubules and collecting ducts originated from distinct embryological structures, the 38-kDa arginase II was detected in the cortical and medullary zones of the male and female rats (Figures 1A and 1B, left). Quantitation of the bands indicates that the highest levels of arginase II were found in osOM and IM (Figures 1A and 1B, right). These experiments also indicate that the peptide of arginase II exhibits the same molecular weight in the six renal zones.
Tubular and Subcellular Localization of Arginase II in Rat Inner Medulla
The quality of our purified arginase II antibody allowed histological investigations. Experiments were designed to demonstrate that arginase II was expressed in IMCD. As shown in Figure 2, arginase II immunostaining was very intense in tubules identified as IMCD and characterized by a large lumen and a relatively thick epithelium compared with that of the descending and ascending thin limbs. IMCD cells are also characterized by nuclei that appear in blue with DAPI near the apical membrane. All IMCD cells were strongly labeled, suggesting that the principal cells and probably the intercalated cells express the arginase II gene. Tubules with thin limbs were less immunostained than IMCD. These results are in good agreement with the high level of urea produced by microdissected IMCD and the low production of urea reported for microdissected descending and ascending thin limbs (Levillain et al. 1989).
Because the NH2-terminal amino acid sequence of the arginase II precursor predicts for mitochondrial import and because the abundance of mitochondria is very low in IMCD compared with OSPST, we decided to identify the subcellular localization of arginase II protein in IMCD by immunoelectron microscopy. The results show that in IM slides incubated without the antibody raised against arginase II but with the goat anti-rabbit IgG gold-conjugated antibody, the cytosol and mitochondria of IMCD were devoid of gold particles (Figure 3A). In contrast, when IM slices were incubated with our purified antibody raised against arginase II, 89% of the immunogold particles were visualized in the mitochondria and 11% in the cytosol (Figures 3B-3D). No labeling was observed in nuclei (data not shown).
To confirm the mitochondrial localization of arginase II in IMCD and to quantify the level of arginase II in each subcellular compartment, the IM of four female rats were dissected to obtain proteins of the cytosolic and mitochondrial fractions and to analyze them by Western blot. In addition, because arginase II has been localized in the mitochondria of the rat OSPST (Levillain et al. 2004), the osOM of the same female rats was dissected to obtain proteins of the cytosolic and mitochondrial fractions to constitute a positive control. As shown in Figure 4A, the mitochondrial fraction was characterized by the presence of the following proteins: α-subunit F1-ATP synthase (55 kDa), core 2 (45 kDa), and ISP (25 kDa). F1-ATP synthase and core 2 are bound to the inner mitochondrial membrane and located on the matrix side of the membrane, whereas ISP is also in the inner mitochondrial membrane but located on the opposite side of the inner membrane (intermembrane space). Core 2 and ISP belong to the protein complex III of the respiratory chain. On this basis, our results clearly show that our arginase II antibody revealed a single band of 38 kDa in the cytosolic and mitochondrial fractions prepared from osOM and IM. In the rat osOM and IM, 95% and 88%, respectively, of the mature arginase II were detected in isolated mitochondria. Interestingly, the significant amount of the mature arginase II detected in cytosol of osOM and IM did not correspond to the predicted size of 40 kDa for the precursor of arginase II (Gotoh et al. 1996). The abundance of the proteins α-subunit F1-ATP synthase, core 2, and arginase II were quantitated by densitometry to calculate the relative abundance of each protein in the mitochondrial fraction of osOM and IM. As shown in Figure 4B, the levels of the α-subunit F1-ATP synthase and core 2 were similar in osOM and IM because their ratios were equal to 1. On this basis, our results indicate that the mitochondrial fraction of IM contained 1.5-fold more arginase II than that of the osOM.
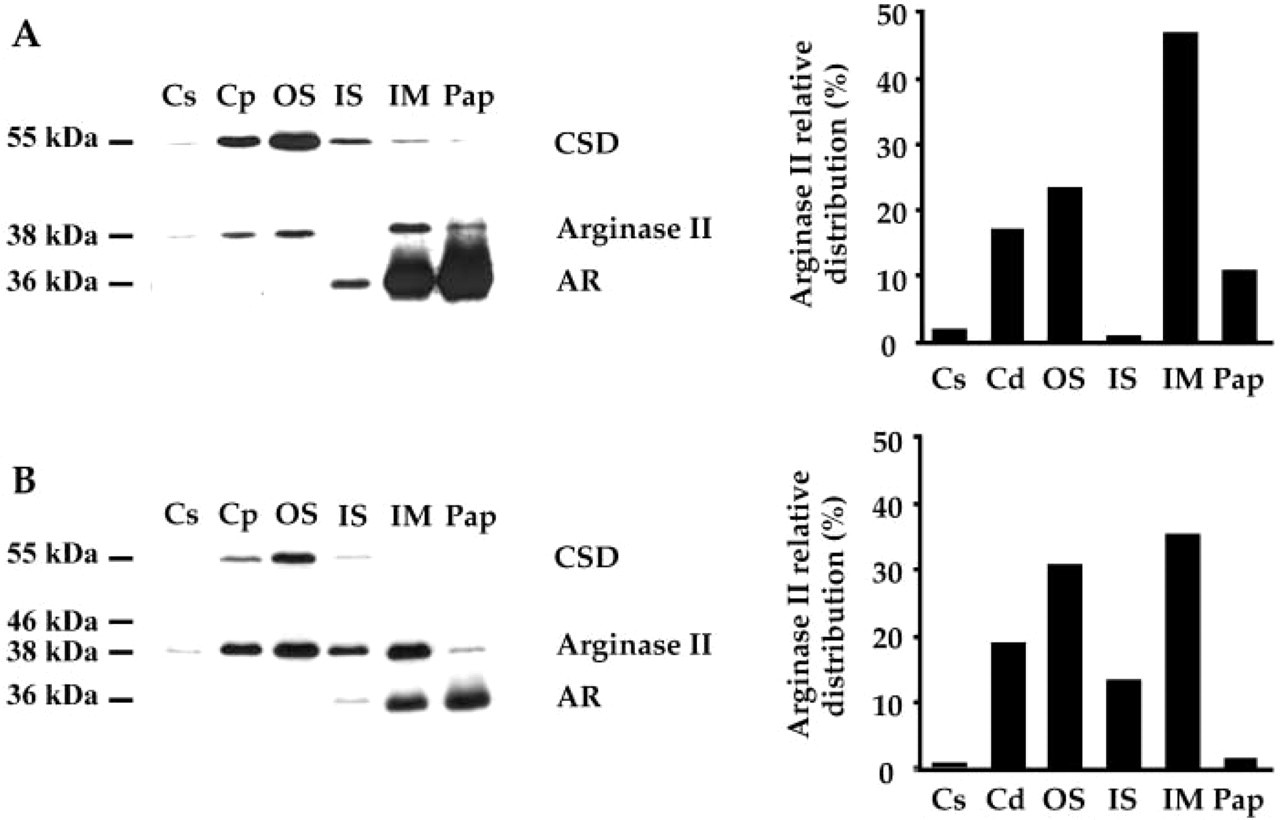
Western blot analysis of cysteine sulfinate decarboxylase (CSD), arginase II, and aldose reductase (AR) levels in the main renal zones of male (
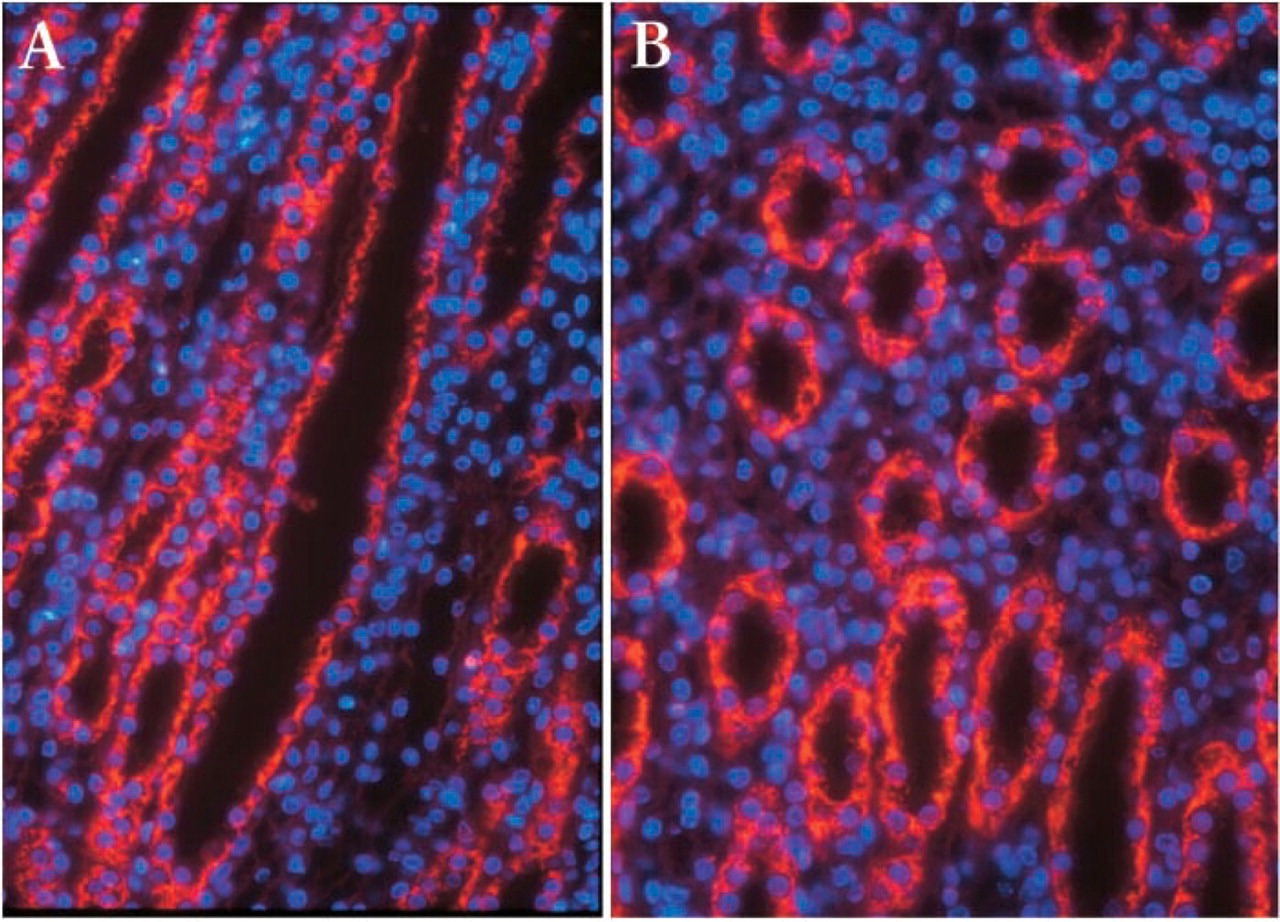
Immunofluorescent detection of arginase II in the male rat kidneys. (
Together our results prove that arginase II protein is highly expressed in IMCD and is almost exclusively located within the mitochondria.
Discussion
Localization of Arginase II in IMCD Mitochondria
These experiments were conducted to reevaluate whether arginase II gene is expressed in the rat IMCD. Indeed, Levillain et al. (1989) reported that a high production of urea occurs precisely in IMCD of male Sprague Dawley rats, whereas Miyanaka et al. (1998) detected the immunoreactive arginase II in PSTs but not in the entire collecting duct including the IMCD of Wistar rats. The discrepancy between these results may be related either to the strain of rats used, a structural difference between the arginase II expressed in PSTs and IMCDs, or explained by the use of an antibody unable to recognize the arginase II expressed in the collecting ducts. This is why we developed a new antibody against arginase II to detect both the precursor and the mature arginase II protein by different techniques.
The rat arginase II gene encodes for a protein with a predicted molecular weight of 38,645 Da (Iyer et al. 1998). As expected, our antibody raised against arginase II revealed a band of ~38 kDa by Western blotting. This single peptide of arginase II was detected in all renal zones of both female and male rats, but its relative abundance was higher in IM than in the other renal zones. This typical distribution pattern of arginase II characterizes the rat kidney and deeply differs from that of the other mammals that express a very low arginase II activity in their collecting duct (Levillain and Hus-Citharel 1996). It remains still unexplained for which physiological interests rats expressed arginase II in the collecting duct. The present immunohistological study reveals that within IM, arginase II is highly expressed in IMCD compared to thin descending and ascending limbs of Henle's loop. The level of arginase II is in good agreement with the production of urea in IMCD (Levillain et al. 1989). Recently, the 38-kDa arginase II was also identified by Matrix Assisted Laser Desorption Ionization Time-of-Flight (MALDI-TOF) Mass Spectrometry in isolated IMCD (van Balkom et al. 2004). Interestingly, IMCD are composed of 90% principal and 10% intercalated cells that differ in morphology and cell functions (Tisher and Madsen 2000). Intercalated cells are present only in the upper third, but not in the two lower thirds, of the rat inner medulla. Because immunofluorescence of arginase II was homogeneously detected along IMCD, our results clearly indicate that the principal cells express the arginase II gene and suggest that intercalated cells may also express this gene. Double staining of the principal cell by using a specific antibody such as aquaporin-3 would have been hepful to clarify this point. However, because the population of intercalated cells is very low in IMCD and because the function of arginase II in this cell type will be very difficult to identify, double immunostaining was not performed.
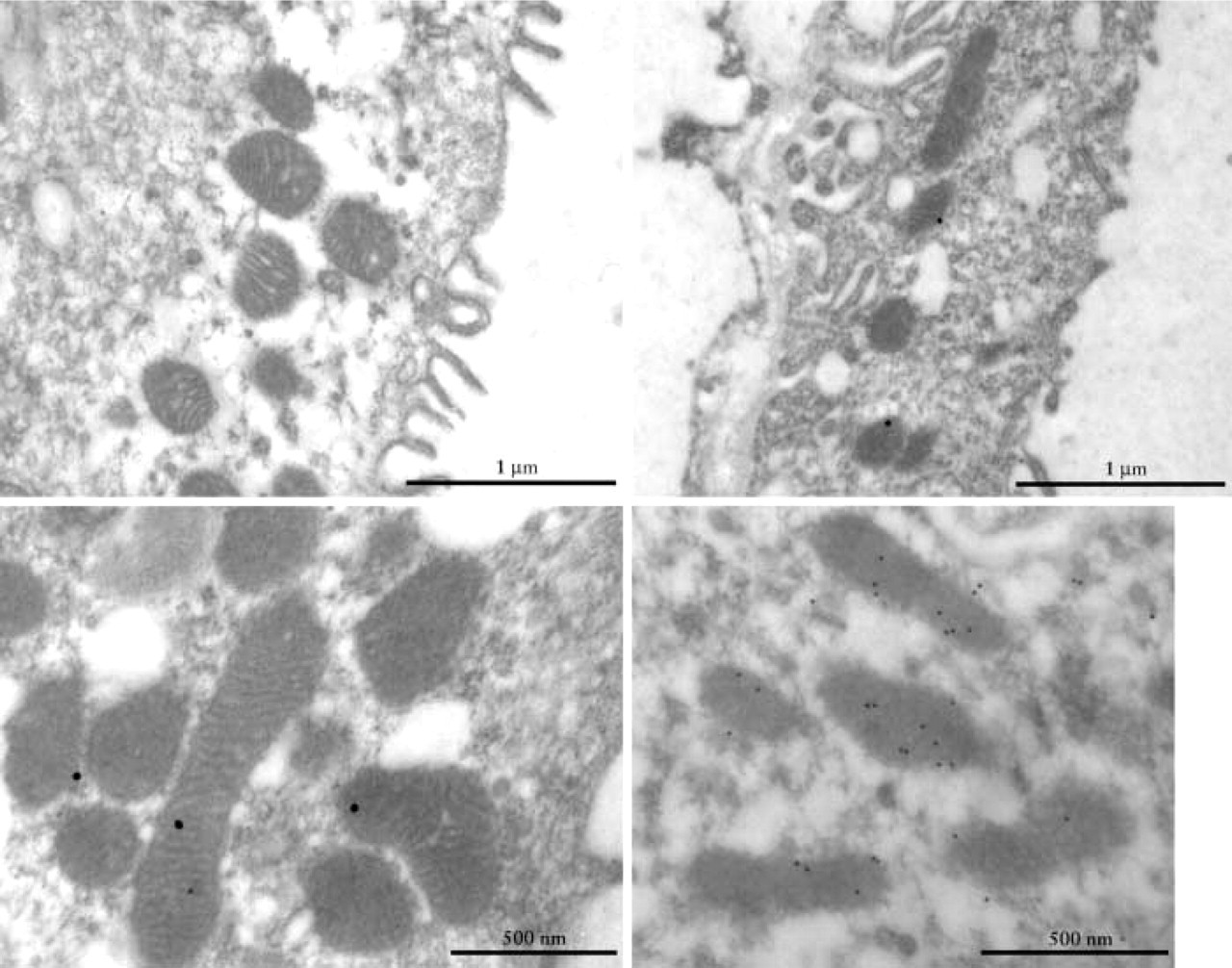
Transmission electron micrograph of mitochondria from female rat IMCD cells subjected to immunogold labeling of arginase II. (Upper left) Gold particles were absent in mitochondria of IMCD cells when slides were incubated without our purified rabbit polyclonal antibody raised against arginase II. (Upper right, lower left, lower right) Immunogold labeling was mainly localized in the mitochondria of IMCD cells when incubated with our purified rabbit polyclonal antibody raised against arginase II. Few gold particles were detected in the cytosol of IMCD cells. (Upper right) Partial view and (lower left) mitochondria of IMCD cell incubated with our arginase II antibody (dilution 1:150) and 20-nm gold particles. (Lower right) Mitochondria of IMCD cell incubated with our arginase II antibody (dilution 1:100) and 20-nm gold particles.
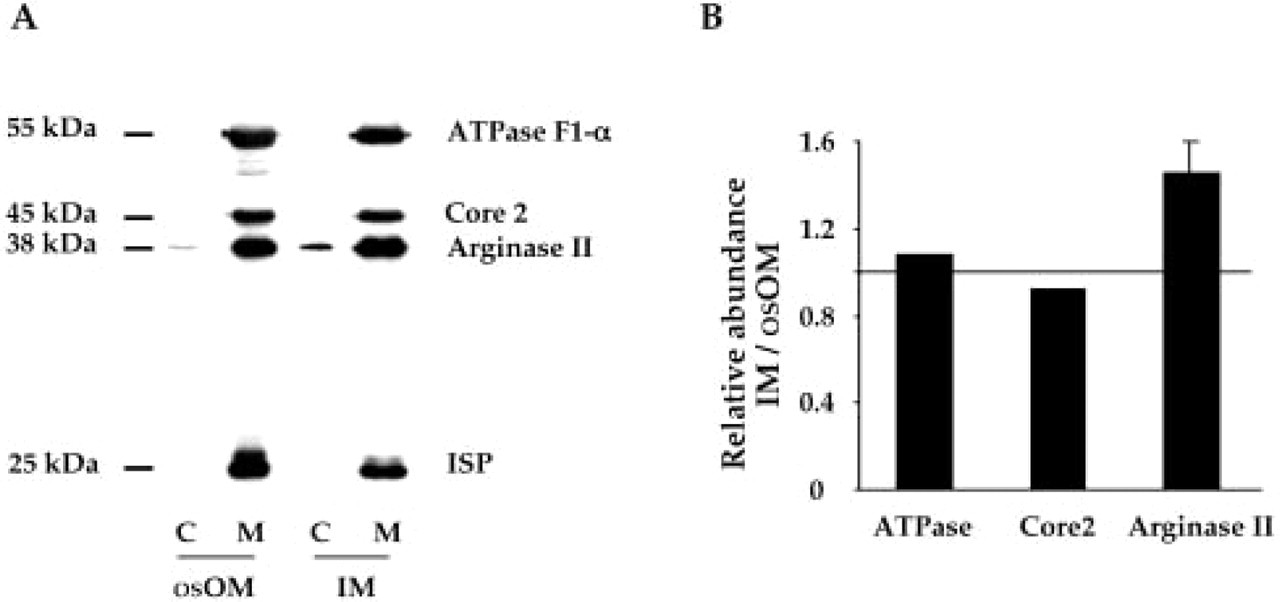
Subcellular localization and relative abundance of arginase II in female rat osOM and IM. Immunoblot performed to analyze the proteins of the cytosolic and mitochondrial fractions obtained from dissected osOM and IM of female rats. Forty-μg protein samples of each subcellular compartment were subjected to 10% SDS-PAGE. (
Molecular biology databases reveal that arginase II is a nucleus-encoded enzyme of the mitochondrial matrix composed of 354 amino acid residues and possesses an N-terminal mitochondrial-targeting sequence (Shi et al. 1998). The in vitro synthesis of arginase II precursor allowed the detection of a polypeptide of ~40 kDa that can be imported in isolated rat liver mitochondria to process to the mature form of ~38 kDa (Gotoh et al. 1996). The molecular size of 38 kDa for the mature arginase II has been confirmed in mitochondria isolated from kidneys of male and female rats and mice (Levillain et al. 2005; Levillain et al. 2004). Regarding the 40-kDa arginase II precursor, it was speculated that the first 22 amino acid residues may correspond to the signal for mitochondrial import. This small peptide sequence is highly conserved in rat and mouse arginase II but differs from that of the human arginase II (see Table 1). However, in the present study, the 40-kDa arginase II was undetectable on Western blot performed with proteins of the cytosolic fractions extracted from either osOM or IM of female rat kidneys. Regarding the subcellular localization of arginase II, our antibody raised against arginase II allows the detection of the 38-kDa mature arginase II almost exclusively in mitochondria of the female rat IMCD and OSPST. Inasmuch as the bulk of the mature arginase II was detected in mitochondria, we hypothesize that the maturation and the import of the 40-kDa arginase II within the mitochondria may occur rapidly after translation. The presence of a low amount of the 38-kDa mature arginase II in the cytosol is quite difficult to explain, unless some mitochondria were injured and broken during the different steps of isolation. In this case, arginase II would have been easily released from the matrix. In contrast, if only a few mitochondria were broken, neither α-subunit F1-ATP synthase, core 2, nor ISP would have been detected in the cytosolic fraction, because these proteins belong to the mitochondrial membrane. Whatever the explanation, our results are in good agreement with those of other authors who reported an arginase II activity in mitochondria isolated from the rat kidney (see review, Jenkinson et al. 1996).
Putative NH2-terminal mitochondrial-targeting presequence of arginase II
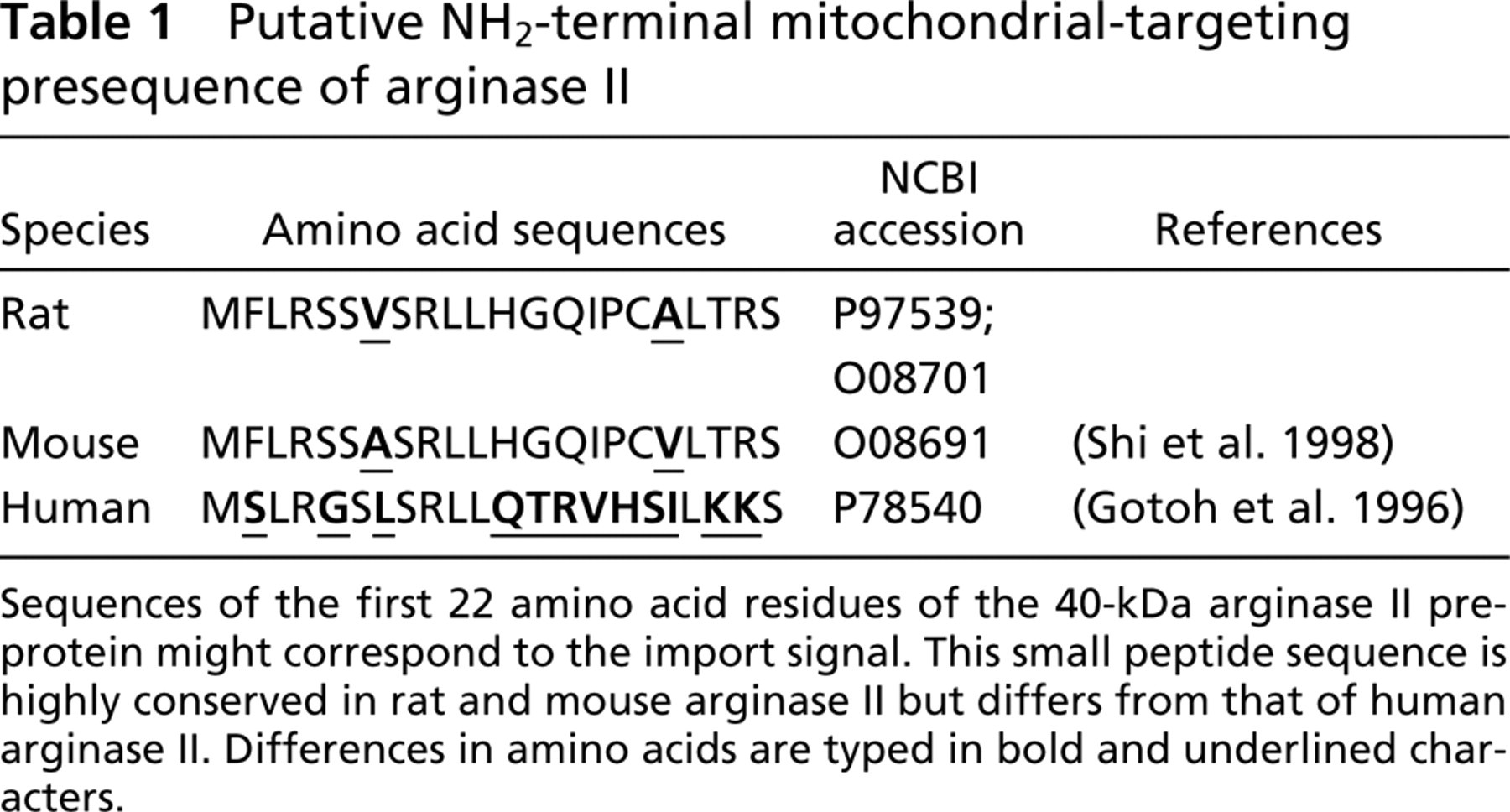
Sequences of the first 22 amino acid residues of the 40-kDa arginase II preprotein might correspond to the import signal. This small peptide sequence is highly conserved in rat and mouse arginase II but differs from that of human arginase II. Differences in amino acids are typed in bold and underlined characters.
The subcellular localization of arginase II in mitochondria of PSTs and IMCD requires an additional comment because CPST and OSPST cells contain more mitochondria than those of IMCD (Pfaller 1982). Indeed, the mitochondrial volume per unit tubular length of nephron segment is 42- and 28-times higher in CPST and OSPST, respectively, than in IMCD (S1 + S2: 359 ± 14 to 325 ± 14; S3 or OSPST: 240 ± 7, and IMCD: 8.1 to 9.1 μm3/μm). However, the production of urea in male rat-dissected OSPST is only twofold higher than that of the IMCD (250 ± 16 vs 127 ± 16 fmol/min/mm tubular length, respectively) (Levillain et al. 1989). When estimating the relative production of urea per mitochondrial volume in both OSPST and IMCD, the result suggests that the mitochondrial concentration of arginase II is more abundant in mitochondria of IMCD compared with those of OSPST. Indeed, our results confirm that the amount of arginase II contained in mitochondria of IM was 1.5-fold higher than in mitochondria of osOM.
Interestingly, the molecular weight of arginase II deduced from our Western blot is similar in the different renal zones and in mitochondria isolated from rat osOM and IM. This result sustains the view of a single gene of arginase II expressed in the kidney but does not exclude posttranslational modifications leading to arginase II isoforms. At present, the mitochondrial localization of arginase II in the rat IMCD has not been associated with physiological roles and a metabolic pathway.
Physiological Roles of Arginase II in IMCD
The biochemical reaction carried out by arginase II simultaneously decreases the concentration of
In conclusion, arginase II gene is expressed in male and female rat IMCD, and the mature arginase II is localized into the mitochondria. Although the physiological roles of arginase II remain to be identified, it is noteworthy that both arginase II and NOS are highly expressed in IMCD.
Footnotes
Acknowledgments
The authors are indebted to Dr. Anne-Marie Lefrançois-Martinez (Université Blaise Pascal, UMR 6547 CNRS, Aubière, France), Dr. Marcel Tappaz (Université Claude Bernard, Fac. Médecine RTH Laennec, U433 INSERM, Lyon, France), and Dr. Catherine Godinot (Université Claude Bernard, CGMC, UMR 5534 CNRS, la Doua, Villeurbanne, France) who kindly provided the rabbit against mouse AR antibody, rabbit against CSD antibody, rabbit against α-subunit F1-ATP synthase antibody, rabbit against core 2 antibody, and mouse against ISP antibody, respectively. Dr. Olivier Levillain is indebted to Prof. Brigitte Kaissling (Institute of Anatomy, University of Zurich, Zurich, Switzerland) for stimulating discussion, Mireille Mutin (Université Claude Bernard, Faculté de Médecine RTH Laennec, U433 INSERM, Lyon, France) for teaching immunohistochemistry, Jocelyne Vial for assistance, and Bernard Marchand for his contribution in the animal house.