
Abstract
Endothelial nitric oxide synthase (eNOS), the major nitric oxide (NO)-generating enzyme of the vasculature, is regulated through multiple interactions with proteins, including caveolin-1, Hsp90, Ca2+-calmodulin, and the recently discovered eNOS-interacting protein, NOSIP. Previous studies indicate that NOSIP may contribute to the intricate regulation of eNOS activity and availability. Because eNOS has been shown to be abundantly expressed in the airways, we determined the expression and cellular localization of NOSIP in rat trachea and lung by RT-PCR and immunohistochemistry and examined the interaction of NOSIP with eNOS in lung by coimmunoprecipitation. In tracheal epithelium and lung, NOSIP mRNA expression was prevalent, as shown by RT-PCR, and the corresponding protein interacted with eNOS, as demonstrated by coimmunoprecipitation. Using immunohistochemistry, we found both NOSIP and eNOS immunoreactivity in ciliated epithelial cells of trachea and bronchi, while Clara cells showed immunoreactivity for NOSIP only. NOSIP and eNOS were present in vascular and bronchial smooth muscle cells of large arteries and airways, whereas endothelial cells, as well as bronchiolar and arteriolar smooth muscle cells, exclusively stained for NOSIP. Our results point to functional role(s) of NOSIP in the control of airway and vascular diameter, mucosal secretion, NO synthesis in ciliated epithelium, and, therefore, of mucociliary and bronchial function.
T
NOSIP is widely distributed in the cardiovascular system (Dedio et al. 2001), the gastrointestinal tract (König et al. 2002), and the nervous system of the rat (Dreyer et al. 2003), where it colocalizes with eNOS and/or nNOS. NOSIP coimmunoprecipitates with NOS from lysates of the rat small intestine (König et al. 2002) and heart (unpublished data), suggesting that NOSIP interacts with NO synthases in vitro and in vivo. In the rat and human lung, eNOS is present in endothelial and epithelial cells (Xue et al. 1996; Rizzo et al. 1998; Steudel et al. 1999; Zhan et al. 1999), where it is thought to have a major impact on lung disease, such as pulmonary hypertension (Hampl and Herget 2000), and exerts a bronchoprotective effect by converting the direct effect of substances such as acetylcholine and bradykinin on bronchial smooth muscle from bronchoconstriction to bronchodilatation through the release of NO from the epithelium (Ricciardolo et al. 2004). The mRNA for NOSIP has also been detected in human lung (Dedio et al. 2001). Given that NOSIP is a prime candidate for an endogenous modulator of eNOS function, we examined the cellular distribution of NOSIP and eNOS in the rat lower airways and lung and probed for the presence of the corresponding complex in coimmunoprecipitation experiments. Our results indicate that NOSIP and eNOS are coexpressed in distinct cell types of the lung and the trachea, suggesting a functional role for NOSIP in regulating NO synthesis and delivery in the airway system.
Materials and Methods
Chemicals
All chemicals were obtained from Sigma, Deisenhofen, Germany, unless otherwise stated.
Animals
Wistar rats (200-300 g) were used throughout. The animals were kept under standard laboratory conditions, and all procedures were performed according to National Institutes of Health guidelines.
Tissue Preparation
Wistar rats of both sexes were sacrificed by inhalation of halothane (Rüsch; Böblingen, Germany). To obtain unfixed tissue for immunohistochemical analyses, the lungs were inflated via the trachea using optimal cutting temperature (OCT) compound (Sakura; Zoeterwoude, The Netherlands). Lungs and trachea were jointly removed, oriented in OCT compound on a piece of filter paper, and snap-frozen in melting isopentane (n=5). For fixed tissues, the animals (n=5) were sacrificed, the thorax was opened, a cannula was inserted in the left atrium for retrograde perfusion of the lung, and the auricle of the right atrium was opened. Blood was removed by rinsing with a solution containing 2.5% polyvinylpyrrolidone and 0.5% procainamide-HCl (Forssmann et al. 1977), followed by 300 ml of Zamboni's fixative (Stefanini et al. 1967). The lungs were inflated with the same fixative; the trachea and lung were removed en bloc, washed repeatedly in 0.1 M phosphate buffer (PB), pH 7.4, immersed in 20% sucrose in PB, oriented in OCT compound on filter paper, and snap-frozen in liquid nitrogen. Sections 10 μm thick were cut with a Leica CM-1900 cryostat (Leica; Bensheim, Germany) and mounted on slides.
For Western blotting and RT-PCR, unfixed tissue was removed (n=3), snap-frozen in liquid nitrogen, and stored at −80C until use. For isolation of RNA from tracheal epithelium, the trachea was removed and cut open, and the epithelium was abraded (n=3). For Western blotting, unfixed tissue was ground under liquid nitrogen and directly lysed in boiling sample buffer (200 mM Tris-HCl, pH 6.8, 6% SDS, 20% glycerol, 10% DTT 0.1 mg/ml bromphenol blue). For Western blotting, a volume equivalent to 1 mg of tissue was applied.
Immunohistochemistry
Nonspecific binding of primary antibodies was blocked for 1 hr at room temperature by 50% normal swine serum (acetone-fixed sections) or 10% normal swine serum containing 0.5% Tween 20 and 0.5% bovine serum albumin (for Zamboni-fixed sections), followed by overnight incubation with a rabbit antiserum against a fusion protein of glutathione S-transferase (GST) and NOSIP at 1:800 (Dedio et al. 2001) in PBS with doubled salt concentration (PBS+S). After washing with PBS, the sections were incubated for 1 hr with Cy3-conjugated donkey anti-rabbit immunoglobulin (Ig) (Dianova; Hamburg, Germany) diluted 1:1000 in PBS+S. After a final washing step with PBS, the sections were cover-slipped with carbonate-buffered glycerol, pH 8.6. To test for specificity, anti-NOSIP was preincubated overnight with 20 μg/ml of the authentic antigen, i.e., GST-NOSIP. All labeling reported here was specific, as judged by its absence in sections incubated with preabsorbed serum.
To discriminate between different epithelial cell types, double-labeling experiments for NOSIP and specific cell-type markers were performed (Table 1). When mouse monoclonal antibodies were applied, FITC-labeled donkey anti-mouse Ig was used as the secondary antibody. For eNOS detection, mouse monoclonal antibody to eNOS (eNOSm) (clone 3; Transduction Laboratories, Lexington, KY) was used at 1:150, or a polyclonal rabbit antibody (eNOSp) to synthetic peptide NH2-PYNSSPRPEQHKSYKC-COOH (residues 599-613 of eNOS) with an extra Cys residue (underlined) for coupling to keyhole limpet hemocyanin was used at 1:400 (Dedio et al. 2001). As secondary antibodies, Cy3-conjugated donkey anti-rabbit Ig or anti-mouse Ig (Dianova) were used at 1:1000. For double labeling, mono- and polyclonal antibodies to eNOS were applied, followed by Cy3-conjugated donkey anti-rabbit Ig and FITC-conjugated donkey anti-mouse Ig, or the same antibodies with swapped labels at 1:400 throughout (Dianova). For control, eNOSp was preincubated overnight at 4C with 100 μg/ml of the corresponding antigen, and all labeling reported for eNOSp was specific, as judged by its absence in sections incubated with preabsorbed antibody.
Mouse monoclonal antibodies used for double-labeling experiments
NADPH Diaphorase Reaction
NADPH diaphorase reaction was utilized to localize NOS by its enzymatic activity (Beesley 1995). Cryosections of snap-frozen tissue were cut, mounted, and fixed in acetone as detailed above. After drying, sections were fixed with 4% paraformaldehyde for 20 min, washed three times in PBS, and incubated with 1 mg/ml NADPH, 0.1 mg/ml nitroblue tetrazolium (both from Biomol; Hamburg, Germany), 0.3% Triton X-100 in PB at 37C for 60 min. To stop the reaction, the slides were washed three times for 10 min each in buffer, and coverslipped with carbonate-buffered glycerol, pH 8.6.
Database Search and RT-PCR
To identify the yet-unknown rat NOSIP cDNA sequence, we used mouse NOSIP cDNA and protein sequences (gi: 12845151) and compared them using the BLAST program (http://www.ncbi.nlm.nih.gov/BLAST/) to expressed sequence tag (EST) databases recruiting data from non-mouse, non-human sequences of GenBank, EMBL, and DDBJ. Two independent yet overlapping EST sequences from rat (gi: 11633451, gi: 20420873) were combined to construct a putative rat NOSIP cDNA sequence. This sequence was used to design a primer pair (forward: 5′-GTCGTTTTTGAGGCGAGTTC-3′; reverse: 5′-GCCTGTTCACGTCCAAGTTT-3′) for RT-PCR to amplify a 1053 bp product. Sequence identity between human and rat proteins was quantified by BLAST, and the alignment of the two sequences was optimized by ClustalW 1.83 software.
Total RNA from lung and abraded tracheal epithelial cells was isolated using the RNeasy method according to the manufacturer's protocol (Qiagen; Hilden, Germany). Contaminating DNA was degraded using 1 U Dnase-I (Invitrogen; Karlsruhe, Germany) per μg of total RNA, and reverse transcription was done for 50 min at 42C using 200 U Superscript II reverse transcriptase (Invitrogen) per μg of RNA. The cDNA was amplified by adding 4 pmol of forward and reverse primer each, 2 U AmpliTaq Gold (Applied Biosystems; Darmstadt, Germany), 4 μl of 10 × PCR Buffer II (Applied Biosystems), 4 mM MgCl2 (Applied Biosystems), 0.02 mM dNTP each (Amersham Biosciences; Freiburg, Germany), and sterile water in a final volume of 50 μl. The cycling protocol was 10 min at 94C, followed by 35 cycles of 45 sec at 95C, 45 sec at 60C, and 45 sec at 73C, and by a final elongation step for 7 min at 73C. Sequencing of the PCR product revealed the total identity of the amplified product with the EST-based predictions of rat NOSIP cDNA (see above).
Coimmunoprecipitation
Frozen tissue was homogenized and lysed for 1 hr on ice in 20 mM Tris-HCl, 150 nM NaCl, 1 mM EDTA, 20 mM 3-[(3-cholamidopropyl)-dimethylammonio]-1-propanesulfonate, and complete protease inhibitor mixture (Boehringer; Mannheim, Germany), and centrifuged at 14,000 × g for 10 min at 4C. Ten μl/ml rabbit anti-NOSIP or anti-eNOSp was added to the supernatant, and the samples were incubated for 1 hr at 4C under rotation. For control, preimmune serum was used to replace rabbit anti-NOSIP. Antibodies were bound to Pansorbin cells (Calbiochem; Nottingham, UK) and precipitated at 8000 × g for 2 min at 4C. The precipitates were washed four times and dissolved in sample buffer (63 mM Tris-HCl, pH 6.8, containing 2.5% SDS, 5% glycerol, and 0.005% bromphenol blue).
Recombinant Expression and Purification of His(6)-tagged NOSIP
The murine NOSIP cDNA sequence was cloned into the vector pQe9 (Qiagen), fusing the protein to an N-terminal His(6)-Tag, and expressed in Escherichia coli SG13009 (Qiagen). Recombinant protein was purified by affinity chromatography on an Ni-NTA column under denaturing conditions as described previously (Oess and Hildt 2000) followed by ion-exchange chromatography using a MonoS column (Amersham Biosciences) and dialysis overnight against PBS. Purified protein was dissolved in sample buffer (200 mM Tris-HCl, pH 6.8, 6% SDS, 20% glycerol, 10% DTT 0.1 mg/ml bromphenol blue), and 1 μg was used for Western blotting.
Western Blotting
Samples were run on 10% SDS-PAGE, followed by electrotransfer of the resolved proteins to nitrocellulose membrane. Immunoblotting was done either with anti-eNOSm antibody at 1:1000 or rabbit anti-NOSIP at 1:1000, a horse-radish-peroxidase-bound goat anti-mouse or anti-rabbit Ig at 1:3000 (Bio-Rad; Munich, Germany), and a chemiluminescence detection assay (Amersham Biosciences).
Results
Identification and Comparison of the Rat NOSIP Sequence
Using the human NOSIP sequence, we identified by BLAST searches of public databases two overlapping EST tags and used them to construct a hypothetical rat NOSIP sequence. A primer pair derived from this sequence was employed for RT-PCR using total RNA from whole lung or from abraded tracheal epithelium of rat (Figure 1A). Sequence analysis of the resultant amplification product of 1053 bp revealed total identity with the hypothetical EST-based coding sequence of rat NOSIP (Figure 2). The identified cDNA sequence fully matches the corresponding DNA sequence of rat chromosome 1 (GenBank, gi: 27731402). The derived amino acid sequence codes for a protein of 301 amino acid residues that shares 91% (98%) identity with the human (mouse) NOSIP sequence. Alignment of the human and rat sequences indicates that the N-terminal, central, and C-terminal portions are well conserved, whereas the intervening sequences (positions 130 to 142 and 199 to 207) diverge considerably (48% and 56% identity, respectively) between the two species (cf. Figure 2). This finding is in good agreement with the previous observation of a tripartite structure of NOSIP (Dedio et al. 2001) where the terminal and central regions are well conserved, whereas the intervening sequences vary significantly (positions 92 to 152 and 204 to 219).
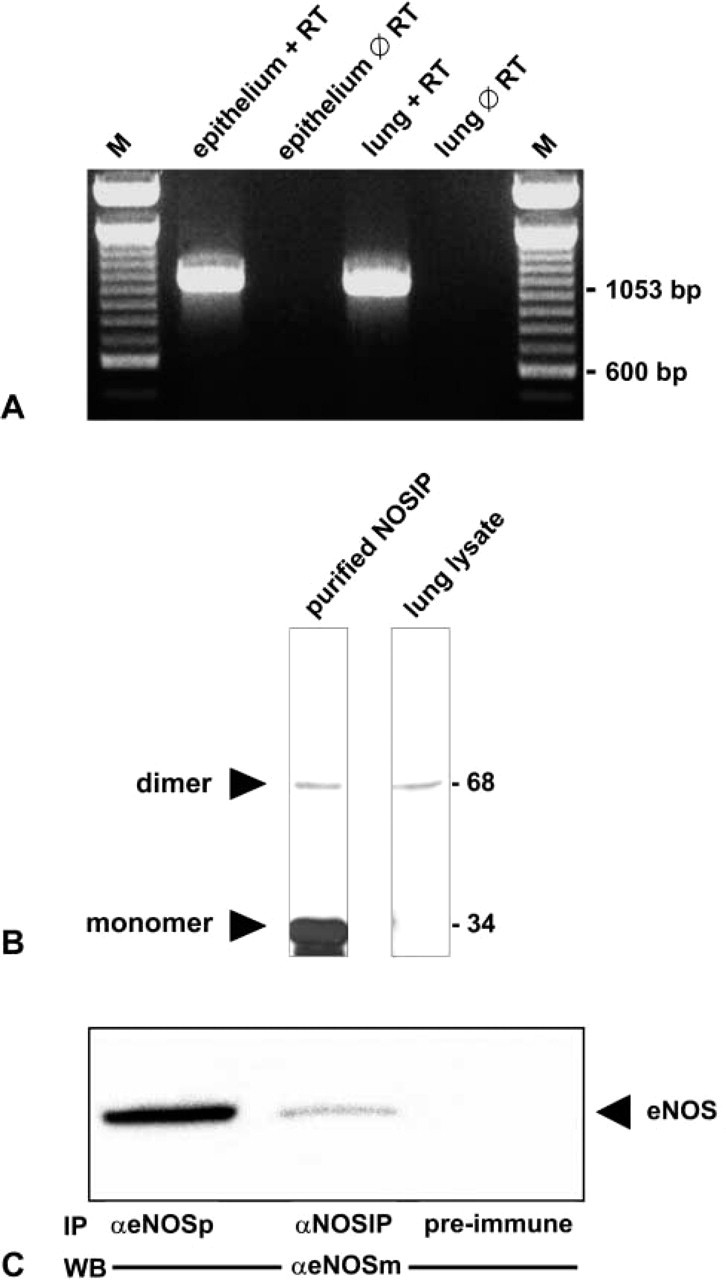
NOSIP is expressed and interacts with eNOS in lung. (
Western Blotting of Lung Homogenates and Comparison to Recombinant NOSIP
Given the high degree of sequence identity between human and rodent NOSIP, we used a well-characterized antibody to human NOSIP (Dedio et al. 2001) to test for the expression of the corresponding protein by Western blotting. A single band of 68 kD was detected in lung homogenates (Figure 1B, lane 2), indicating that in the lung, the NOSIP dimer is the predominant form of NOSIP, as previously reported for gastrointestinal tissues (König et al. 2002). Recombinant NOSIP shows a band at 34 kD and a weaker band at 68 kD (Figure 1B, lane 1), indicating that recombinant NOSIP is predominantly monomeric but also capable of dimer formation.
Coimmunoprecipitation of NOSIP and eNOS from Lung Homogenates
To probe for the presence of a NOSIP-eNOS complex, we used anti-NOSIP for indirect immunoprecipitation of eNOS, followed by Western blotting with a monoclonal antibody to eNOS (Figure 1C). Approximately 5% of total eNOS was associated with NOSIP in whole lung homogenates, as judged from the comparison with the results from direct immunoprecipitations with polyclonal anti-eNOS (Figure 1C). Failure of the corresponding preimmune serum to coprecipitate eNOS demonstrated the specificity of the antibody. We conclude that at least a fraction of eNOS present in lung homogenates is complexed with NOSIP. Owing to the harsh solubilization conditions applied, this value (5%) probably reflects the lower limit of the complexed fraction.
Immunohistochemical Detection of NOSIP win Rat Airways
In the trachea and in bronchi, NOSIP immunoreactivity was found in the epithelium and in the smooth muscle cells. In bronchial arteries, NOSIP immunoreactivity was confined to vascular smooth muscle cells (Figures 3A-3C). Only occasionally, single endothelial cells were NOSIP immunoreactive. In pulmonary veins, NOSIP immunoreactivity was found in cardiac myocytes and in the microvascular endothelium in the venous wall (Figure 3D). NOSIP immunoreactive endothelial cells occurred more frequently in the pulmonary vein than in bronchial arteries. NOSIP immunoreactivity was also found in chondrocytes of the tracheal cartilage (Figure 2A). In the alveolar region, NOSIP immunoreactivity could not be detected in alveolar epithelium or in microvascular endothelial cells (Figure 3C).

Sequence alignment of rat and human NOSIP. Identical amino acids (∗), conservative exchanges (:) and semi-conservative exchanges (.) are indicated.
Identification of NOSIP-positive Epithelial Cell Types
Throughout the bronchial tree, we found NOSIP immunoreactivity associated with ciliated cells, identified by their typical morphology and by immunolabeling with a monoclonal antibody to eNOS (Xue et al. 1996) (Figure 4A). Unlike ciliated cells, brush cells labeled with anti-villin (Kasper et al. 1994) and neuroepithelial cells or nerve fibers identified with anti-PGP (protein gene product) 9.5 (Lauweryns and Van Ranst 1988) were not labeled by the NOSIP antiserum (Figures 4D and 4E). In the epithelium of the trachea and the main bronchi, NOSIP immunoreactivity was conspicuously absent from secretory cells labeled with anti-surfactant protein D (Kasper et al. 2002) (Figure 4C), whereas surfactant protein-positive Clara cells of the smaller airways (Crouch et al. 1992; Kasper et al. 2002) were also positive for NOSIP (Figure 4B).
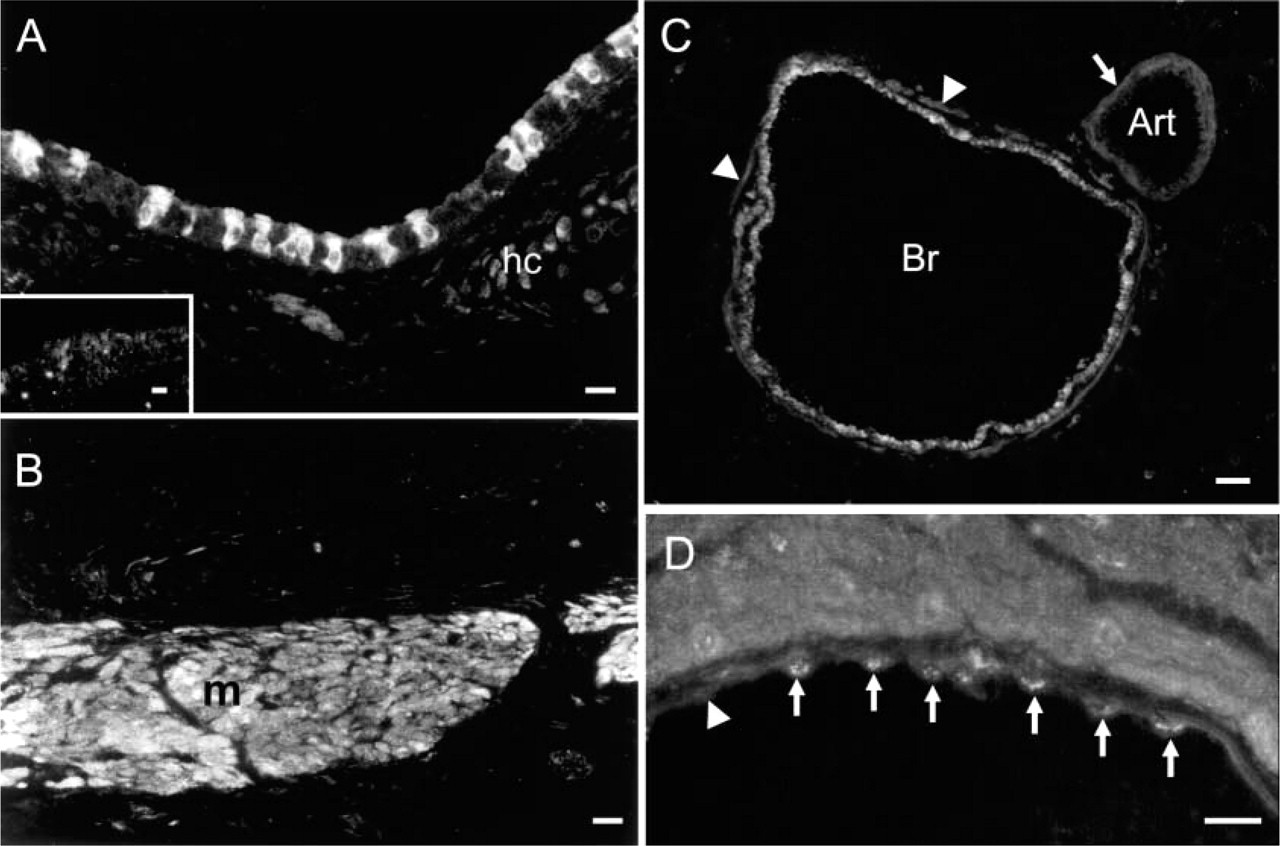
Immunohistochemistry of NOSIP in rat trachea and lung. (
Differential Distribution of eNOS in the Lower Airways and Lung
To compare the distribution of NOSIP immunoreactivity with the pattern of its interaction partner eNOS, we employed two distinct antibodies, i.e., a monoclonal antibody (eNOSm) to the C terminus of human eNOS and a polyclonal affinity-purified antibody (eNOSp) raised to positions 599 to 613 of bovine eNOS (Dedio et al. 2001). Antibody eNOSm labeled endothelial cells of small vessels in the connective tissue of trachea, bronchial arteries and veins, and capillaries present in the cardiac muscle coat of pulmonary veins (Figure 5, center panel). No labeling was seen in the microvascular endothelium of alveolae. In the bronchial epithelium, eNOSm selectively labeled a thin apical area of ciliated cells. Consonant with these observations, antibody eNOSp labeled endothelial cells of small vessels in the connective tissue of the trachea, bronchial arteries and pulmonary veins, and capillaries in the cardiac muscle coat of pulmonary veins (Figure 5, left panel). In addition, this antibody labeled cardiomyocytes in the wall of the pulmonary veins and revealed weak eNOS immunoreactivity in smooth muscle cells of the larger airways and bronchi (Figure 5, left panel). Unlike the eNOSm antibody, eNOSp labeled the bronchial epithelial cells only in larger airways (Figure 5, left panel). Double labeling of these sections demonstrated that both antibodies labeled the same ciliated epithelial cells, although with a slightly different pattern. Antibody eNOSp showed a punctate labeling localized basally to the eNOSm staining (Figure 5, right panel). Double labeling also revealed that eNOSp recognized endothelial cells that showed weak, if any, staining for eNOSm (Figure 5, right panel). Taken together, we identified overlapping patterns of NOSIP and eNOS immunoreactivity in ciliated epithelial cells, cardiomyocytes, smooth muscle cells of larger bronchi, and arteries, as well as in a minority of endothelial cells.
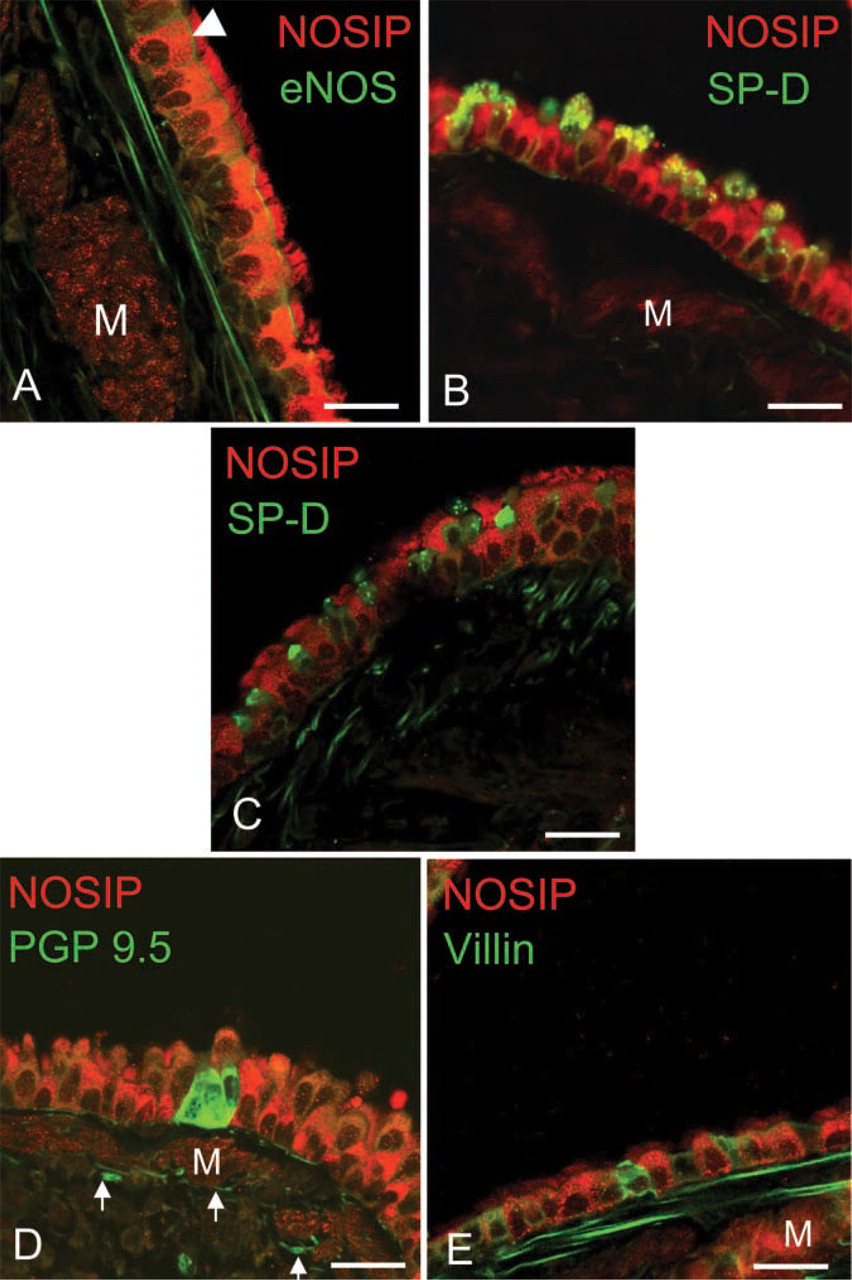
Double-labeling immunohistochemistry for NOSIP and epithelial markers in bronchi. (
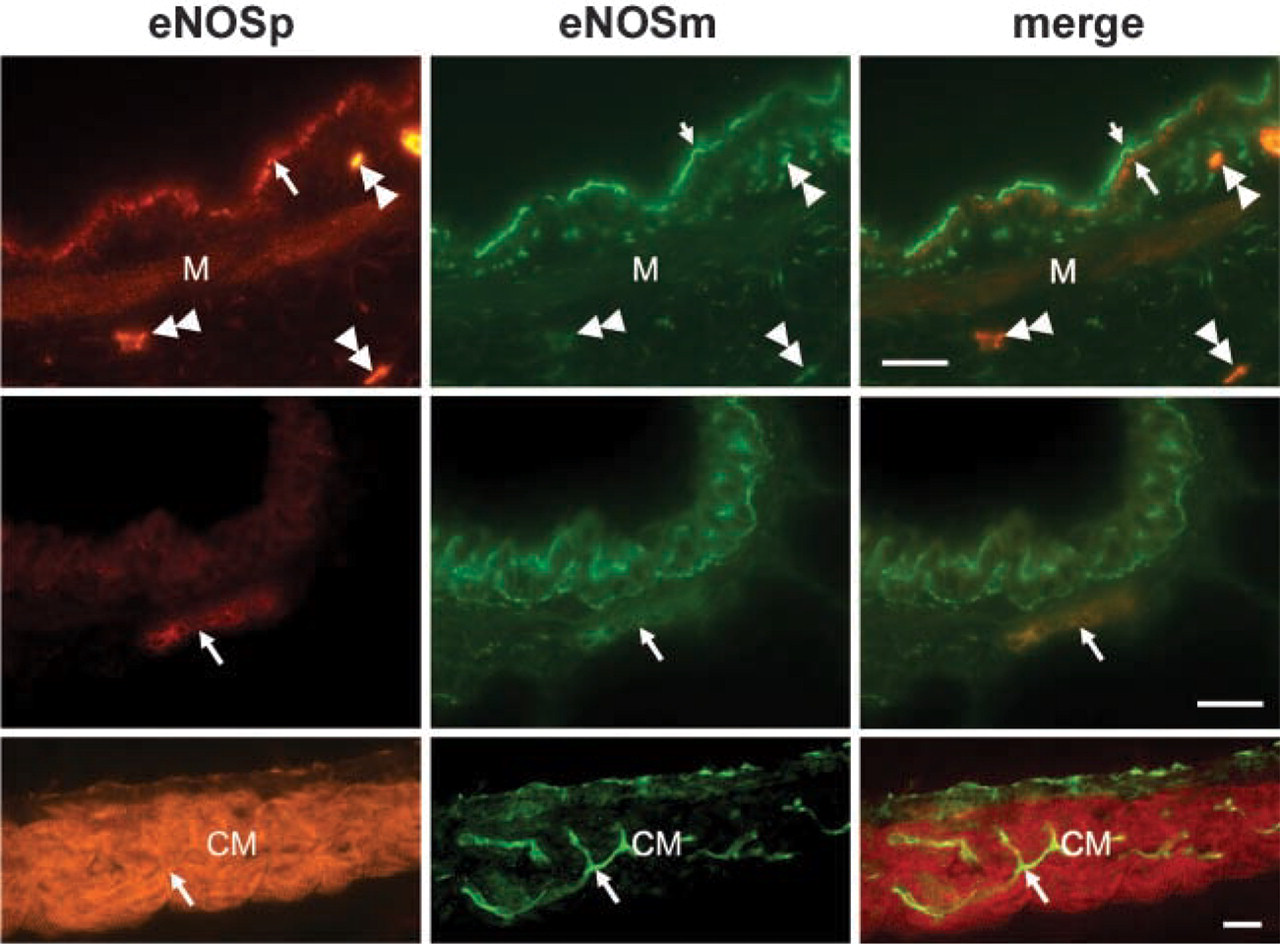
Double labeling with eNOSm and eNOSp in airways. (Top panel) In large airways, both antibodies label bronchial epithelial cells. eNOSm labels an area (small arrow) located apically to that marked by eNOSp (large arrow). Antibody eNOSp shows a weak labeling of bronchial smooth muscle (M); it labels capillary endothelium stronger than eNOSm (doubled arrowheads). (Center panel) In small bronchi, immunoreactivity for eNOSm is exclusively present in the epithelium, whereas the capillary (arrow) is labeled by both antibodies. (Bottom panel) In the pulmonary vein, eNOSp strongly labels cardiac muscle (CM), whereas eNOSm fails to stain cardiomyocytes. Both antibodies react with the endothelium of the pulmonary vein and the capillaries of the cardiac muscle sheath (arrow). Bars = 20 μm.
NADPH Diaphorase Reaction for NOS Activity
To underpin our immunohistochemical staining for eNOS, we employed the NADPH diaphorase reaction in various airway tissues and found reaction products indicative of NOS activity in the endothelium of arteries and veins, in cardiomyocytes that coat pulmonary veins, in airway epithelium, and in small cells of the alveolar region (Figures 6A-6C). A faint staining was also seen in the smooth muscle layer of larger arteries and bronchi, and, inconsistently, in the surrounding connective tissue (Figure 6B). The localization of NADPH diaphorase reaction fully matches the distribution of eNOS immunoreactivity with the exception of Clara cells that were NADPH diaphorase positive but were not immunoreactive for eNOS.
Discussion
Because NO is a key messenger in the body, NO generation and turnover are tightly controlled with respect to time and space (Dedio et al. 2001). Multilateral control of eNOS activity affects transcriptional activity, mRNA stability, posttranslational modifications, subcellular targeting, and reversible protein-protein interactions, e.g., via NOSTRIN (eNOS traffic inducer) (Zimmermann et al. 2002) and NOSIP (Dedio et al. 2001; König et al. 2002; Dreyer et al. 2003). The present study establishes the presence of NOSIP mRNA and protein in airways and provides evidence for its interaction with eNOS in the lung.
At the cellular level, we have probed colocalization of NOSIP and eNOS by double-labeling immunohistochemistry. Because the available eNOS antibodies tend to discriminate between various posttranslationally modified isoforms and even conformational states of eNOS (Bloch et al. 2001; König et al. 2002), we used two distinct antibodies with complementary labeling patterns that have previously been shown to identify the vast majority of eNOS-expressing cells, e.g., of the gastrointestinal tract (König et al. 2002) and in the heart (Bloch et al. 2001). In this way, we have been able to demonstrate the unequivocal colocalization of NOSIP and eNOS immunoreactivities in ciliated airway epithelial cells, in cardiomyocytes associated with pulmonary veins, and in some, but not all, endothelial cells. On the molecular level, our coimmunoprecipitation experiments provide direct evidence for the interaction of NOSIP and eNOS in the respiratory tract, in line with the data from a yeast two-hybrid screen that led to the identification of NOSIP (Dedio et al. 2001).
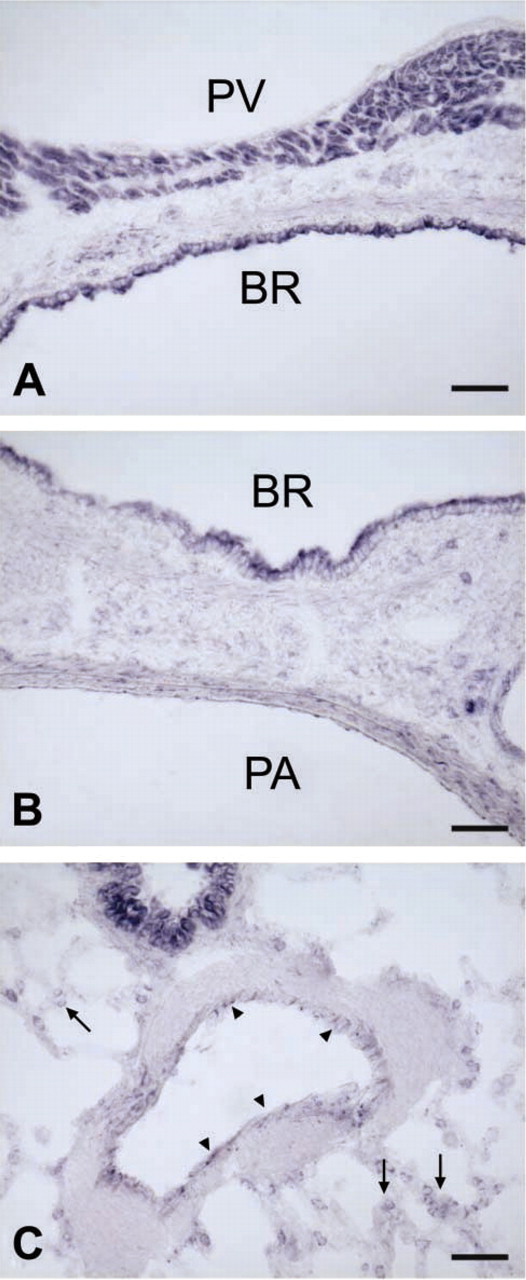
NADPH diaphorase reaction in the lung. (
Previous studies have demonstrated that ciliated epithelial cells of the airways produce NO, which in turn increases ciliary beat frequency (CBF) and induces bronchodilatation (Li et al. 2000; Ricciardolo 2003). Using distinct antibodies to eNOS, we were able to confirm the presence of eNOS in ciliated epithelial cells, although the two types of antibodies tended to label eNOS at distinct subcellular locations. This finding is in line with previous observations that both antibodies detect different modification states of eNOS, owing to the fact that they bind to different parts of the protein. Therefore, the conformational or phosphorylation states of eNOS or other bound proteins are thought to interfere with antibody binding (Bloch et al. 2001; König et al. 2002). Because eNOS and NOSIP colocalize in ciliated cells, we postulate that NOSIP is a prime candidate for modulating eNOS function. Data from the overexpression system suggest that NOSIP does not directly inhibit eNOS but is able to govern its subcellular localization (Dedio et al. 2001). Therefore, it might reduce eNOS activity by dragging the enzyme to areas of lower arginine availability. The change of subcellular localization of eNOS might also result in translocation from the apical part of the ciliated cell to a more basal localization. This could redirect the NO synthesis toward the bronchial smooth muscle cells. Both mechanisms would influence bronchial diameter and, through changes in CBF, the mucociliary clearance. The situation is less clear for Clara cells, which consistently showed NOSIP immunoreactivity and NADPH diaphorase positivity but failed to show eNOS immunoreactivity with either of the two antibodies to eNOS used herein (Bloch et al. 2001; König et al. 2002). Other authors have, however, demonstrated eNOS immunoreactivity in rat Clara cells (Zhan et al. 1999) as well as in human NCI-H441 cells thought to be derived from Clara cells (Shaul et al. 1994). Therefore, it is quite possible that strain differences and/or unusual eNOS modifications have prevented us from successfully detecting eNOS immunoreactivity in Clara cells. Hence, at this time we cannot draw firm conclusions as to the potential role(s) of NOSIP in Clara cells.
As has already been demonstrated for the gastrointestinal tract (König et al. 2002), smooth muscle cells are a major source of NOSIP immunoreactivity. In larger airways and arteries, these cells also show weak though significant eNOS immunoreactivity as well as NADPH diaphorase activity. In line with these observations, eNOS mRNA and protein have been found by RT-PCR, immunohistochemistry, and Western blotting in smooth muscle cells of diverse origins (Murthy et al. 1998; Teng et al. 1998; Buchwalow et al. 2002). For instance, in rabbit, gastric smooth muscle eNOS was shown to be functionally active and stimulatable via the natriuretic peptide clearance receptor (Murthy et al. 1998). In vascular smooth muscle cells, NO production was demonstrated by the fluorescent NO indicator DAF-2DA (Brown et al. 1999; Haberberger et al. 2003). Also, eNOS immunoreactivity was present in smooth muscle cells associated with the central pulmonary vessels of rat lung (Steudel et al. 1999). Thus, NOSIP and eNOS coexist in at least a subpopulation of pulmonary smooth muscle cells and, although we have not directly demonstrated their physical interaction, it is likely that NOSIP serves a distinct regulatory role in the NO production of these cells.
In view of the promiscuous interactions of eNOS with various cytosolic and membrane-associated proteins such as Ca2+-calmodulin, Hsp-90, protein kinase B/Akt, and caveolin-1 that influence its activity as well as its subcellular localization but also serve a wide array of interactions far beyond eNOS binding (Pratt 1997; Lawlor and Alessi 2001; Razani et al. 2002), one may expect that a widely expressed protein such as NOSIP has eNOS-independent functions, e.g., in bronchial and vascular smooth muscle cells lacking demonstrable eNOS. One possibility is that NOSIP is functionally linked to nNOS, as is the case in neuronal cells of the dorsal spinal horn, where NOSIP appears to modulate NO homeostasis in pathological pain conditions (Dreyer et al. 2003). Alternatively, NOSIP may interact with yet unknown partner proteins; our present experimental efforts are directed toward the identification of non-NOS proteins targeted by NOSIP. Finally, one should consider the possibility that NOSIP may function in smooth muscle cells of smaller vessels under conditions that induce the expression of eNOS, for example, under hyperoxic conditions (Steudel et al. 1999).
In summary, we conclude that the coexistence of eNOS and NOSIP in ciliated epithelial cells, in a subset of smooth muscle and endothelial cells, and possibly in Clara cells of the rat lung support the notion that NOSIP may modulate eNOS activity and location, and that the presence of NOSIP in smooth muscle cells apparently devoid of eNOS may point to roles of NOSIP independent of its eNOS-binding capacity.
Footnotes
Acknowledgements
This study was supported by a Young Scientist's grant from the Justus-Liebig-Universität (to PK), the Graduiertenkolleg 534 (to WK), and the Sonderforschungsbereich 553 of the Deutsche Forschungsgemeinschaft (to SO and WM-E).
We thank Karola Michael for expert help with the figures.