
Abstract
The localization of the type-2 angiotensin II receptor (AT2) in the adrenal glands of rats, guinea pigs, bovines, and humans was examined at the mRNA and protein levels. PCR products for AT2 were detected in the adrenal cortices and adrenal medullae of all the mammals examined with an RT-PCR technique. Three different anti-AT2 antibodies (Abs), whose specificity was confirmed in our hands, recognized a 50-kDa protein in the adrenal glands of the four mammals, and this recognition was abolished by the preabsorption of an Ab with an antigen. Immunoblotting and immunohistochemistry revealed that the 50-kDa protein was expressed consistently and variably in the adrenal cortices and medullae of various mammals, respectively. We conclude that the 50-kDa AT2 is consistently expressed in the adrenal cortex in a wide variety of mammals.
T
We have previously reported that exposure to a low concentration of ouabain in guinea pig AM cells results in an increase in the stored Ca2+ concentration, with the consequent facilitation of Ca2+ mobilization (Lin et al. 2005). Ouabain or ouabain-like substances have been found in AC cells of various mammals (Bagrov et al. 2009), such as bovines (Laredo et al. 1997), rats (Ludens et al. 1992), and humans (El-Masri et al. 2002). Therefore, if ouabain or a ouabain-like substance is released from AC cells in response to AT2 activation in such mammals (Schoner and Scheiner-Bobis 2007; Manunta et al. 2009; Nicholls et al. 2009), AT2 could be assumed to be physiologically important for catecholamine secretion from AM cells. The aim of the present experiment was to elucidate whether AT2 is expressed in AC cells in a wide range of mammals at the mRNA and protein levels.
Materials and Methods
All experimental procedures involving humans and animals were approved by the Institutional Ethics Committee and Animal Care and Use Committee of the University of Occupational and Environmental Health, respectively.
Immunoblot
The rats (n = 5) and guinea pigs (n = 5) were killed by cervical dislocation, and the adrenal glands were excised and immediately put into ice-cold Ca2+ -deficient balanced salt solution, which contained 137 mM NaCl, 5.4 mM KCl, 0.5 mM MgCl2, 0.53 mM NaHPO4, 5 mM
Immunohistochemistry
The rats (n = 2) and guinea pigs (n = 2) were anesthetized with sodium pentobarbital (50 mg/kg−1 intraperitoneally) and perfused through the ascending aorta with 30 ml of saline, then 250 ml of Zamboni's fixative. The adrenal glands were fixed in the fixative overnight at 4C. After fixation, they were rinsed in PBS, and then the medium was successively exchanged with 10%, 20%, and 30% sucrose-containing PBS. Thin sections (10 mm thick) were obtained with a cryostat (OTF5000 CRYOSTAT; Bright, Huntingdon, UK) and mounted on glass slides (MAS-coated Superfrost; Matsunami, Kishiwada, Japan). After treatment with 0.2% casein for 60 min to reduce the nonspecific binding, the sections were incubated overnight with the SC-made anti-AT2 Ab diluted 1:50, followed by treatment with anti-rabbit IgG Abs conjugated with Alexa 488 or Alexa 546 (Molecular Probes; Eugene, OR). The fluorescence was observed with a confocal laser scanning microscope (LSM5 Pascal; Carl Zeiss, Oberkochen, Germany). For Alexa 488, a 488-nm laser was used, and emission of 510–560 nm was observed (FITC fluorescence); for Alexa 546, a 543-nm laser was used, and emission >570 nm was observed (rhodamine fluorescence).
The adrenal glands of bovines (n = 3) were fixed in the fixative overnight at 4C. After fixation and rinsing in PBS, they were dehydrated through a graded ethanol series and embedded in paraffin (Histosec; Merck, Darmstadt, Germany). Thin sections (5 μm thick) were obtained with a microtome, mounted on glass slides, dried overnight, and deparaffinized. Next, the sections were rinsed for 10 min in PBS. Endogenous peroxidase activity was inhibited by pretreatment with 0.1% hydrogen peroxide in methanol for 20 min. After treatment with 0.2% casein for 60 min to reduce the nonspecific binding, the sections were incubated overnight with the AD-made anti-AT2 Ab. After rinsing in PBS, the immunoreaction was examined using the indirect immunoperoxidase method (Histofine Simple Stain Max-PO; Nichirei, Tokyo, Japan). The peroxidase complex was visualized by treatment with a freshly prepared solution of diaminobenzidine tetrahydrochloride (DAB) (DAB Substrate Kit; Nichirei), and the diaminobenzidine reaction was enhanced by the addition of nickel ammonium sulfate.
Immunocytochemistry
AM cells were dissociated from adrenal medullae of guinea pigs (n = 2) with collagenase treatment, as described previously (Inoue et al. 2008). For indirect immunofluorescence studies, cells were treated overnight with the SC anti-AT2 Ab diluted 1:50 to 1:200. After incubation, the cells were washed three times in PBS and then treated with an anti-rabbit IgG Ab conjugated with Alexa 488 (Molecular Probes). Fluorescence was observed with an LSM5 Pascal microscope.
Cell Culture and Transfection
Human embryonic kidney (HEK) 293T cells were cultured in DMEM (Invitrogen; Carlsbad, CA) supplemented with 10% fetal bovine serum (Nichirei). Lipofectamine 2000 Reagent (Invitrogen) was used to transfect HEK293T cells with the expression vector for hemagglutinin (HA)-tagged human AT2 (Missouri S and T cDNA Resource Center; Rolla, MO) according to the manufacturer's instructions. The transfected cells were placed onto glass coverslips coated with collagen type I (BD Biosciences; San Jose, CA), and then cultured for 24 hr. The cells were fixed with 4% parafor-maldehyde in PBS for 30 min at room temperature. After washing three times with PBS, the cells were incubated in PBS containing 0.1% Triton X-100 for 30 min and then with PBS containing 1% FBS for 1 hr at room temperature. The cells were treated with primary Abs and then with secondary Abs. The coverslips were mounted in 50% glycerol containing 1 mg/ml 1,4-diaminobenzene, and immunostaining was observed with an LSM5 Pascal microscope.
RT-PCR
The Micro-fast Track Kit (Invitrogen) was used according to the manufacturer's instructions to isolate poly(A)+ RNA from the brain, adrenal medulla, and/or adrenal cortex of rats (n = 5), bovines (n = 1), and humans (n = 2). To digest the genomic DNA, mRNA prepared from rat and human tissues was treated with DNase I (Invitrogen). Oligo dT primer was utilized for the reverse transcriptase (RT) reaction to obtain cDNAs. PCR reactions were carried out with 1.25 μl of DNA template, 4 pmol of primer, 2 mM of dNTPs, 0.5 U KOD FX DNA polymerase (Toyobo; Osaka, Japan), and PCR buffers supplied with the kit, in a final volume of 25 μl. Table 1 lists the primers for the PCR. The PCR protocol used started with an initial 3-min denaturation step at 94C, followed by 30–40 cycles of the profile consisting of 30 sec of denaturation at 94C, 30 sec of annealing at 54C to 60C, and 30 sec of extension at 72C. To obtain the maximum fidelity, a hot-start procedure was used. In each PCR reaction, a 198-bp PCR product of β-actin mRNA was amplified simultaneously or separately and used as an internal standard. The PCR products were separated by 1.5% agarose gel electrophoresis, and stained with ethidium bromide.
Results RT-PCR
The expression of AT2 at the mRNA level was examined with the RT-PCR technique. Primer sequences for PCR analysis were selected so that an intron was present between the forward and backward primers. Therefore, PCR products originating from the genome were clearly differentiated from PCR products from cDNAs (Figure 1A). PCR products of 285 bp, the size estimated for AT2 mRNA, were detected in cDNA samples of rat brain, adrenal cortex, and adrenal medulla. PCR products of the same size were also recognized in samples of both bovine adrenal cortex and adrenal medulla (Figure 1A). Next, the same set of primers was used to examine whether mRNA for AT2 was expressed in human adrenal cortex and adrenal medulla. PCR products of 285 bp were detected in cDNA samples of human adrenal medulla, but not adrenal cortex, whereas β-actin amplicons were found in both samples (not shown). This failure of detection in the adrenal cortex was possibly due to deterioration of the human adrenal cortex, which was excised during surgery. To reduce the effect of deterioration, backward and forward primer sequences were selected to be near the beginning of the coding region in mRNA for AT2 without insertion of an intron. Therefore, non-RT samples were simultaneously subjected to PCR analysis to exclude the possible contamination of genomic DNA. Figure 1B shows that AT2 amplicons were detected in both adrenal cortex and adrenal medulla cDNAs, but not in non-RT samples, indicating that PCR products for AT2 originate from cDNA, but not genomic DNA. The results indicate that mRNAs for AT2 are produced in both the adrenal cortex and the adrenal medulla of rats, bovines, and humans.
Primer sequences used for PCR of AT2 receptor and β-actin

AT2, type-2 angiotensin II.
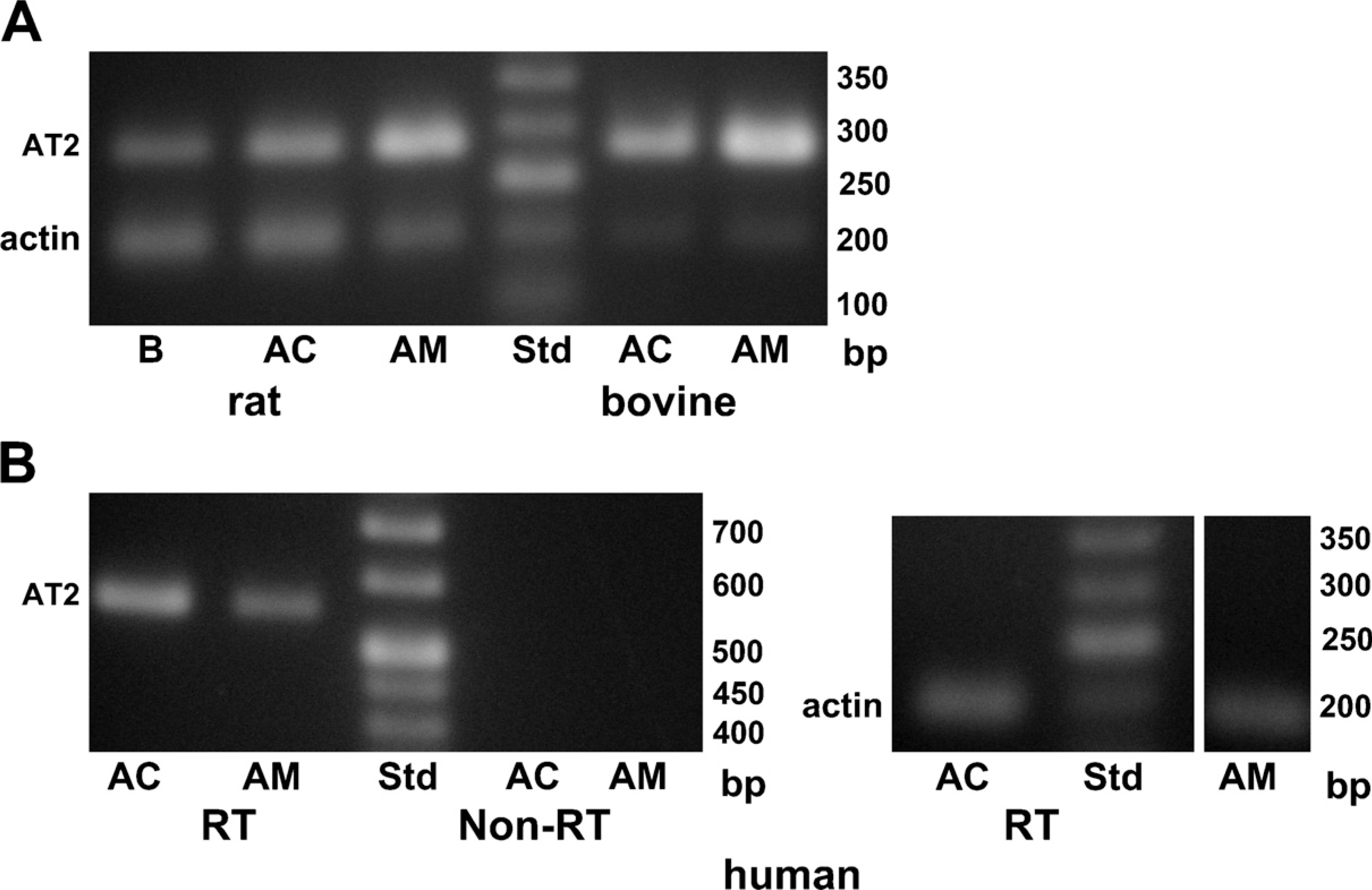
PCR analysis for expression of the type-2 angiotensin II receptor (AT2).
Immunoblotting
The molecular mass of AT2 identified immunologically or autoradiographically varied among the organs and the species (Ouali et al. 1993; Servant et al. 1996; Nora et al. 1998; Wang et al. 1998; Frei et al. 2001; Carneiro-Ramos et al. 2007). Thus, three anti-AT2 Abs that were raised against different antigens were used. Figure 2A shows immunoblotting with an AL-made anti-AT2 Ab. The Ab clearly recognized a 50-kDa band in the rat adrenal cortex but not in the adrenal medulla, and this detection was markedly suppressed by preabsorption of the Ab with the antigen, suggesting that the Ab is specific for AT2. The AL Ab also detected the 50-kDa band in guinea pig adrenal medulla as well as adrenal cortex (Figure 2B). Figure 2C summarizes the expression levels of AT2 in rat and guinea pig adrenal glands. The amount of protein expressed in the guinea pig adrenal medulla was about half the amount in the adrenal cortex, whereas the amount of AT2 in the rat adrenal medulla was one-tenth that in the adrenal cortex. Immunoblotting of rat and guinea pig adrenal glands was also performed with an SC-made anti-AT2 Ab with similar results (not shown). To explore the expression of AT2 in bovine and human adrenal glands, SC- and AD-made anti-AT2 Abs were used. As shown in Figure 3A, both Abs recognized 50-kDa bands in bovine adrenal cortex and medulla. Similarly, the 50-kDa band was recognized in human adrenal glands. However, the expression levels of the protein in humans differed from those in the other three mammals; the amount of 50-kDa protein in the human adrenal medulla was about twice that in the adrenal cortex (Figure 3B).
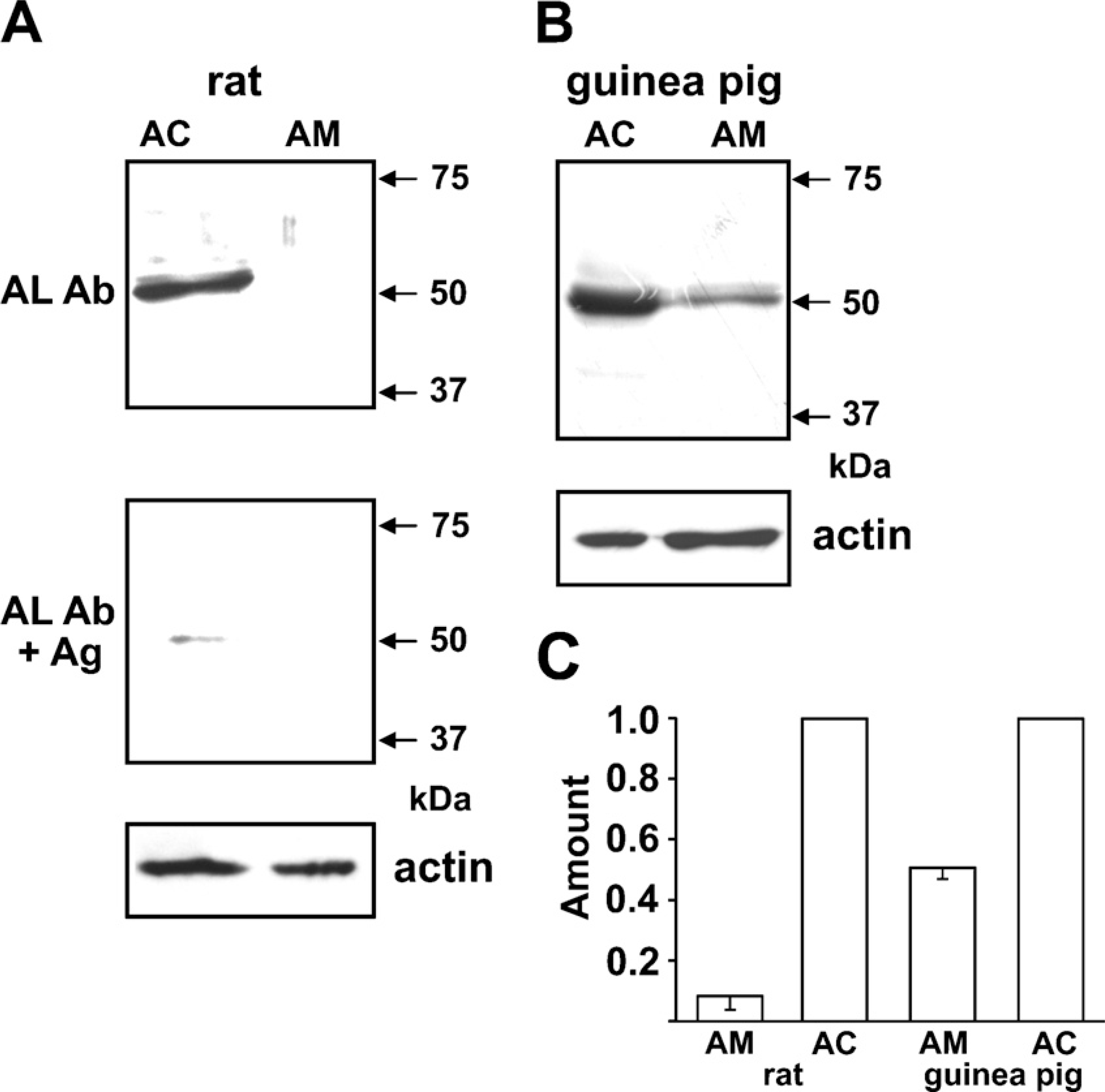
Immunoblot of rat and guinea pig adrenal glands for AT2.
Immunohistochemistry
We first examined the specificity of the AD- and SC-made Abs in immunohistochemistry. Figure 4A shows that the AD-made Ab selectively stained the HEK294T cells transfected with an AT2 vector encoding the human AT2 and HA tag, indicating that the Ab is specific for AT2 in staining. Similar results were obtained with the SC-made Ab (not shown). The AD-made Ab produced significant staining in the bovine adrenal cortex (n = 6), compared with that in the adrenal medulla, whereas an anti-dopamine-β-hydroxylase Ab stained the adrenal medulla and not the adrenal cortex (Figure 4B). Next, the rat adrenal gland was immunohistochemically examined with the SC anti-AT2 Ab. Consistent with the expression levels obtained with immunoblotting, the intensity of immunofluorescence in the adrenal cortex was significantly higher than that in the adrenal medulla (n = 5). On the other hand, fluorescence was not observed in any part of the section not treated with the Ab (Figure 4C), suggesting that fluorescence was due to immunoreaction with the anti-AT2 Ab. The section of guinea pig adrenal gland was also examined with the SC-made AT2 Ab. As shown in Figure 5A, the adrenal medulla and cortex were rather homogeneously stained (n = 6), whereas the section treated without the Ab did not show noticeable fluorescence. Finally, the intracellular localization of AT2 was examined in isolated guinea pig AM and AC cells. In both cells (AM, n = 11; AC, n = 8), AT2-like immunoreactivity was mainly located in the plasma membrane or in its vicinity (Figure 5B).
Discussion
Whether ouabain or a ouabain-like substance is secreted from AC cells remains controversial (Murrell et al. 2005; Schoner and Scheiner-Bobis 2007; Manunta et al. 2009; Nicholls et al. 2009). Ouabain or a ouabain-like substance, however, has consistently been found in the adrenal cortex of rats (Ludens et al. 1992), bovines (Laredo et al. 1997), and humans (El-Masri et al. 2002). If endogenous ouabain is released from AC cells following stimulation of AT2, as has been suggested in bovine AC cells (Laredo et al. 1997), the secreted ouabain may be expected to enhance catecholamine secretion in AM cells. In fact, application of a low concentration of ouabain resulted in an increase in Ca2+ mobilization–induced secretion in guinea pig AM cells (Lin et al. 2005). If this hypothesis is correct, ouabain would be expected to ameliorate, through its direct and catecholamine-mediated effects on the heart, a decrease in renal blood flow, which may be a cause for an increase in blood AGII in the classical renin-angiotensin system. From this point of view, it would be important to elucidate whether AT2 is consistently expressed in mammalian AC cells or not. In the present experiment, this issue was examined at the mRNA and protein levels. The RT-PCR analysis revealed that the AT2 receptor was expressed at the mRNA level in adrenal cortices and medullae of humans, bovines, and rats. On the other hand, PCR products were not detected in any cDNA samples of guinea pig brain, adrenal cortex, or adrenal medulla with a set of primers that was successful for PCR of rat samples. This failure was probably due to a slight difference in gene structure. Expression at the protein level was explored with three distinct anti-AT2 Abs, which had been raised against different epitopes. All three Abs clearly recognized a 50-kDa protein in the adrenal cortex in rats, guinea pigs, bovines, and/or humans, and recognition of the protein by the AL-made Ab was suppressed by preabsorption of the Ab with the antigen. On the other hand, the AT2 in the adrenal medulla was detected at low levels in bovines and guinea pigs and prominently in humans, but it was scarcely recognized in rats. These differences in expression of the 50-kDa protein between the adrenal cortices and medullae of bovines, rats, and guinea pigs were also observed in immunohistochemical studies with the SC- and AD-made Abs. On the basis of the immuno-blotting and immunohistochemical findings, we conclude that the 50-kDa protein represents AT2 in the adrenal gland and is expressed consistently and variably in the adrenal cortex and medulla, respectively. The finding that AT2 was expressed predominantly in the rat adrenal cortex and scarcely in the adrenal medulla conflicts with previous results showing that AT2-like immunoreactive substances were exclusively present in the rat adrenal medulla and that mRNA for AT2 was detected in the rat adrenal medulla and zona glomerulosa, and not in the zona reticularis or zona fasciculata, with the in situ hybridization technique. The anti-AT2 Ab used in the previous immunohistochemical analyses recognized a protein of ×70 kDa, which differed from the 50-kDa protein in the present experiment. Therefore, the immunoreactivity obtained with the previous Ab might have reflected a protein unrelated to AT2. Furthermore, the expression of mRNA for AT2 may not necessarily indicate that of AT2 at the protein level. In our previous experiments, TASK3 channels in the rat adrenal medulla were detected at the mRNA level but not at the protein level (Inoue et al. 2008). A similar discrepancy of expression between the mRNA and protein levels has been noted with membrane proteins, such as other K+ channels (Barry et al. 1995).
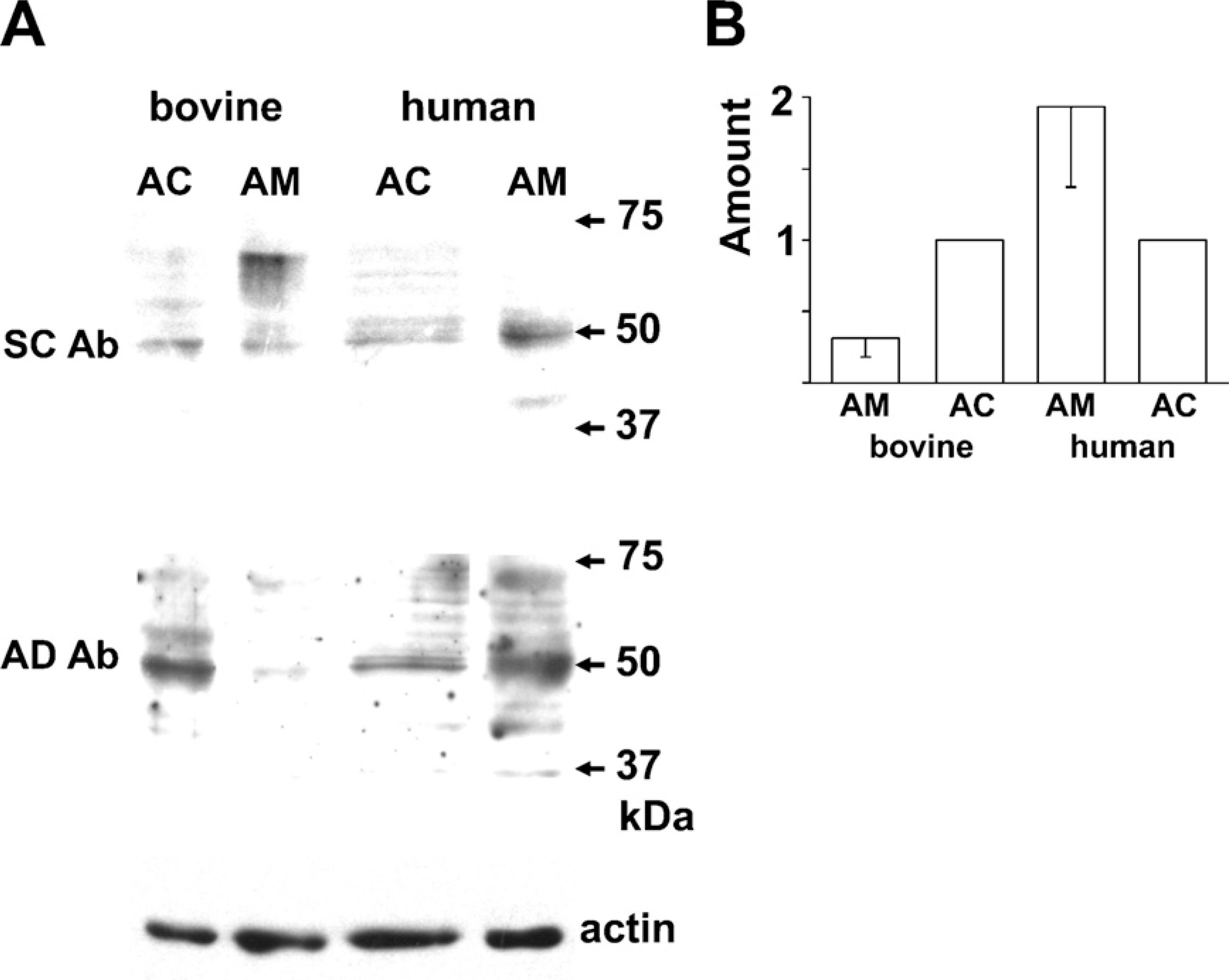
Immunoblot of bovine and human adrenal glands for AT2.
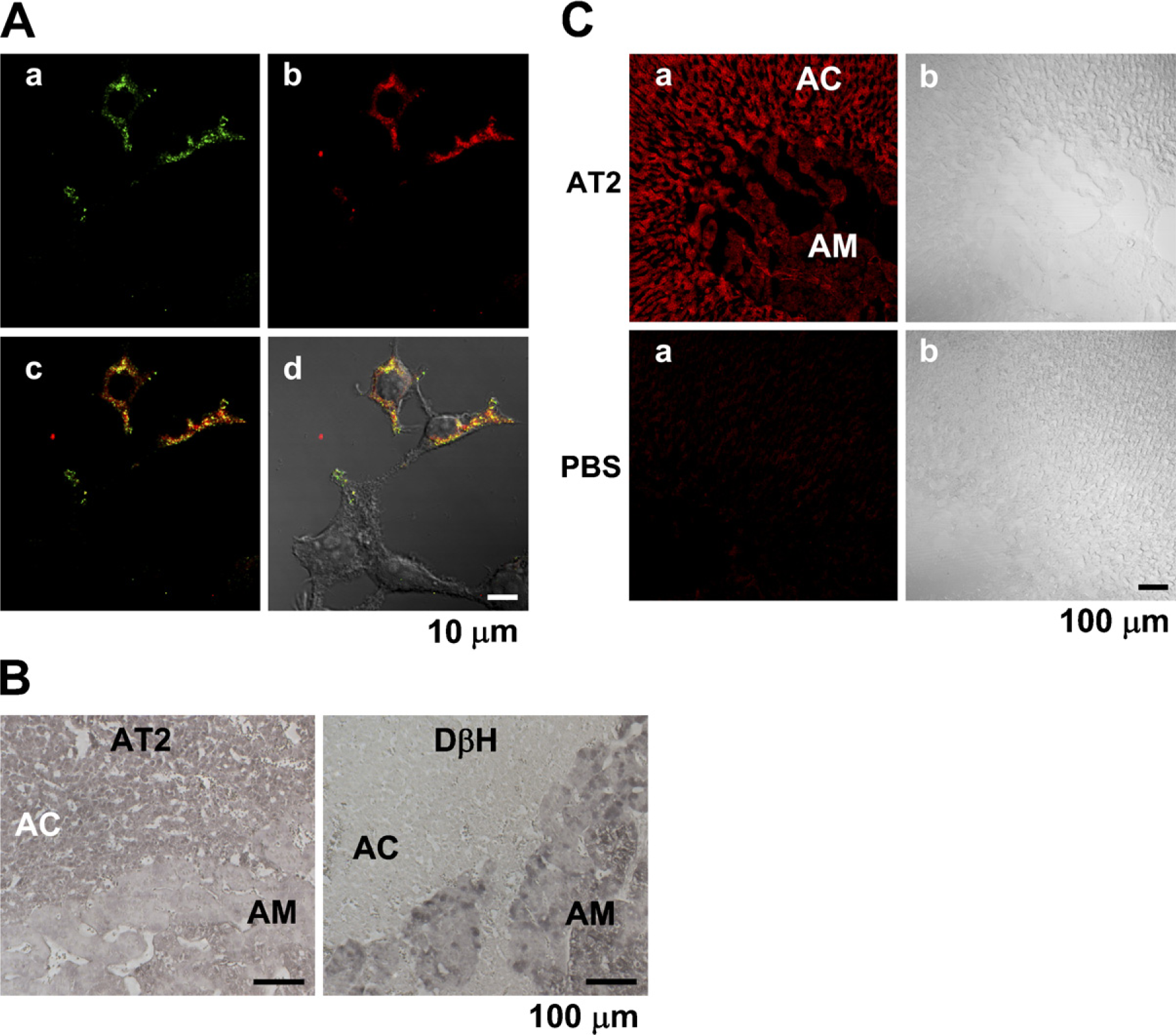
Immunohistochemistry of bovine and rat adrenal glands for AT2.
Belloni et al. (1998) noted that [125I]AGII binding in the rat adrenal gland occurred in the medulla and zona glomerulosa. The binding in the former and latter were markedly suppressed by CGP42112, an AT2 agonist, and DuP753, an AT1 antagonist, respectively, and AGII-induced catecholamine secretion was conspicuously inhibited by PD123319, an AT2 antagonist, but not by DuP753. These results seem to suggest that AT2 is selectively expressed in the rat adrenal medulla and is involved in catecholamine secretion in response to AGII. This notion, however, is not compatible with the electrophysiological findings that in bovine AM cells, AGII receptors involved in catecholamine secretion showed rapid desensitization (Teschemacher and Seward 2000), which is characteristic of AT1 and not of AT2 (De Gasparo et al. 2000; Landon and Inagami 2005; Mogi et al. 2007), that AGII produced a Ca2+ mobilization with the consequent release of catecholamine through a phospholipase C pathway (Stauderman and Pruss 1990; Cheek et al. 1993; Teschemacher and Seward 2000), and most importantly that AGII-induced secretion was completely suppressed by DuP753, but not by PD123319 (Teschemacher and Seward 2000). This difference would warrant a more detailed study to identify the subtype of AGII receptor expressed in the rat adrenal medulla at the protein level.
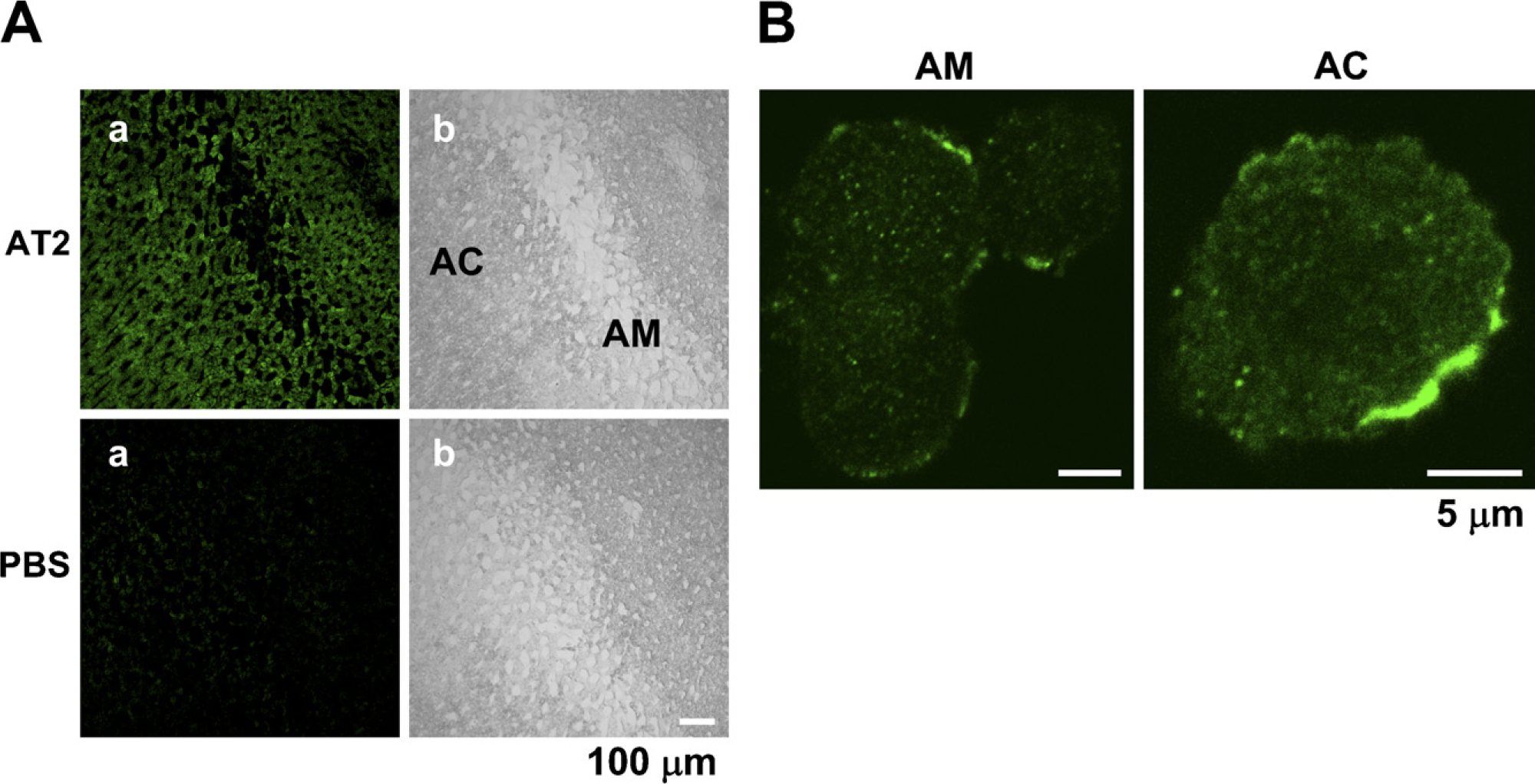
Immunostaining of guinea pig adrenal gland for AT2.
In summary, three different anti-AT2 Abs, whose specificity was confirmed in our hands, recognized a 50-kDa protein in adrenal glands of humans, rats, guinea pigs, and bovines, and the recognition of the protein was abolished by the preabsorption of an Ab with an antigen. Immunoblotting and immunohistochemistry revealed that the 50-kDa protein was expressed consistently and variably in adrenal cortices and medullae of various mammals, respectively. We conclude that AT2 receptors of 50 kDa are consistently expressed in the adrenal cortex in a wide variety of mammals. This consistent expression of AT2 in the adrenal cortex may suggest that AT2 receptors play an essential role in the functions of the adrenal cortex. One such function might be the regulation of the secretion of ouabain or a ouabain-like substance.
Footnotes
Acknowledgements
This work was supported in part by Grant-in-Aid for Scientific Research on Priority Areas (21026029 to MI) and Grant-in-Aid for Scientific Research (C) (21500360 to MI).
We are grateful to Y. Doi for guidance on the histochemical studies and T. Hatama for technical assistance.