
Abstract
Apolipoprotein AI (apoAI), a major component of high-density lipoproteins, is one of the major amyloid fibril proteins and a minor constituent of the senile plaques observed in Alzheimer's disease. We examined colocalization of apoAI in various kinds of systemic amyloidosis in this study. Forty-three of 48 formalin-fixed paraffin-embedded heart specimens with various forms of systemic amyloidosis reacted immunohistochemically with anti-human apoAI antibody. ApoAI was also detected in water-extracted amyloid material by immunoblotting. In addition, we observed colocalization of apoAI and murine amyloid A (AA) amyloidosis in human apoAI transgenic mice. This is the first report of colocalization of apoAI with amyloid deposits in various forms of human systemic amyloidosis and murine AA amyloidosis in human apoAI transgenic mice. ApoAI may not always be a major component of amyloid fibrils, even when it is present in systemic amyloid deposits.
Keywords
A
Apolipoprotein AI (apoAI) is a major component of high-density lipoproteins (HDL) (Eisenberg 1984; Coetzee et al. 1986). It is synthesized in both the liver and small intestine as a 267-amino-acid (aa) prepropeptide. The prepropeptide is cleaved intracellularly to the 249-aa propeptide and secreted into the plasma. Several studies have examined the association between apoAI and amyloidosis (Wisniewski et al. 1995a, b; Genschel et al. 1998). Several forms of amyloidosis, including hereditary systemic amyloidosis (Booth et al. 1996), hereditary cutaneous and cardiac amyloidosis (Hamidi Asl et al. 1999), and amyloidosis in atherosclerotic plaques of the human aorta (Westermark et al. 1995; Mucchiano et al. 2001a, b), are caused by apoAI variants. One form of amyloidosis in aortic atherosclerotic plaques is caused by wild-type apoAI (Mucchiano et al. 2001a). Canine pulmonary vascular amyloidosis is also caused by apoAI (Roertgen et al. 1995). In addition, it has been reported that apoAI colocalizes with senile plaques in patients with Alzheimer's disease (Wisniewski et al. 1995a; Harr et al. 1996).
In the present study we examined colocalization of apoAI in various human systemic amyloidosis immunohistochemically and biochemically. In addition, we examined association of apoAI with murine AA amyloidosis in human apoAI transgenic mice.
Materials and Methods
Immunohistochemical Analysis of ApoAI in Human Systemic Amyloidosis
Specimens were obtained at autopsy from 25 hearts with AA amyloidosis, 12 with immunoglobulin (Ig) λ light chain amyloidosis (Aλ amyloidosis), 5 with Igκ light chain amyloidosis (Aκ amyloidosis), 3 with Aβ2M amyloidosis, and 3 with ATTR amyloidosis. The cases had all been classified previously by immunohistochemical studies with antibodies against anti-λ (118-134), anti-κ (116-133) (Hoshii et al. 2001), and antibodies against AA protein (Imada 1981), Aλ, Aκ, transthyretin, and β2-microglobulin. Antibodies against Aλ and Aκ were kindly provided by Dr. George G. Glenner (University of California, San Diego). Formalin-fixed, paraffin-embedded tissue sections (4-μm thick) were stained with Congo red and examined by conventional and polarized microscopy. Immunohistochemical analysis of apoAI was performed as follows. After deparaffinization, sections were treated with 3% H2O2 to block endogenous peroxidase activity and with Protein Block Serum-Free (DAKO; Carpinteria, CA) to block nonspecific reactions. Affinity-purified goat anti-human apoAI antibody (1:200) (Harlan Sera-Lab; Leicestershire, UK) was applied as the primary antibody for 30 min at room temperature. According to the product information, cross-reactivity of this antibody with apolipoprotein AII, B, CI, CII, CIII, E, and J is less than 1%. Peroxidase-conjugated rabbit anti-goat immunoglobulin (1:50) (DAKO; Glostrup, Denmark) was used as the secondary antibody (30 min at room temperature) and visualized with 3-3′-diaminobenzidine tetrahydrochloride (DAB). Some sections were immunostained with or without formic acid pretreatment as part of a preliminary study. Formic acid pretreatment improved apoAI immunoreactivity; therefore, antigen retrieval was performed in 100% formic acid for 1 min at room temperature (Kitamoto et al. 1987). The specificity of apoAI antibody was confirmed by preabsorption test as follows. Purified human apoAI (100 μg/ml) (Chemicon; Temecula, CA) was mixed with an equivalent volume of goat anti-human apoAI antibody (1:100) (Harlan Sera-Lab). The mixture was incubated for 24 hr at 37C. After centrifugation at 10,000 × g for 20 min, the supernatant was used as the primary antibody in immunohistochemistry experiments. Immunohistochemical staining for apoAI in the representative cases and preabsorption test were also performed with a different goat anti-human apoAI antibody (Chemicon).
Immunoblotting of Human AL Amyloidosis
For immunoblotting experiments, Aλ amyloid was extracted from the spleen of a patient with primary systemic amyloidosis by the water-extraction method (Pras et al. 1969; Imada 1981). Aλ amyloid deposition was confirmed in a tissue section by Congo red and immunohistochemical staining. The water-extracted material was dissolved in 8 M urea, mixed with Laemmli sample buffer (Bio-Rad; Hercules, CA) containing 5% 2-mercaptoethanol (Sigma; St Louis, MO), and heated at 99C for 6 min. The sample was loaded onto a 15-20% gradient gel (Daiichi Pure Chemicals; Tokyo, Japan) for sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transblotted onto polyvinylidene difluoride (PVDF) membranes (Millipore; Billerica, MA). Nonspecific reactions were blocked with 5% skim milk, and the membrane was then incubated with affinity-purified goat anti-human apoAI antibody (1:4000) (Harlan Sera-Lab) for 1 hr at room temperature. Membranes were then incubated with peroxidase-conjugated rabbit anti-goat immunoglobulins (1:1000) (DAKO) for 1 hr at room temperature and visualized with True Blue (KPL; Gaithersburg, MD). As a positive control, purified human apoAI (Biogenesis; England, UK) was resolved on the same SDS-PAGE gel. Immunostaining with anti-λ (118-134) was also performed to confirm whether the sample contained Aλ amyloid protein.
To increase confidence in the specificity of the anti-apoAI antibody by examining whether this antibody cross-reacts with other apolipoproteins and amyloid-associated proteins, immunoblotting of apoE (Calbiochem; La Jolla, CA), serum amyloid A (SAA) (American Research Products; Belmont, MA), and serum amyloid P-component (SAP) (Techne; Minneapolis, MN) with affinity-purified goat anti-human apoAI antibody (Harlan Sera-Lab) was also performed.
Immunohistochemical Analysis of ApoAI in Human apoAI Transgenic Mice
ApoAI transgenic mice [C57BL/6-TgN(APOA1)1Rub] (Rubin et al. 1991) were purchased from Jackson Laboratory (Bar Harbor, ME). For the first experiment, AA amyloidosis was induced in the mice according to the method of Ram et al. (1968) but with some modifications. Briefly, a mixture (0.25 ml) of Freund's complete adjuvant (FCA) and Mycobacterium butyricum was injected subcutaneously into three mice. A second subcutaneous injection of the FCA-M. butyricum mixture (0.25 ml) was administered 2 weeks later. These mice were killed under ether anesthesia 2 weeks after the second injection. Spleens were isolated, fixed in 10% formalin, and embedded in paraffin.
For the second experiment, AA amyloidosis was induced in mice by injecting amyloid-enhancing factor (AEF) and AgNO3 to evaluate the earliest stage of amyloid deposition. AEF was prepared from ICR mice according to the method of Axelrad et al. (1982) but with some modifications as described previously (Hoshii et al. 1997). In three human apoAI transgenic mice, 0.5 ml of AEF was administered intraperitoneally, and 0.5 ml of 2% AgNO3 was administered subcutaneously. Two of these mice were killed under ether anesthesia 72 hr after administration of AEF and AgNO3. Spleens were isolated, fixed in 10% formalin, and embedded in paraffin.
For both experiments, sections (4-μm thick) were cut from the paraffin-embedded tissues, stained with Congo red, and examined by conventional and polarized light microscopy. Immunohistochemical examinations were performed with an indirect immunoperoxidase method as described for human tissues. Rabbit anti-mouse AA (1:1000) (Imada 1981) or affinity-purified goat anti-human apoAI (1:200) (Harlan Sera-Lab) was applied as the primary antibody for 30 min at room temperature. Peroxidase-conjugated goat anti-rabbit immunoglobulin (1:20) (DAKO) or peroxidase-conjugated rabbit anti-goat immunoglobulin (1:50) (DAKO) was applied as the secondary antibody for 30 min at room temperature. Anti-human and anti-mouse apoAI antisera showed <1% cross-species reactivity (Rubin et al. 1991); therefore, polyclonal anti-human apoAI antibody did not react significantly with murine apoAI. These experiments were carried out under the control of the Guidelines for Animal Experiments of Yamaguchi University School of Medicine.
Results
Histological and Immunohistochemical Findings in Human Systemic Amyloidosis
Congo red-positive amyloid deposits were present in the vessel walls in all cases. Amyloid deposits were also present in the interstitium of the myocardium, the endocardium, and the pericardium in most cases. Human apoAI immunoreactivity in amyloid deposits (Figure 1) was observed in 22 of 25 (88%) cases of AA amyloidosis, 12 of 12 (100%) cases of Aλ amyloidosis, 4 of 5 (80%) cases of Aκ amyloidosis, 2 of 3 (67%) cases of Aβ2M amyloidosis, and 3 of 3 (100%) cases of ATTR amyloidosis. Therefore, 43 (90%) of 48 cases of various kinds of systemic amyloidosis were positive for apoAI. In the positive cases, the immunoreactive area and staining intensity of amyloid deposits were variable. More cases showed partial anti-human apoAI immunoreactivity than showed complete anti-human apoAI immunoreactivity. Anti-human apoAI immunoreactivity with amyloid deposits was blocked completely by preabsorption. The immunoreactivity in the representative cases and the result of preabsorption test with anti-apoAI antibody (Chemicon) were similar to those with affinity-purified goat anti-human apoAI antibody (Harlan Sera-Lab).
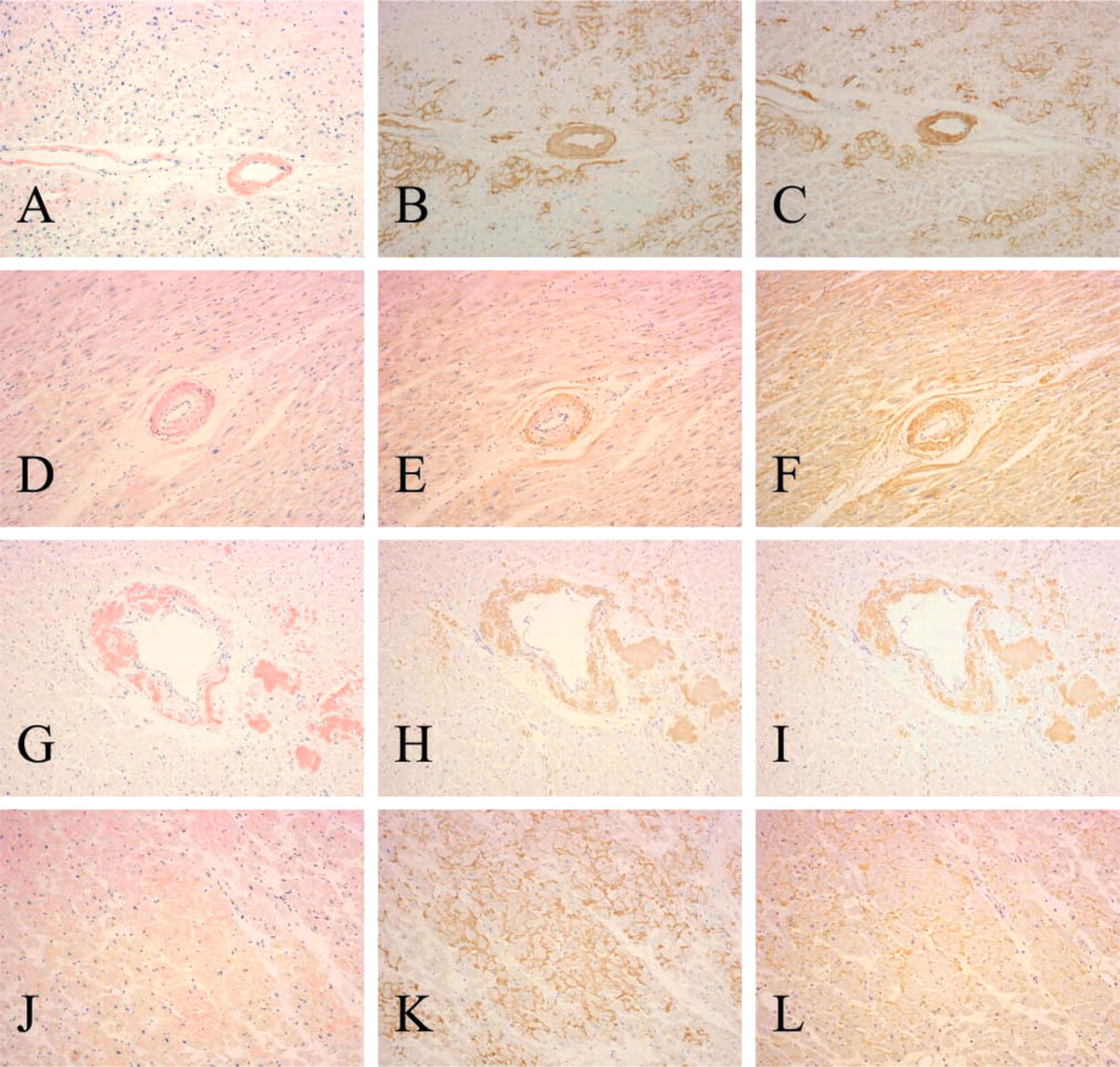
Case of AA amyloidosis
Immunoblotting of AL Amyloid
Anti-human apoAI antibody reacted with an approximate 25-kD band on Western blots of Aλ amyloid and purified human apoAI (Figure 2). Anti-λ (118-134) antibody reacted with several bands in Aλ amyloid but did not react with purified human apoAI. Furthermore, the anti-apoAI antibody did not recognize apoE, SAP, and SAA (data not shown).
Histological and Immunohistochemical Findings in Human ApoAI Transgenic Mice
After the injections of FCA and M. butyricum, amyloid deposits were observed in the perifollicular area of the spleen of all apoAI transgenic mice examined. Seventy-two hr after administration of AEF and AgNO3, all apoAI transgenic mice also had amyloid deposits in the perifollicular area of the spleen, but the amyloid deposits were smaller than those in mice treated with FCA and M. butyricum. These amyloid deposits reacted with anti-mouse AA antibody (Figures 3A and 3C). Anti-human apoAI antibody staining was not detected in amyloid deposits in apoAI transgenic mice after administration of AEF and AgNO3, but staining was present in the interstitium (Figure 3B). In contrast, anti-human apoAI immunoreactivity was present in both amyloid deposits and interstitium in apoAI transgenic mice after injection of FCA and M. butyricum (Figure 3D). The staining was irregular and weaker than that with anti-mouse AA antibody.
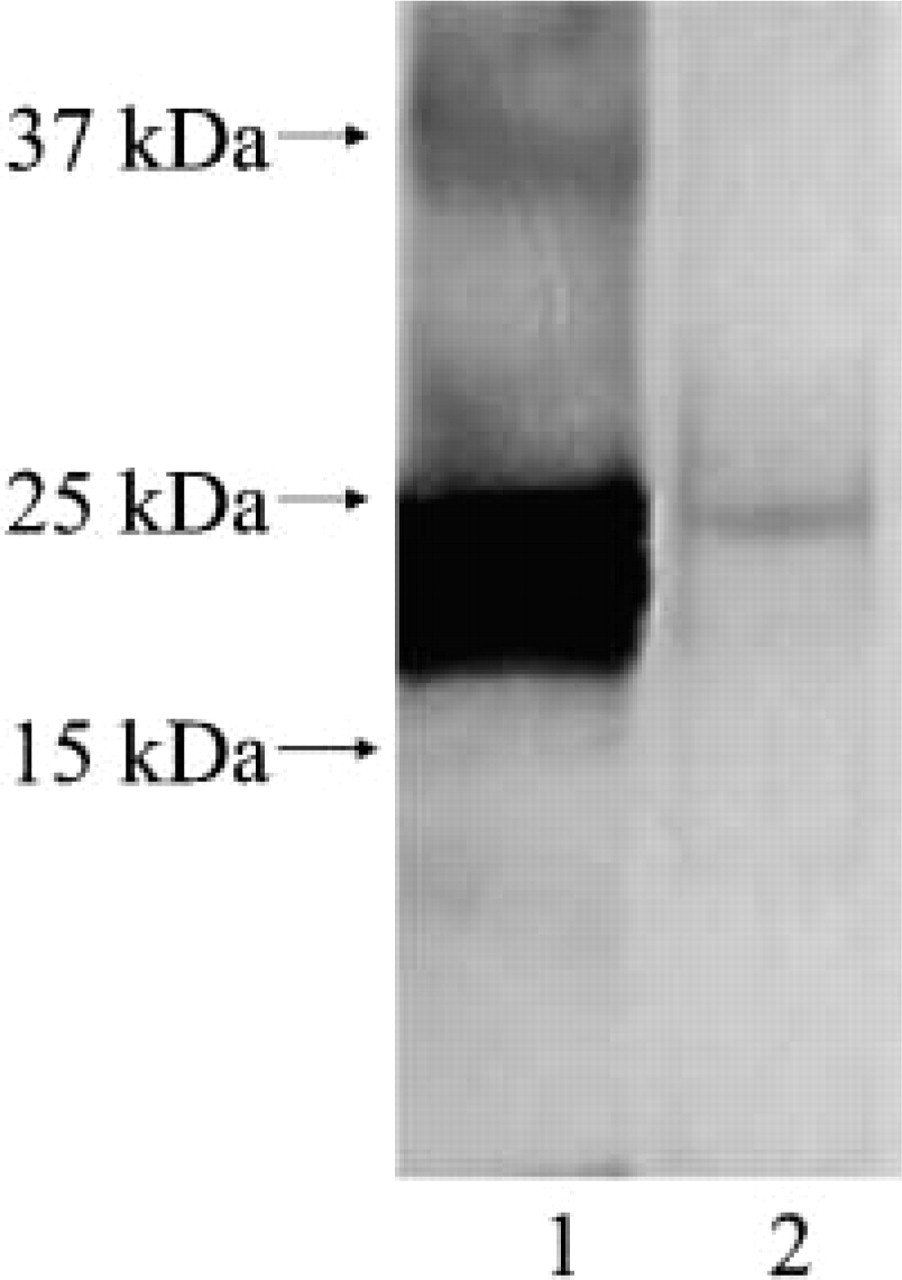
Immunoblotting of AL amyloid with anti-human apoAI antibody. Lane 1: purified human apoAI, lane 2: water-extracted Aλ amyloid. Anti-human apoAI antibody reacted with an approximate 25-kD band in Aλ amyloid and purified human apoAI.
Discussion
In the present study we observed colocalization of apoAI with amyloid deposits in various kinds of human systemic amyloidosis immunohistochemically and biochemically. We also detected colocalization of apoAI and murine AA amyloidosis in human apoAI transgenic mice. ApoAI is known to form amyloid fibrils as a major fibril protein and colocalizes with amyloid-β (Aβ) protein in Alzheimer's disease. However, this is the first report of the colocalization of apoAI with amyloid deposits in various kinds of human systemic amyloidosis. We should consider that apoAI may not always be a major component of amyloid fibrils even when amyloid deposits in systemic amyloidosis are positive for apoAI immunohistochemically. Serum proteins trapped in amyloid deposits are sometimes detected in the immunohistochemical analysis (Röcken et al. 1996; Strege et al. 1998). Therefore, it is possible that apoAI may only be trapped in amyloid deposits as one of serum protein. However, we found that apoAI was present in crude water-extracted Aλ amyloid, suggesting that apoAI was not merely trapped in amyloid deposits but rather was an insoluble component of amyloid deposits.
Involvement of apolipoproteins in amyloidosis has been reported to date. SAA (Benditt et al. 1979; Eriksen and Benditt 1980) and apoAII (Higuchi et al. 1986; Benson et al. 2001) are precursors of major amyloid fibril proteins in humans and mice. ApoE, which has not been known as a major amyloid fibril protein, is a minor constituent of various kinds of systemic amyloidosis (Wisniewski and Frangione 1992). ApoJ is present in the senile plaques of Alzheimer's disease (Harr et al. 1996) and may also be a minor constituent of amyloidosis.
ApoAI contains a high proportion of class A amphipathic α-helical domains similar to other apolipoproteins, including apoAII, apoJ, and apoE (Hatters and Howlett 2002). Both full-length apoAI and the N-terminal fragment of apoAI can form amyloid-like fibrils in vitro (Wisniewski et al. 1995a), and the C-terminal fragment of apoE can also form amyloid-like fibrils in vitro (Wisniewski et al. 1995b). Furthermore, apoAI and apoE can bind Aβ and promote Aβ fibrillogenesis in vitro (Castano et al. 1995; Wisniewski et al. 1995a, b). These similarities between apoAI and apoE suggest that the role of apoAI in various kinds of human systemic amyloidosis may be similar to that of apoE. Coincubation of AA with apoE in vitro resulted in a high degree of polymerization of AA peptides, and apoE can also form SDS-resistant AA and Aβ complexes (Castano et al. 1995). These findings suggest that apoE not only promotes amyloidogenesis but also resists proteolysis in vivo. Although similar findings have not been reported for apoAI, it may also promote amyloidogenesis or resist proteolysis in vivo. Chang et al. (2001) reported that apoE colocalizes with primary and secondary cutaneous amyloidosis but that apoAI is not associated with them. However, the immunostaining method and primary antibody used by Chang et al. (2001) differed from that used in the present study; therefore, their finding of no apoAI in primary and secondary cutaneous amyloidosis is not surprising.
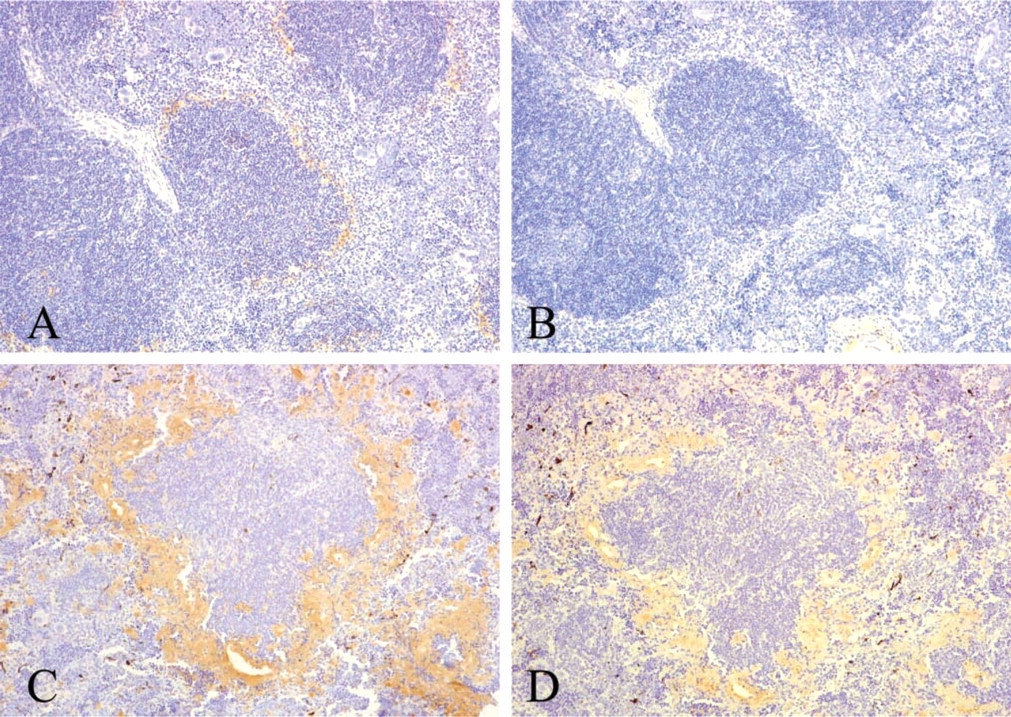
On the other hand, AA amyloidosis develops in both apoE-deficient mice (Hoshii et al. 1997) and apoAI-deficient mice (Elliott-Bryant and Cathcart 1997) after inflammatory stimulation. In our immunohistochemical study of AA amyloidosis in human apoAI transgenic mice, we did not observe apoAI immunoreactivity in amyloid deposits at the early stage of murine AA amyloidosis after administration of AEF and AgNO3. Furthermore, in our immunohistochemical analysis of various kinds of human systemic amyloidosis, the immunoreactive area and staining intensity of amyloid deposits were variable within each case. Although it is possible that formalin fixation of tissue specimens may reduce antigenicity and cause uneven staining, apoAI may not be distributed uniformly in amyloid deposits. These findings suggest that apoAI may not be essential for amyloidogenesis.
The exact role of apoAI in amyloidogenesis of various kinds of systemic amyloidosis is unclear. Further studies of the relation of apoAI and amyloid fibril proteins in human systemic amyloidosis are needed. A better understanding of these amyloid-associated proteins may contribute to development of strategies for the treatment of amyloidosis.