
Abstract
The endothelin/endothelin-receptor system is a key player in the regulation of vascular tone in mammals. We raised and characterized an antiserum against rat ETB receptor and investigated the distribution of ETB receptors in different vascular beds during postnatal development (day 0 through day 28) and in the adult rat. We report the tissue-specific and age-dependent presence of vasoconstrictor ETB receptors. At the time of birth, vascular smooth muscle cells from all tissues examined did not exhibit ETB receptor immunoreactivity. The occurrence of ETB receptor immunoreactivity in the postnatal development was time dependent and started in small coronary and meningeal arteries at day 5, followed by small mesenteric arteries as well as brachial artery and vein at day 14. At day 21, ETB receptors were present in the media of muscular segments of pulmonary artery, large coronary arteries, and intracerebral arterioles. At day 28, ETB receptor immunoreactivity was evident in interlobular renal arteries, vas afferens, and efferens. Large renal arteries, mesenteric artery, and elastic segments of pulmonary arteries, as well as coronary and mesenteric veins, did not exhibit ETB receptor immunoreactivity. These data demonstrate the age-dependent and tissue-specific presence of ETB receptors, mainly on arterial smooth muscle cells in the vascular system of the rat.
E
ET-1 exerts its biological effects through two distinct ET-receptor subtypes, ETA and ETB. ETA receptors on smooth muscle cells exclusively mediate vasoconstriction, whereas ETB receptors can mediate both vasodilation as well as vasoconstriction, depending on their location on endothelial cells and vascular smooth muscle, respectively. While the role of the ETA receptor subtype in mediating vasoconstriction is beyond debate, the existence of ETB receptors on vascular smooth muscle cells in distinct vascular beds is still controversial, and systematic histological investigations on the distribution pattern of vascular ET receptors are missing. Recently, colocalization of ETA and ETB receptors on arterial smooth muscle cells in the coronary circulation (Wendel-Wellner et al. 2002) and small pulmonary arteries of adult rats were demonstrated (Soma et al. 1999).
Pharmacological studies aimed at defining the role of vasoconstrictor ETB receptors are highly controversial, and studies from different groups lead to contradictory results even for identical organ systems (Takase et al. 1995; Sharifi and Schiffrin 1996; Touyz et al. 1995; Mickley et al. 1997; Rizzoni et al. 1997; Adner et al. 1998). In addition, using the synthetic agonists and antagonists RES 701-1, BMS 184696, and BQ 788, the existence of two ETB receptor subtypes was postulated (Natarajan et al. 1995; Gellai et al. 1996; Schroder et al. 1998), namely, ETB1 receptors on the endothelium mediating vasodilation and vasoconstrictor ETB2 receptors located on vascular smooth muscle cells.
In the ovine fetal lung, mRNA levels of both ETA and ETB receptors were studied and increased during late gestation, thereby providing evidence for developmental regulation of the ET/ET-receptor system (Ivy et al. 2000). Studies in newborn piglets and lambs indicated that endogenous ET-1 exerts a tonic vasoconstrictor effect on swine intestinal exchange vasculature (Nankervis and Nowicki 2000) and fetal lamb pulmonary circulation (Ivy et al. 1996). During postnatal development, a loss of ETB receptor-mediated vasodilation and increased sensitivity toward the vasoconstrictory effects of exogenous ET-1 in piglets was observed (Perreault and De Marte 1993; Nankervis and Nowicki 2000). In contrast, in fetal lamb pulmonary circulation, ET-1-induced vasodilation was observed in pulmonary veins but not in arteries (Wang and Coceani 1992; Wang et al. 1994).
In the newborn rat, idiopathic pulmonary hypertension is associated with increased lung ET-1 (Stelzner et al. 1992), and in a model of genetic ETB receptor deficiency, hypoxic pulmonary hypertension and ET-1-induced vasoconstriction were exaggerated (Ivy et al. 2001). In addition to the pulmonary circulation, information on the developmental aspects of ET-receptor expression or function in the rat is very limited. In rat kidney, Abadie and coworkers reported a decrease of ETA receptor-binding capacity during the first month of life, while binding to the ETB receptor was not affected (Abadie et al. 1996). However, no data on the cellular distribution of the ET-receptor subtypes were provided. In the adult rat kidney, the ETB receptor seems to play a significant role in mediating ET-1-induced vasoconstriction of resistance vessels (Wellings et al. 1994; Endlich et al. 1996), but ETB receptor protein or mRNA could not be identified on vascular smooth muscle cells by either immunohistochemistry (Yamamoto and Uemura 1998) or in situ hybridization (Hocher et al. 1995).
The goal of our study was to determine the localization of ETB receptors in the vascular system of the adult rat and during postnatal development. For this purpose, we raised an antiserum against rat ETB receptor and investigated tissue samples from heart, lung, kidney, small intestine, brain, the brachial vessels, and skeletal muscle at different time points after birth. Our results show differential distribution and postnatal development of ETB receptors on smooth muscle of the rat vasculature.
Materials and Methods
Animals
Tissue samples from heart, lung, kidney, small intestine, brain, brachial vessels, and biceps brachii muscle were obtained from male Wistar rats at postnatal day (PD) 0, PD 5, PD 14, PD 21, and PD 28 (n=2, each) after decapitation and from adult Wistar rats after chloroform inhalation (n=5). Immediately after removal, tissues were frozen in isopentane/liquid nitrogen and stored at −80C.
Generation of the ETB Receptor Antibody
A peptide sequence corresponding to amino acid residues 39 to 46 from the non-homologous aminoterminal region of the rat ETB receptor was chosen for generation of a polyclonal anti-ETB receptor antiserum. Immunization of rabbits with the peptide coupled to keyhole limpet hemocyanine, and affinity chromatographic purification were performed by Eurogentec (Seraing, Belgium).
Characterization of the ETB Receptor Antibody by Western Blot
The polyclonal rabbit anti-ETB receptor antibody was first characterized by Western blot. Rat kidneys and lungs were homogenized at 4C in homogenization buffer (100 mM HEPES, 250 mM sucrose, 5 mM EDTA, 0.5% Triton X-100, 200 μM Pefabloc, 2 μM pepstatin, and 2 μM leupeptin) using an Ultra Turrax tissue homogenizer (IKA; Staufen, Germany). After centrifugation at 2500 rpm at 4C (5417 R; Eppendorf, Hamburg, Germany), the supernatant was collected and centrifuged again at 14,000 rpm. The supernatant was then mixed with Rotiload buffer (Roth; Karlsruhe, Germany) and heated to 70C for 5 min. The probe was then electrophoresed on a 10% acrylamide gel under non-reducing conditions. The gel was then blotted on a polyvinylidene difluoride membrane (PVDF; Millipore, Eschborn, Germany) by semidry blotting at 100 mA for 75 min using Novex Transfer buffer (Invitrogen; Karlsruhe, Germany).
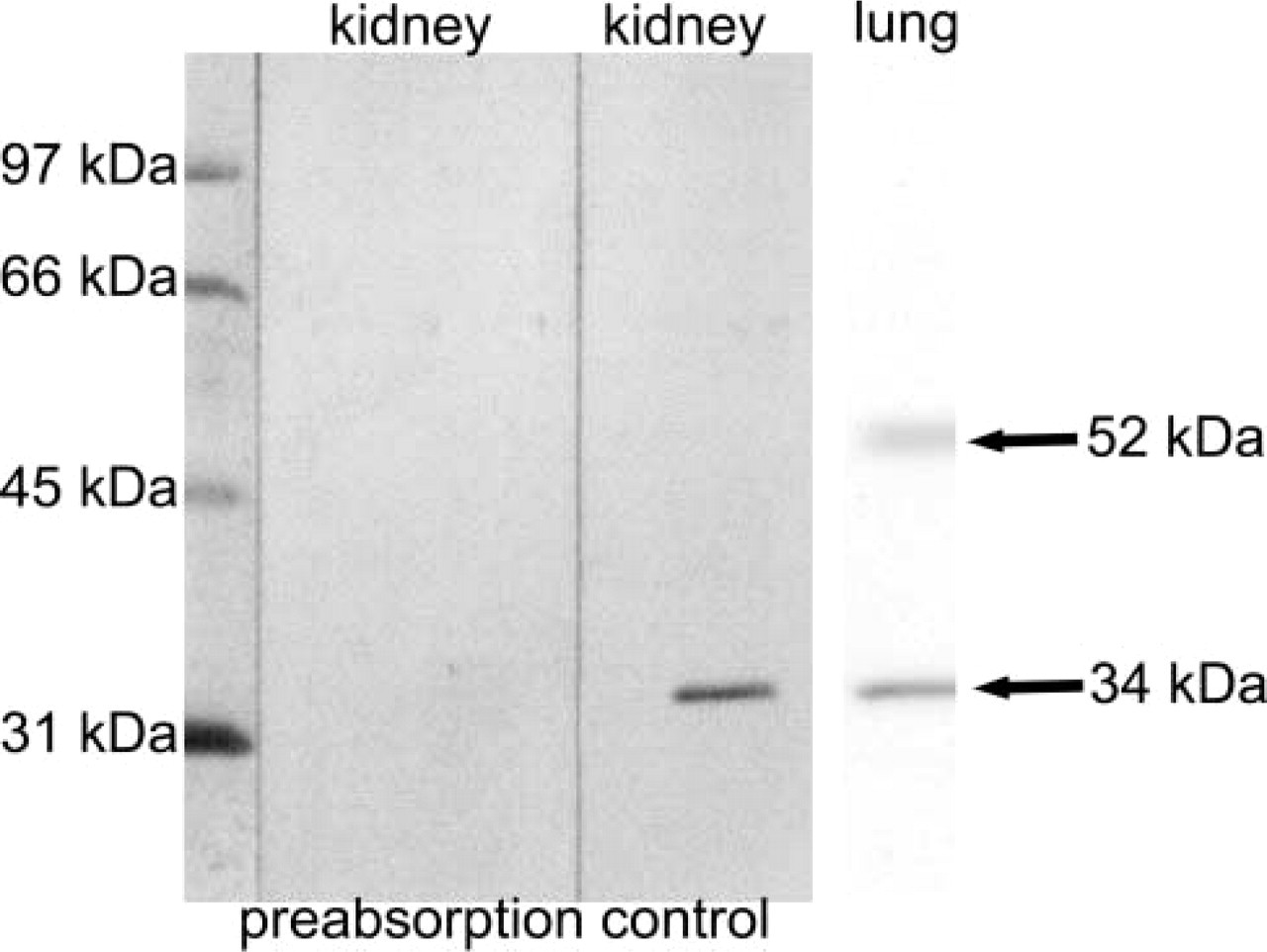
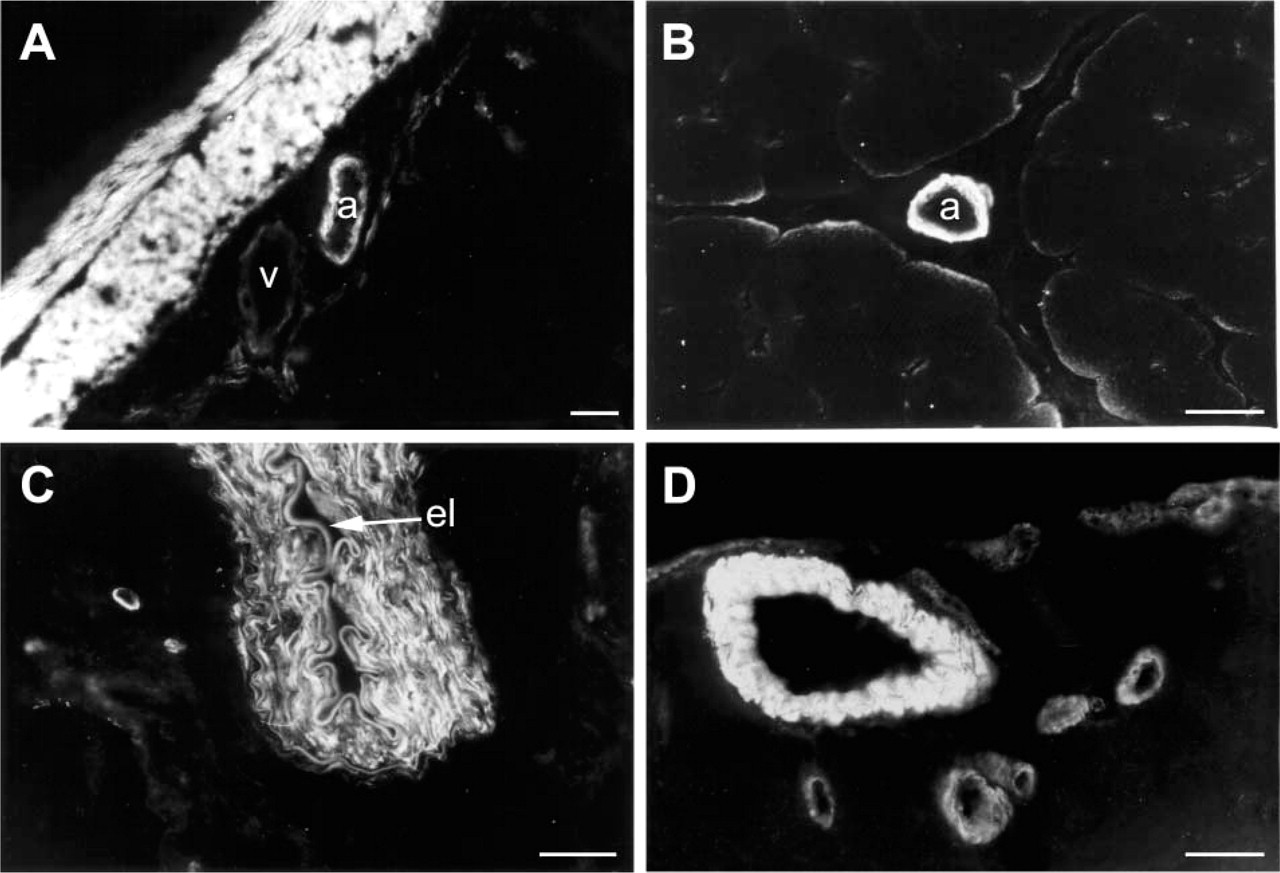
Western blot and preabsorption for the ETB receptor antibody. Rat kidney and lung homogenates were loaded on SDS-acrylamide gels (20 μg per lane). On rat kidney blots, the antiserum recognizes a protein with a molecular weight of ∼34 kD (control). The band is abolished by preabsorption with the peptide used for immunization (preabsorption). Incubating rat lung blots with the ETB receptor antiserum resulted in two bands, of ∼34 kD and 52 kD.
Blots were blocked with 5% dry milk powder in Tris-buffered saline (TBS), pH 7.4, for 1 hr. The primary antibody was then incubated at 4C overnight. Blots were washed twice with TBS and incubated with alkaline phosphatase-conjugated anti-rabbit immunoglobulin, 1/2500 (Promega; Mannheim, Germany) for 1 hr at room temperature. After washing twice, blots were incubated with NBT/BCIP substrate (Boehringer; Mannheim, Germany) for a maximum of 10 min.
Working dilution for the antibody in Western blot was determined to be 1/80.
Specificity was evaluated by preabsorption. Primary antibody was diluted 1/80 and preabsorbed with the immunization peptide (20 ng/ml) at 4C overnight. Then, one blot each was incubated with either preabsorbed or non-preabsorbed primary antibody as described above.
Single-labeling Immunofluorescence
Six-μm-thick cryosections from rat tissues were fixed with acetone at −20C for 10 min and air dried for 2 hr. They were then blocked with 10% normal porcine serum in phosphate-buffered saline (PBS), pH 7.4, for 30 min. Thereafter, sections were incubated with serial dilutions of the polyclonal rabbit anti-ETB receptor antibody at room temperature overnight. After washing twice in PBS, sections were incubated with fluoresceinisothiocyanate (FITC)-labeled goat anti-rabbit immunoglobulin, 1/400, (Diagnostic International; Schriesheim, Germany) for 1 hr, then washed again and coverslipped in carbonate-buffered glycerol, pH 8.6. Sections were evaluated with the BX 60 epifluorescence microscope (Olympus; Hamburg, Germany). Working dilution for the antibody was determined to be 1/80 for immunofluorescence. Specificity was evaluated by preabsorption as described above.
Double-labeling Immunofluorescence
Acetone-fixed cryosections (6 μm) from rat tissues were blocked and then incubated with rabbit anti-ETB receptor antibody, 1/80, and monoclonal mouse anti-rat endothelial antigen (RECA)-1 antibody (Serotec; Hamburg, Germany), 1/40. The anti-RECA-1 antibody was demonstrated to recognize a defined epitope that is present on all rat endothelial cells (Duijvestijn et al. 1992). Secondary antibodies used for double-labeling immunofluorescence were FITC-labeled goat anti-rabbit IgG, 1/400, and Cy3-labeled donkey anti-mouse IgG, 1/1000 (Dianova; Hamburg, Germany).
Results
Characterization of the ETB Receptor Antiserum
Specificity of the antiserum was evaluated by Western blotting and immunofluorescence with preabsorption.
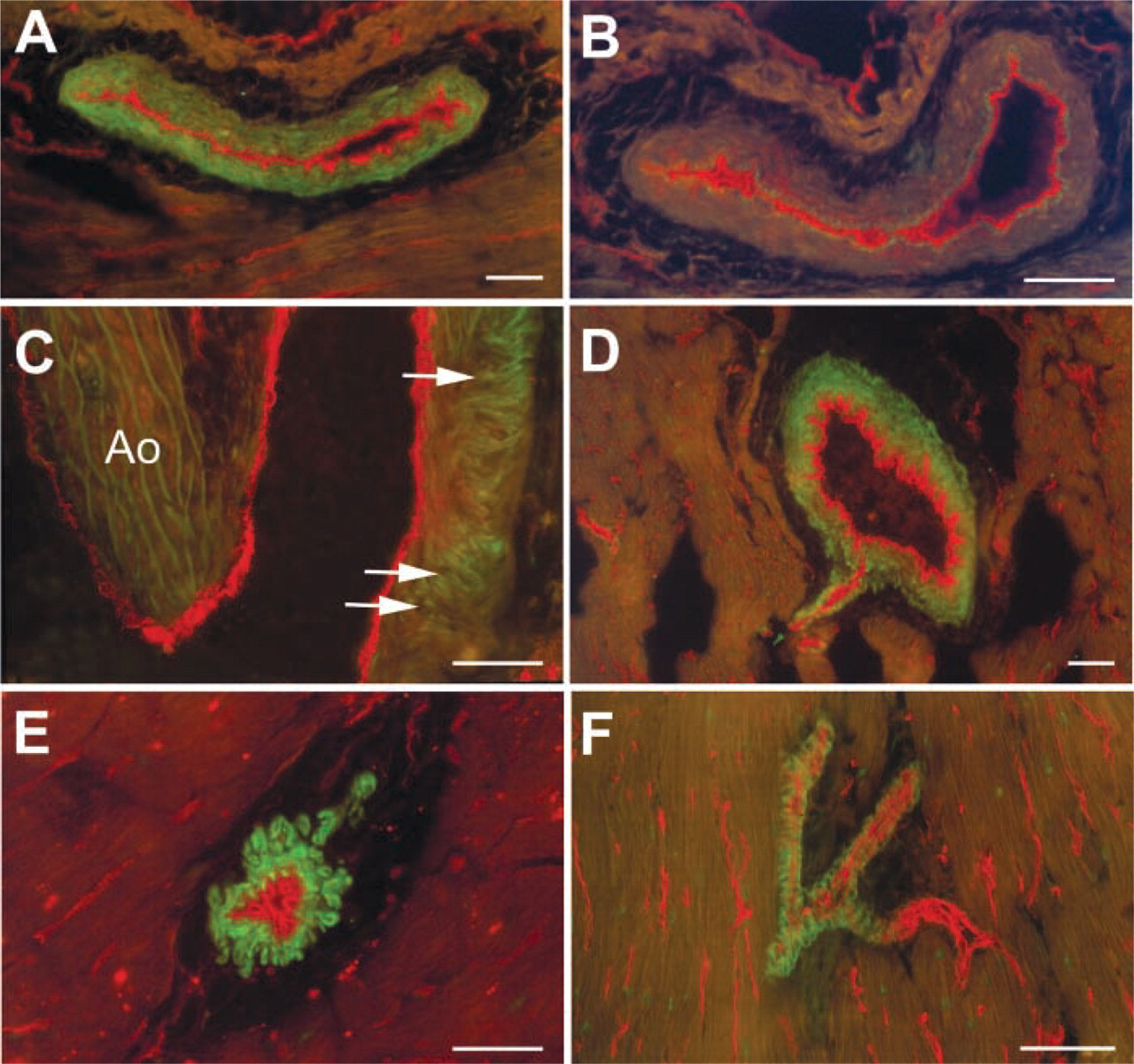
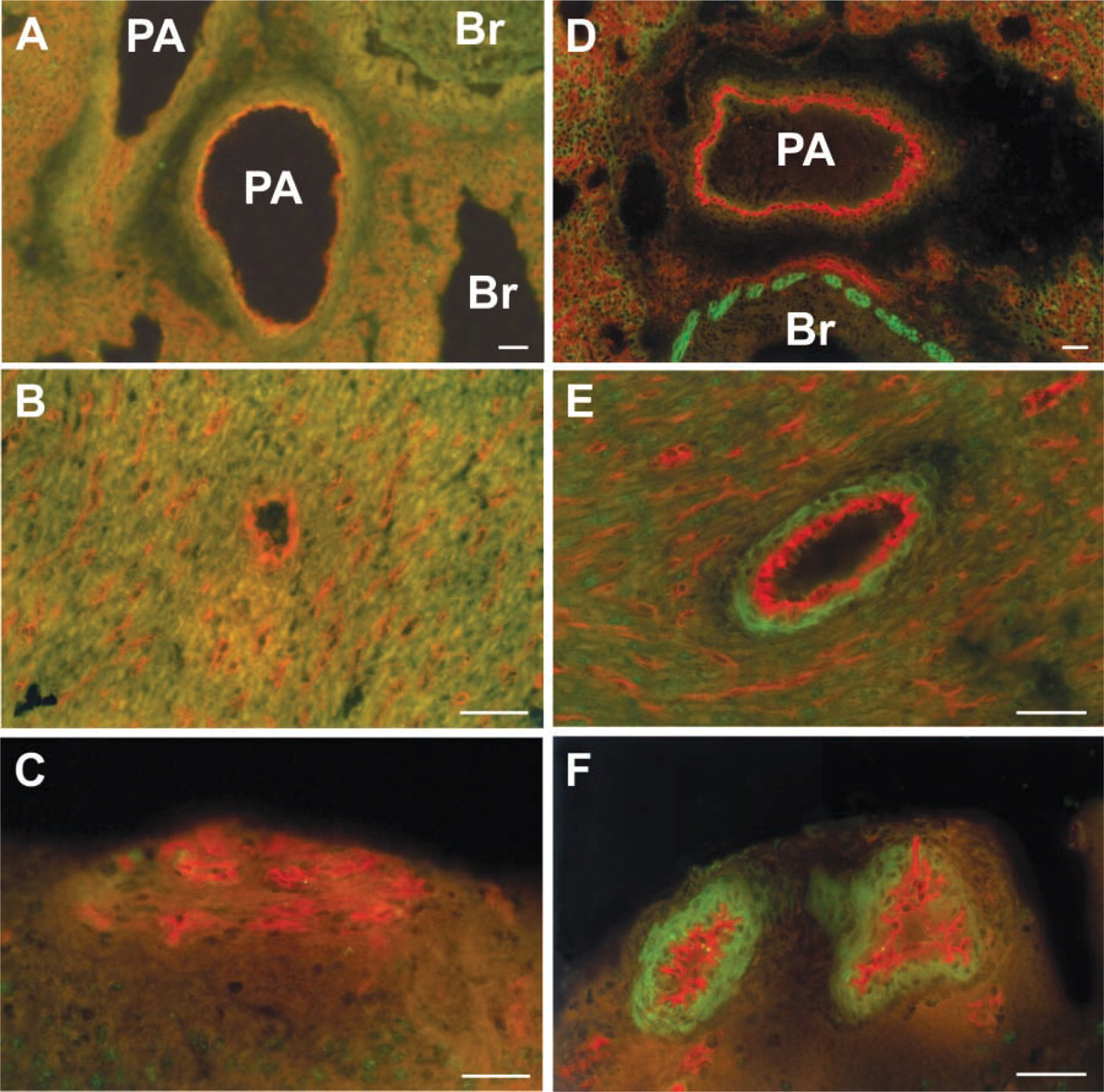
Double-labeling immunofluorescence of the coronary circulation for the ETB receptor antibody (green fluorescence) and the panendothelial marker RECA-1 (red fluorescence). Labeling of coronary artery smooth muscle cells (
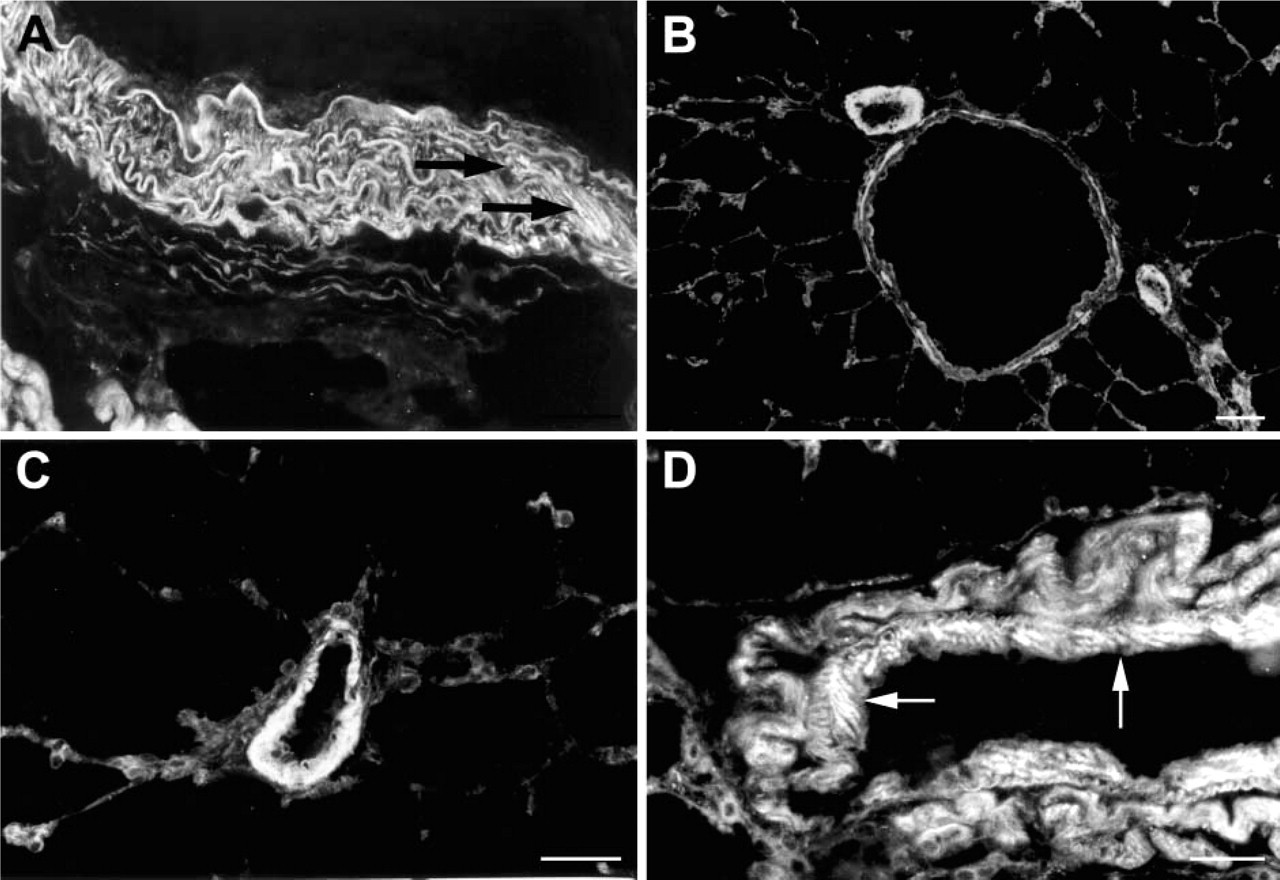
Single-labeling immunofluorescence of the ETB receptor in the pulmonary circulation. In elastic segments of the pulmonary artery, only scarce labeling of vascular smooth muscle cells by the ETB receptor antibody can be observed (arrows in
Western Blotting. Incubation of rat kidney Western blots with the ETB receptor antiserum resulted in a single band of ∼34 kD, which was not detected after preabsorption. Incubation of rat lung Western blots with the ETB receptor antiserum resulted in a major band of 34 kD and a weaker band of ∼52 kD (Figure 1).
Immunofluorescence. ETB receptor immunoreactivity was observed on vascular smooth muscle cells and abolished by preabsorption with the peptide used for immunization. Endothelial cells were not labeled by the antiserum (Figures 2A and 2B).
Distribution of ETB Receptors in the Vascular System of the Adult Rat
Cardiopulmonary Circulation. In the coronary circulation, ETB receptors were present on smooth muscle of coronary arteries and arterioles (Figures 2C-2F). In the lung circulation, ETB receptor immunoreactivity was evident on smooth muscle cells of bronchial arteries, muscular segments of pulmonary arteries, and pulmonary veins (Figure 3).
Renal Circulation. In the renal circulation, ETB receptors were present on smooth muscle cells from arcuate and interlobular arteries as well as vas afferens and efferens. No ETB receptors could be detected in the renal artery and its interlobar branches (Figure 4).
Mesenteric and Skeletal Muscle Circulation. Mesenteric and skeletal muscle vascular beds contribute significantly to total vascular resistance and determine systolic blood pressure. ETB receptors were located on smooth muscle cells of arterioles but not veins in both the mesenteric and skeletal muscle vascular tree (Figures 5A and 5B). However, although larger segments of mesenteric artery were devoid of ETB receptors, ETB receptor immunoreactivity was evident in the brachial artery and vein (Figure 5C).
Cerebral Circulation. In the cerebral circulation, smooth muscle cells of pial as well as intracerebral arterioles exhibited ETB receptor immunoreactivity (Figure 5D), whereas the media of the carotid artery did not (not shown).
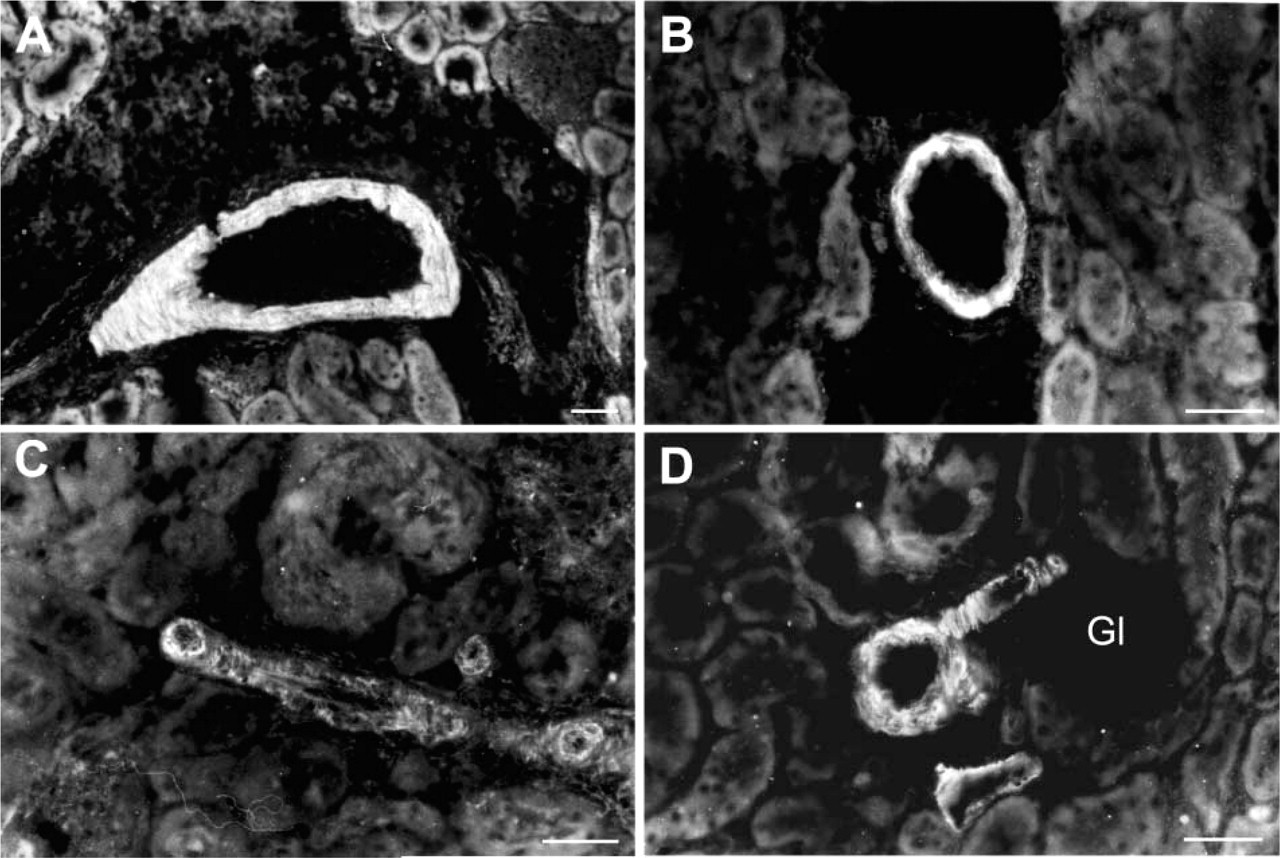
Single-labeling immunofluorescence of the ETB receptor in the renal circulation. Pre-glomerular arcuate and interlobular arteries display ETB receptor immunoreactivity (
ETB Receptors in Postnatal Development
PD 0. No vascular ETB receptor immunoreactivity could be detected at PD 0 in any tissue examined (Figures 6A-6C).
PD 5. At PD 5, pulmonary vascular smooth muscle cells were still devoid of ETB receptors, whereas bronchial smooth muscle exhibited ETB receptor immunoreactivity (Figure 6D). In the heart and brain, the media of small coronary and meningeal arteries displayed ETB receptor immunoreactivity (Figures 6E and 6F).
PD 14. Beginning on PD 14, smooth muscle cells of mesenteric arterioles, brachial artery and vein, and arterioles of the skeletal muscle vascular bed were labeled by the ETB receptor antiserum (Figures 7A-7C).
PD 21 and PD 28. Starting on PD 21, ETB receptors were also present on smooth muscle cells from intracerebral arterioles (Figure 7D), bronchial arteries (Figure 7E), and large coronary artery segments (not shown). Renal and mesenteric arteries, as well as the pulmonary artery, did not exhibit ETB receptor immunoreactivity. On PD 28, strong labeling of the media of muscular pulmonary arteries by the ETB receptor antiserum was observed (Figure 7F).
Discussion
This is the first study that systematically characterizes the distribution of ETB receptors in the vascular system of the rat. The antiserum used in this study was obtained by immunization with a peptide consisting of amino acid residues 39 to 46 from the amino terminus of rat ETB receptor. In Western blots, this affinity-purified antiserum recognized a single band of ∼34 kD. Kozuka and coworkers isolated and characterized the ETB receptor from bovine lung and reported two products, of 52 kD and 34 kD (Kozuka et al. 1991). Similar findings were reported by Shvartz et al. (1990), who identified two proteins, of ∼52 kD and 30 kD. The lower-molecular-weight form of ETB receptor is thought to result from proteolytic degradation of the ETB receptor molecule. In line with the report by Abe et al. (2000), who described the ETB receptor as mainly intracellular and degraded, we observed cytoplasmatic labeling of vascular smooth muscle cells. Endothelial cells were not labeled by the antiserum. The reason for this is unknown. Differential cell-type-specific posttranslational modification of the ETB receptor molecule may account for the lack of endothelial binding, because the epitope used for immunization lies in close proximity to a potential glycosylation site at amino acid residue 60 (Roos et al. 1998). However, there are no studies addressing this issue in vascular smooth muscle and endothelial cells. In addition, there is no evidence that ETB receptors on vascular smooth muscle and endothelial cells represent different ETB receptor gene products generated by alternative splicing.
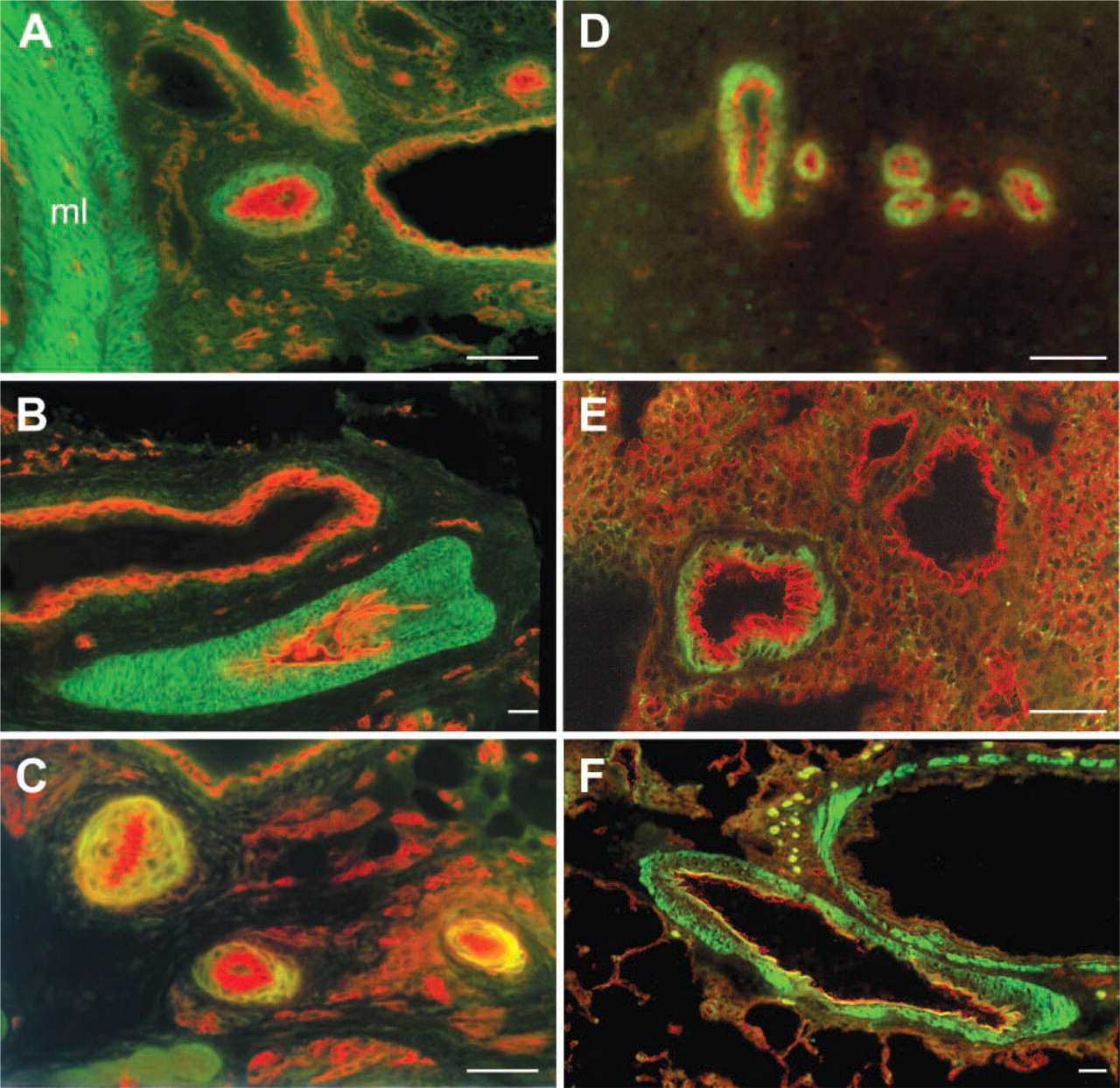
Single-labeling immunofluorescence of the ETB receptor. In the mesenteric (
Terminal arteries and arterioles play a central role in the regulation of total vascular resistance and blood flow distribution in different organs. About half of total peripheral resistance is regulated at this level. In all tissues of adult rats examined, we found ETB receptor immunoreactivity in the media of these vessels. In larger arteries and veins, however, there were marked tissue-specific differences concerning the presence of ETB receptors. In the mesenteric and skeletal muscle vascular beds, ETB receptor immunoreactivity was evident exclusively in the media of arterioles, whereas venules were devoid of ETB receptors. In mesenteric vessels, ETB receptors were restricted to smooth muscle cells of intramural arterioles. This explains why pharmacological studies performed on isolated mesenteric arteries were unable to detect a contribution of the ETB receptor subtype to the vasoconstriction induced by ET-1 (Rizzoni et al. 1997; Adner et al. 1998). In meningeal and intracerebral arteries and arterioles, the ETB receptor antiserum also labeled vascular smooth muscle cells. These findings are in contrast to those of previous reports in which ETB receptor immunoreactivity was detected in neuronal but not in vascular cells (Yamamoto and Uemura 1998) and ETB receptor mRNA in glial, ependymal, and plexus chorioideus cells (Hori et al. 1992).
Double-labeling immunofluorescence of the ETB receptor and the panendothelial cell marker RECA-1 at PD 0 and PD 5. No ETB receptor immunoreactivity can be observed in pulmonary (
In large conductive vessels, the media of the brachial artery and vein exhibited ETB receptor immunoreactivity, whereas carotid, renal, and mesenteric arteries were devoid of ETB receptors. The significance of these findings is difficult to interpret, inasmuch as comparative studies are missing, but tissue-specific regulation of vascular smooth muscle ETB receptors is obvious. In addition to its effect on vascular tone, ET-1 acting through the ETB receptor subtype has been demonstrated to contribute to mechanical stress-induced apoptosis of vascular smooth muscle cells (Cattaruzza et al. 2000; Lauth et al. 2000). These observations suggest that ET-1 could contribute to instability of atherosclerotic plaques by ETB receptor-mediated apoptosis of vascular smooth muscle cells.
In the pulmonary circulation, strong ETB receptor immunoreactivity was observed in the media of muscular segments of the pulmonary arteries and in pulmonary veins, whereas in elastic segments of pulmonary arteries, only scarce labeling of vascular smooth muscle cells was detected. These findings are in accordance with the report by Soma et al. (1999), who observed colocalization of ETA and ETB receptors on smooth muscle cells of small pulmonary arteries by immunohistochemistry and in situ hybridization. Functional data also demonstrated that ET-1-induced vasoconstriction in intraparenchymal arteries is mediated by both the ETA and the ETB receptor subtypes (McLean et al. 1994; Higashi et al. 1997). ETA and ETB receptors are known to be differentially coupled to G-proteins, and in small pulmonary arteries, levels of the cyclic nucleotides cAMP and cGMP were shown to be differentially regulated by either ETA or ETB receptor stimulation (Mullaney et al. 2000). In the rabbit pulmonary circulation, vasoconstriction induced by ET-1 shifted from the ETA to the ETB receptor subtype after preconstriction (Schmeck et al. 1999), suggesting that the contribution of ETB receptors to the vasoconstriction elicited by ET-1 depends on vascular tone. Our finding that ETB receptors are also present on smooth muscle cells of pulmonary veins is in line with ETB receptor-dependent increases in postcapillary vascular resistance reported by Uhlig et al. (1995).
Double-labeling immunofluorescence of the ETB receptor and the panendothelial cell marker RECA-1 at PD 14, PD 21, and PD 28. AT PD 14, vascular smooth muscle cells from the mesenteric arterioles (
In the coronary circulation, we observed ETB receptors in the media of all parts of the coronary arterial tree. ETB receptor immunoreactivity started immediately at the origin of the coronary artery from the aorta and continued until the terminal arterioles. While Hori et al. (1992) did not detect ETB receptor mRNA in coronary vessels by in situ hybridization, our findings are in line with the reports by Goodwin et al. (1999) and Balwierczak (1993), who demonstrated coronary vasoconstriction by ETA and ETB receptor stimulation. In a previous study, we reported that both ETA and ETB receptors colocalize on coronary arterial smooth muscle cells (Wendel-Wellner et al. 2002). The ETB receptor antiserum used in this study was directed against an epitope at the carboxy terminus of the ETB receptor molecule and also recognized ETB receptors on endothelial cells.
In the rat kidney, ETB receptor immunoreactivity was evident on vascular smooth muscle cells of arcuate and interlobular arteries as well as vas afferens and efferens. These findings are in accordance with functional data from different groups (Wellings et al. 1994; Endlich et al. 1996), demonstrating a significant role for ETB receptors in mediating ET-1-induced vasoconstriction in the rat kidney, partly by generation of vasoconstrictory eicosanoids (Hercule and Oyekan 2000). As in all other vascular beds studied, we did not observe endothelial labeling. Renal tubuli were also devoid of ETB receptor immunoreactivity. Previous studies employing immunohistochemistry and in situ hybridization did not identify ETB receptors on vascular smooth muscle cells of rat kidney. ETB receptor immunoreactivity was observed mainly in proximal tubuli and collecting ducts, whereas a weak signal was obtained over glomerular capillaries (Yamamoto and Uemura 1998). By in situ hybridization, Hocher et al. (1995) identified strong ETB receptor mRNA expression in medullary tubuli and glomerular capillaries. They found only a scattered signal for ETB receptor mRNA in endothelial cells, so the level of ETB receptor mRNA expression in vascular endothelial and smooth muscle cells may be very low. Despite the presence of ETB receptors in the arcuate arteries evidenced by immunofluorescence in our study, Wu et al. (1997) reported that the vasoconstriction induced by ET-1 was mainly dependent on the ETA receptor subtype.
During postnatal development, enhanced sensitivity of the intestinal (Nankervis and Nowicki 2000) and pulmonary vascular beds (Wang and Coceani 1992; Ivy et al. 1996) to the vasoconstricting effects of ET-1 has been reported, mediated mainly by the ETA receptor subtype. However, these studies were performed mainly in species known not to express ETB receptors on vascular smooth muscle cells. Our findings show that in the rat, ETB receptors on vascular smooth muscle cells develop postnatally in a tissue-specific manner and are present on vascular smooth muscle cells throughout the vascular system in the adult rat. ETB receptors are known to desensitize rapidly after agonist binding (Cramer et al. 1998), and repeated agonist exposure leads to a rapid decrease in the vasoconstriction elicited by ETB receptor-selective ligands (Sharifi and Schiffrin 1996). These characteristics make them ideal scavenging receptors, blunting the vasoconstrictor response of locally elevated levels of ET-1.
Taken together, we demonstrate that in the adult rat, ETB receptors are present on smooth muscle of resistance and exchange vessels in all tissues examined, whereas they are differentially distributed in conductive and venous vessels. During postnatal development, ETB receptors on vascular smooth muscle are regulated in a time- and tissue-specific manner.
Footnotes
Acknowledgements
This work was supported by a grant from the Deutsche Forschungsgemeinschaft (DFG Ko 1814/2).
The authors wish to thank Elke Richter, Karola Michael, Tamara Papadakis, and Martin Bodenbenner for technical assistance.