
Abstract
Apoptosis competence is central to the prevention of cancer. Frequency of apoptotic cells, after a sample of colonic tissue is stressed, can be used to gauge apoptosis competence and, thus, possible susceptibility to colon cancer. The gold standard for assessment of apoptosis is morphological evaluation, but this requires an experienced microscopist. Easier-to-use immunohistochemical markers of apoptosis, applicable in archived paraffin-embedded tissue, have been commercially developed. Potentially useful apoptosis markers include cleaved cytokeratin-18 (c-CK18), cleaved caspase-3 (c-cas-3), cleaved lamin A (c-lam-A), phosphorylated histone H2AX (γH2AX), cleaved poly(ADP ribose) polymerase (c-PARP), and translocation of apoptosis-inducing factor (AIF). When tissue samples from freshly resected colon segments were challenged ex vivo with the bile acid deoxycholate, ∼50% of goblet cells became apoptotic by morphologic criteria. This high level of morphologic apoptosis allowed quantitative comparison with the usefulness and specificity of immunohistochemical markers of apoptosis. The antibody to c-CK18 was almost as useful and about as specific as morphology for identifying apoptotic colonic epithelial cells. Antibodies to c-cas-3, c-lam-A, and γH2AX, though specific for apoptotic cells, were less useful. The antibody to c-PARP, though specific for apoptotic cells, had low usefulness, and the antibody to AIF was relatively nonspecific, under our conditions.
A
A number of pathways appear to be important in stress-induced apoptosis, some of which depend on cell type as well as on type of apoptosis-inducing agent (Inagaki-Ohara et al. 2002). A schematic of events accompanying some known stress-induced apoptosis pathways is shown in Figure 1. One set of early steps in apoptosis induction involves alteration of the mitochondrial membrane with release of cytochrome c, apoptosis-inducing factor (AIF), and endonuclease G (Antonsson 2004). Release of cytochrome c activates a caspase-dependent pathway involving cytochrome c assembly with procaspase-9 and Apaf-1 in close association with cytokeratin 18 (CK18) (Dinsdale et al. 2004). This leads to formation of the apoptosome on CK18. Next, procaspase-9 is cleaved and activated, forming c-cas-9, which then cleaves and activates both procaspase-3, producing c-cas-3, and procaspase-7, producing c-cas-7 (Slee et al. 1999). This is followed by c-cas-3-catalyzed cleavage of procaspase-6 (and additional cleavage of procaspase-9) (Slee et al. 1999; Ruchaud et al. 2002). Then, c-cas-3, c-cas-6, and c-cas-7 (c-cas-7 action not shown) cleave CK18, forming c-CK18 (Caulin et al. 1997). C-cas-3 and c-cas-7 both cleave poly(ADP-ribose) polymerase (PARP), forming c-PARP (Soldani and Scovassi 2002). C-cas-6, subsequent to its cleavage by c-cas-3 (Slee et al. 1999), cleaves lamin A in the nucleus, forming c-lam-A (Ruchaud et al. 2002).
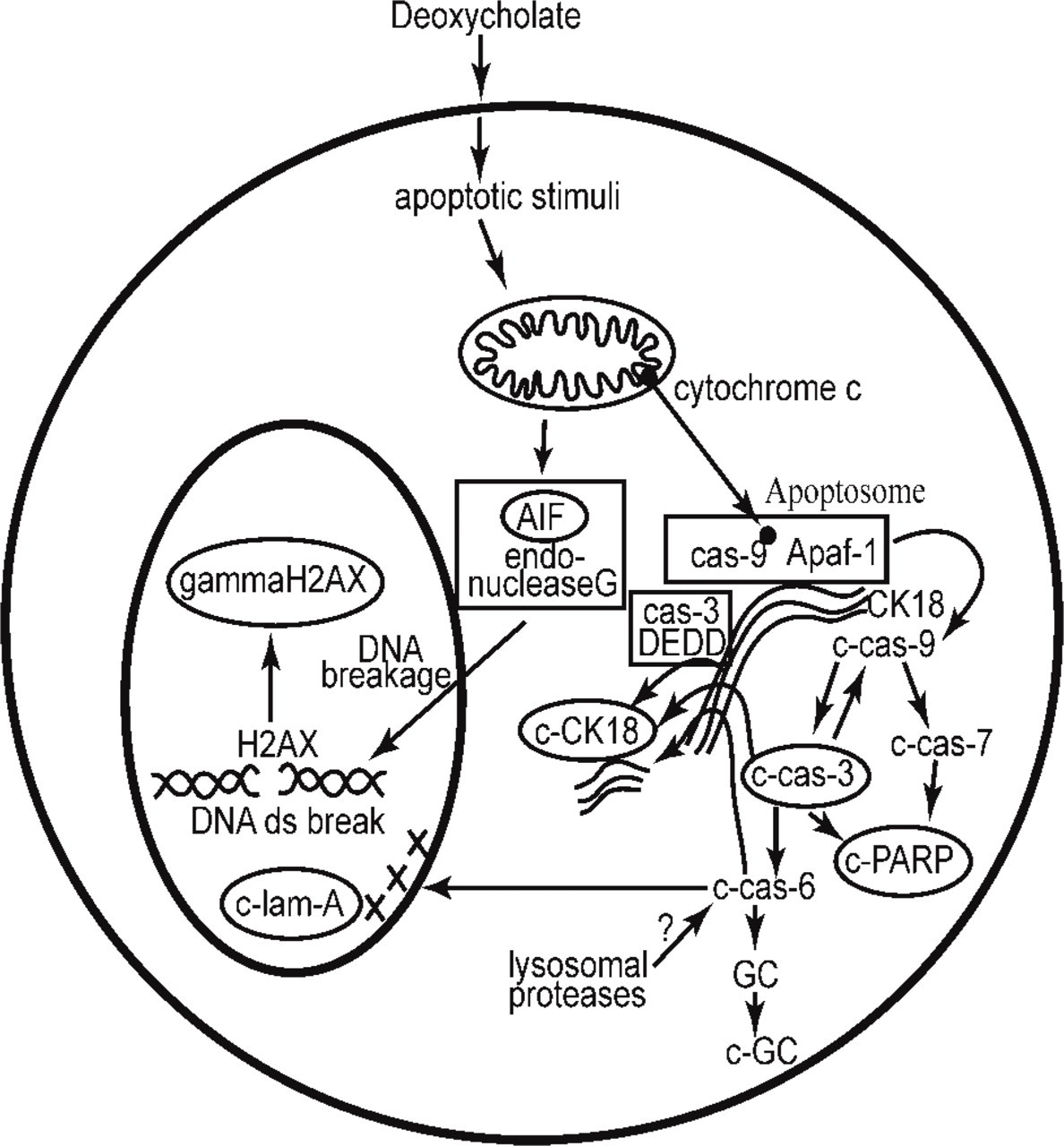
A schematic diagram of some events accompanying stress-induced apoptosis. The immunohistochemical markers evaluated in this study are shown within ovals.
Another pathway, which is caspase independent, is initiated by AIF and endonuclease G translocation to the nucleus with subsequent DNA double-strand breakage (Antonsson 2004), followed by formation of phosphorylated H2AX (γH2AX) from H2AX (Rogakou et al. 2000). Yet another pathway is implemented through the protein DEDD (death effector domain containing DNA-binding protein). DEDD, in its cytoplasmic location, mediates recruitment of procaspase-3 (cas-3) to CK18 during apoptosis, leading to cleavage of CK18 (Lee et al. 2002). In addition to the pathways illustrated in Figure 1, there may be a further pathway for cleavage of CK18. C-cas-6, known to be highly effective in cleavage of CK18 (Caulin et al. 1997), may be activated by steps outside the apoptosome-initiated pathway [e.g., lysosomal protease activity (Payne et al. 2003)]. In addition, we have shown that the cleavage of guanylate cyclase, a step in deoxycholate-induced apoptosis, was dependent on caspase-6 but not on caspase-3, −7, or −9 (Payne et al. 2003).
The modified proteins selected as potential markers for apoptosis in our colonic mucosa samples were chosen from the schematic of known steps of apoptosis outlined in Figure 1 and include c-CK18, c-cas-3, c-lam-A, γH2AX, c-PARP, and AIF. This set of apoptosis-related proteins represents events taking place in the nucleus (γH2AX, c-PARP, c-lam-A, AIF, and c-cas-3) and cytoplasm (c-cas-3, c-CK18) and represents distinct events in the apoptotic process. It should be noted, however, that cytokeratin 18 is present in the simple (glandular) epithelium of the colonic mucosa but present only at low levels or not present at all in many other cell types (Bosch et al. 1988; Chu and Weiss 2002). Antibodies to the set of six modified proteins listed above are commercially available, known to be effective in archived paraffin-embedded tissue, and thought to recognize neo-epitopes (or altered location) frequently present within, and specific to, apoptotic cells. We assessed the usefulness and specificity of these antibodies in comparison to morphological measurements of apoptotic cells using multiple colonic tissue samples from nine individuals, treated ex vivo with deoxycholate.
Materials and Methods
Multiple samples were obtained from each of the surgical resections of colons of seven patients. From these seven patients, two small samples of colonic mucosa were taken for assessing apoptosis by morphology, with the tissue to undergo 3-hr incubation in “stress media” [tissue culture media containing 1.0 mM deoxycholate (NaDOC)] as described below. From two of these seven patients, tissue samples were also fixed immediately for assessing background level of apoptosis by morphology, before any stress was encountered. Further, for two patients with colons free of neoplasia (who were in addition to the seven who underwent colonic resection), four tissue samples were obtained by biopsies from each patient. Two of the tissue samples from each of these patients were incubated in media alone for 3 hr (to determine the stress of media incubation) and two were incubated in media plus 1.0 mM NaDOC. For the tissues from these two patients, apoptosis was evaluated by strictly morphological criteria as described below.
For the seven patients undergoing colonic resection, one to four larger samples were taken for assessing apoptosis by immunohistochemistry in ethanol-fixed tissue. These mucosal samples were either immediately fixed in 70% alcohol and embedded in paraffin, incubated in tissue culture media alone, or incubated in tissue culture medium with 1 mM Na-DOC under standard apoptosis-inducing conditions (3 hr at 37C, 5% CO2) (Bernstein et al. 1999,2002) and then fixed in 70% alcohol and embedded in paraffin. There were one to four samples each for assessment at time zero (no stress), for incubation for 3 hr in tissue culture media, and for incubation for 3 hr in media plus 1.0 mM NaDOC. In addition, we wished to compare results in formalin-fixed tissue with results in ethanol-fixed tissue. Thus, one to four samples were taken from the colon resections from two patients, incubated in media plus 1.0 mM NaDOC for 3 hr, fixed in formalin, embedded in paraffin, and then evaluated for apoptosis by immunohistochemistry with cleaved cytokeratin 18 and cleaved caspase 3.
All patients gave written informed consent under protocols approved by the Human Subjects Committee (the Institutional Review Board) of the University of Arizona. Three of the patients who had a colon resection had a colonic adenocarcinoma, three had diverticulitis, and one had no evidence of colonic disease. The patient with the normal colon, but a resection, had a recurrent liposarcoma external to, but not involving, the colonic wall. The colon was resected as part of the surgical procedure to remove the liposarcoma (and surrounding tissues).
We previously determined that stressing normal colonic tissue ex vivo with 1 mM NaDOC under standard conditions generates an average of 57.6% apoptosis (±13.0%, SD) among goblet cells (Bernstein et al. 1999). This high level of apoptosis was helpful for the quantitative evaluation of the immunohistochemical apoptotic markers tested here.
Expression of antibodies to c-CK18, c-cas-3, c-lam-A, γH2AX, c-PARP, and AIF were evaluated using a modified immunohistochemical method described previously (Payne et al. 1998). Briefly, the paraffin-embedded tissues were cut into 4-μm sections, deparaffinized, and rehydrated. Endogenous peroxidase activity was blocked by incubation in 1% hydrogen peroxide in methanol for 30 min, and the sections were then rinsed with phosphate-buffered saline (PBS). To prevent nonspecific binding, the slides were incubated with 1.5% appropriate normal serum (Vector Laboratories; Burlingame, CA). The primary antibodies used were mouse monoclonal antibody M30 specific for c-CK18 (Roche Molecular Biochemicals; Indianapolis, IN), rabbit polyclonal antibody for c-cas-3 (Cell Signaling Technology, Inc.; Beverly, MA), rabbit polyclonal antibody for c-lamin-A (Cell Signaling Technology, Inc.), rabbit polyclonal antibody for γH2AX (Upstate Biotechnology, Inc.; Lake Placid, NY), mouse monoclonal antibody for c-PARP (Cell Signaling Technology, Inc.), and rabbit polyclonal antibody for AIF (Santa Cruz Biotechnology, Inc.; Santa Cruz, CA). After rinsing with PBS, biotinylated rabbit anti-mouse IgG F(ab')2 (DAKO; Carpinteria, CA) and goat anti-rabbit IgG (Vector Laboratories) were used for the mouse monoclonal and rabbit polyclonal primary antibodies, respectively. Immunocontrol slides were prepared by replacing the primary antibody with mouse IgG2b (for c-CK18 and c-PARP) or rabbit IgG (for c-cas-3, c-lam-A, γH2AX, and AIF) at the same protein concentration as the primary antibody. After rinsing in PBS, the Vectastain Elite ABC kit (Avidin, Biotin Enzyme Complex; Vector Laboratories) was used according to the manufacturer's instructions. Color was developed by applying diaminobenzidine tetrahydrochloride. (Sigma; St Louis, MO) supplemented with 0.04% hydrogen peroxide. Sections were counterstained with hematoxylin (Sigma) and mounted using Cytoseal. Observation of immunohistochemical staining of crypts was performed by one individual.
Apoptosis was determined by morphology in 1-μm epoxy resin sections as described previously (Bernstein et al. 1999, 2002). Briefly, deoxycholate-stressed tissue was removed from its incubation medium and immersed in 2 ml of cold, half-strength Karnovsky's fixative (pH 7.2) overnight at 4C and then transferred to 0.1 M phosphate buffer (pH 7.2). The tissue was then post-osmicated, dehydrated in a graded series of ethanols, and embedded in Spurr's epoxy resin. Epoxy sections (1 μm) were prepared using glass knives, and the sections were heat-attached to slides for 5 min on a hot plate maintained at 80C. The sections were then stained with methylene blue-azure II-basic fuschin (polychrome stain) and rinsed with distilled water. The proportion of apoptotic goblet cells (ratio of darkly stained apoptotic cells to lightly stained non-apoptotic cells) was ascertained by light microscopy under a x100 oil-immersion lens. Only nuclei that clearly belonged with the mucin droplet-containing cytoplasm of the goblet cells were scored when obtaining an apoptotic index (AI). A mean AI was obtained as the ratio of apoptotic goblet cells in a tissue sample to the total number of clearly identified goblet cells in that tissue. At least 100 goblet cells obtained from more than 10 different crypts were scored. Goblet cells throughout entire crypts were evaluated in obtaining the AI. Inter-observer variability was previously determined for this method of evaluating AI, and the correlation between observers was found to be 0.89 (p<0.001), with 1.0 representing complete agreement (Bernstein et al. 1999).
In addition, a rapid crypt apoptotic index (CAI) was measured, using the same polychrome-stained 1-μm sections as used for AI. Here, only the basal one-fourth of well-oriented crypts (with the crypt lumen visible) was examined. The strictly morphologic CAI was measured as the percent of these crypt basal regions with at least one cell recognized as apoptotic by morphologic features, whether a goblet cell or non-goblet cell. An immunohistochemical CAI was also measured on the 4-μm immunohistochemically stained, paraffin-embedded tissue sections. In this case, CAI was the percent of crypt basal regions with at least one immunohistochemically stained epithelial cell. CAI was determined by conjoint observation by two individuals.
Results
Apoptotic cells were distributed fairly evenly throughout each crypt in each of the deoxycholate-stressed tissue samples, as assessed using strictly morphologic criteria (Figures 2A and 2B). In these strictly morphologic measurements, 1-μm epoxy polychrome-stained sections were used, because it is harder to visualize early condensation of chromatin in 4-μm, paraffin-embedded sections (Figures 2C-2F).
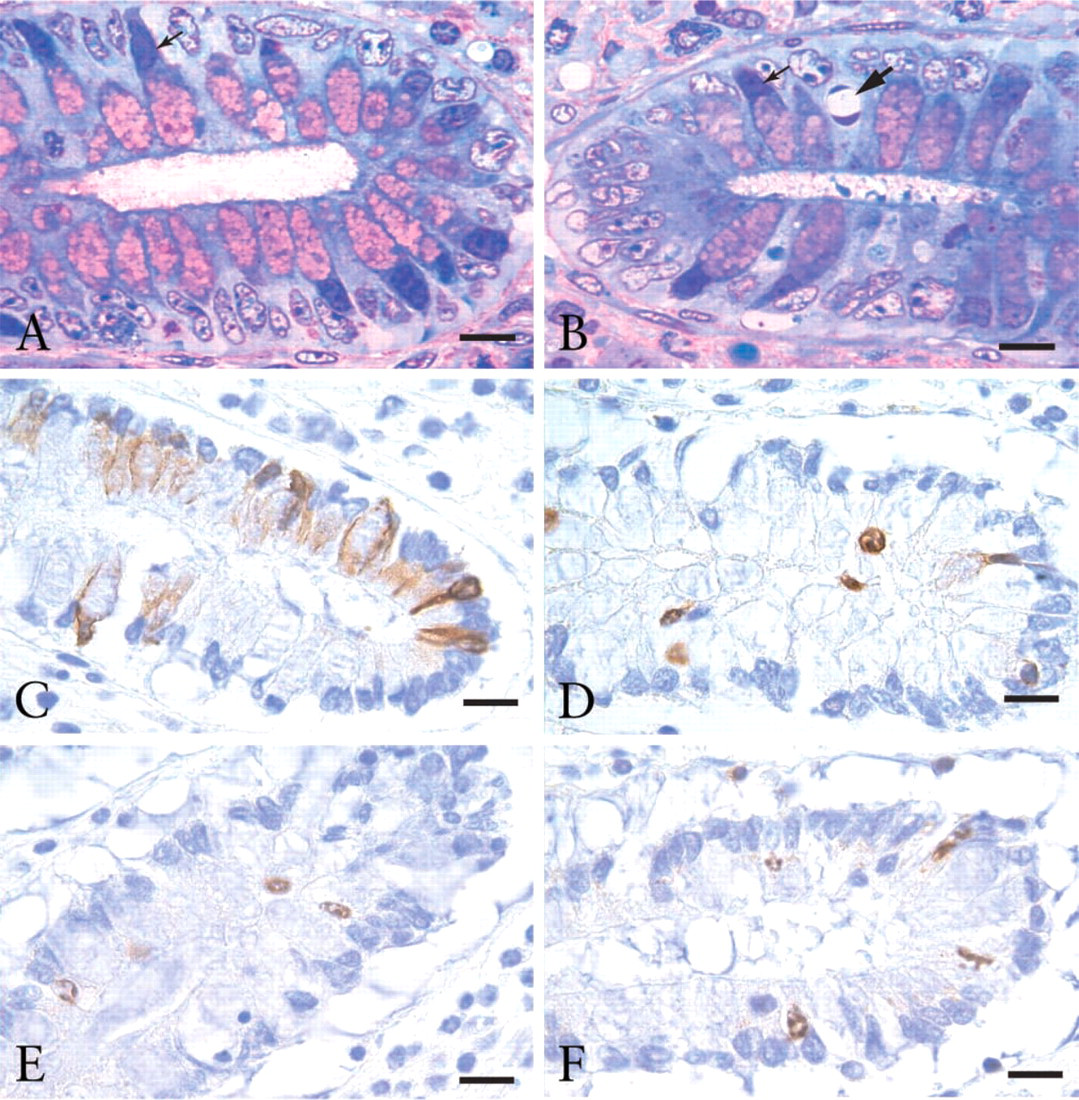
Apoptotic cells within the basal one-fourth of crypts from the flat colonic mucosa subjected to ex vivo deoxycholate-induced stress. In (
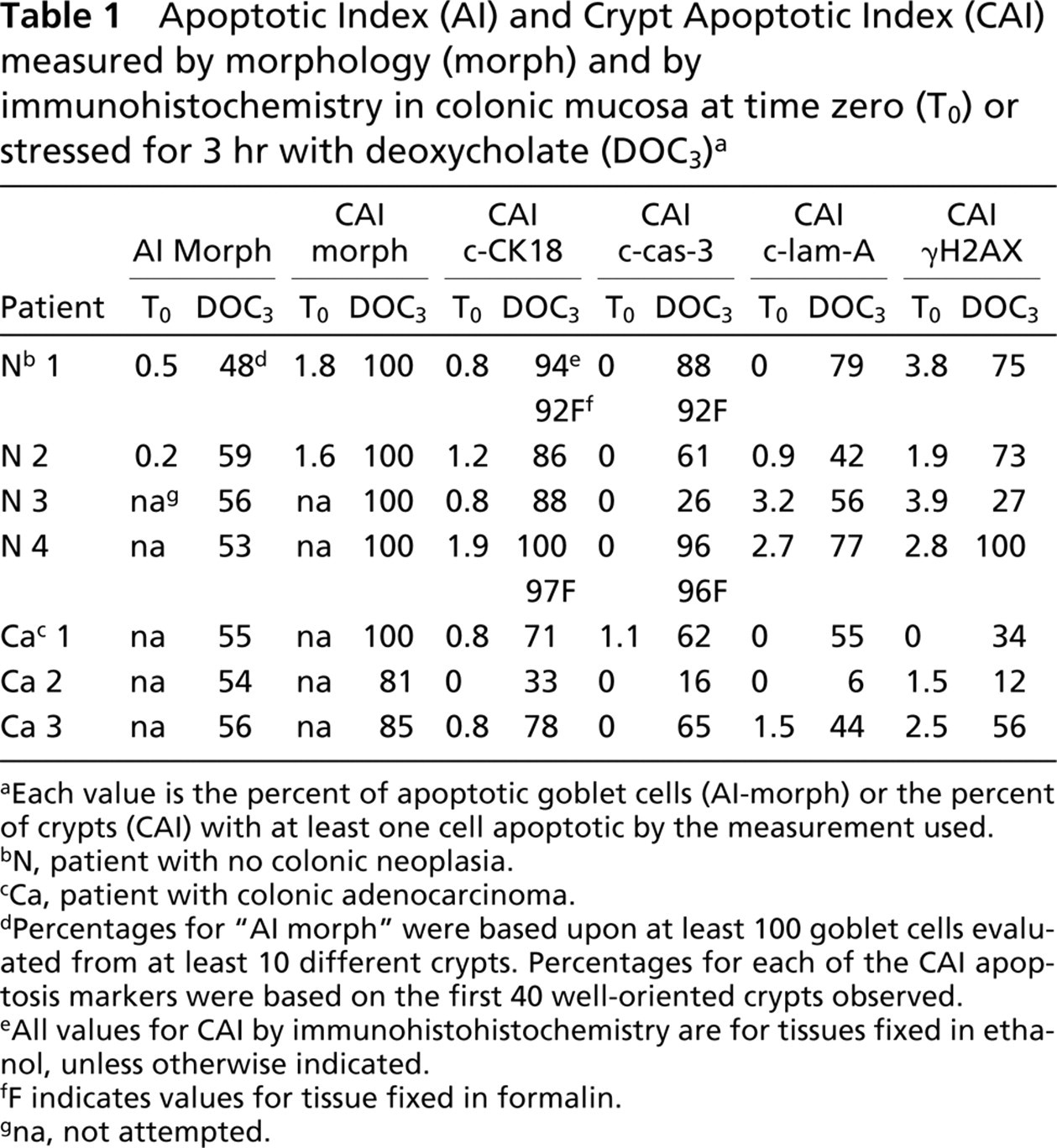
The colonic epithelial cells from subjects without colonic neoplasia are sensitive to NaDOC-induced apoptosis, whereas for approximately half of colon cancer patients, normal-appearing colonic epithelial cells of the flat mucosa are relatively resistant to apoptosis (Bernstein et al. 1999,2002). The resistance to apoptosis tends to occur in patches, with other areas having normal levels of apoptosis (Bernstein et al. 1999,2002). Tissue samples from the three patients with adenocarcinoma and the four patients without colonic neoplasia were stressed ex vivo with deoxy-cholate, embedded in epoxy resin, and polychrome stained. Tissues from each of the seven patients were found to have relatively high levels of AI, measured strictly by morphology (Table 1). Overall, the deoxy-cholate-stressed tissue samples from patients with colons free of neoplasia, as well as from the patients with colonic adenocarcinoma, had a mean AI of 54%, based on our standard goblet cell apoptotic counts (Bernstein et al. 1999). Tissues from two of the patients without colonic neoplasia (N1 and N2 in Table 1) were also evaluated for apoptosis at time zero, before stress was applied. As shown in Table 1, apoptosis evaluated strictly by morphology gave very low background values of AI of 0.5% and 0.2%, and of CAI of 1.8% and 1.6%. For each of the patients, multiple samples treated in the same way showed little variation from sample to sample.
Incubation of tissue in media alone, without Na-DOC present, was also stressful, causing considerable apoptosis. The AIs were evaluated for two samples of tissue taken from each of two patients with non-neoplastic colons, where the tissues were incubated in media alone for 3 hr. In parallel, AIs were evaluated for two further samples of tissue from the same two patients, where the tissues were incubated in media plus 1.0 mM NaDOC for 3 hr. For these two patients, the mean AI for tissue incubated in media alone was 67% of the mean AI for tissue incubated in the presence of NaDOC for 3 hr. Similarly, CAI values were obtained for tissues from patients without colonic neoplasia, patients N1 and N4 listed in Table 1, stressed either by incubation in media alone, or stressed by media plus 1.0 mM NaDOC, and fixed in ethanol. These CAI values were obtained using antibodies to c-CK18 and c-cas-3. The CAI values obtained with c-CK18 were, on average, 82% as high after 3 hr incubation in media alone compared with incubation in media plus NaDOC. The CAI values obtained with c-cas-3 were, on average, 72% as high after 3 hr incubation in media alone compared with incubation in media plus NaDOC.
Apoptotic Index (AI) and Crypt Apoptotic Index (CAI) measured by morphology (morph) and by immunohistochemistry in colonic mucosa at time zero (To) or stressed for 3 hr with deoxycholate (DOC3) a
Each value is the percent of apoptotic goblet cells (Al-morph) or the percent of crypts (CAI) with at least one cell apoptotic by the measurement used.
N, patient with no colonic neoplasia.
Ca, patient with colonic adenocarcinoma.
Percentages for “AI morph” were based upon at least 100 goblet cells evaluated from at least 10 different crypts. Percentages for each of the CAI apoptosis markers were based on the first 40 well-oriented crypts observed.
AII values for CAI by immunohistohistochemistry are for tissues fixed in ethanol, unless otherwise indicated.
F indicates values for tissue fixed in formalin.
na, not attempted.
Apoptosis after stress by incubation in media with NaDOC, quantitated by strictly morphologic criteria and by four of the immunohistochemical markers (c-CK18, c-cas-3, c-lam-A, and γH2AX), yielded fairly high values, and these were quantitated, along with immunohistochemical values obtained at time zero, as controls for background (Table 1). On the other hand, the antibody to c-PARP was considerably less useful for marking deoxycholate-induced apoptotic cells in colonic mucosa, and the antibody to AIF was fairly nonspecific under our conditions. Therefore, these latter two markers were not quantitated in this study. Table 1 gives, for all seven patients, the strictly morphologic AIs and CAIs (using 1-μm epoxy sections), plus the immunohistochemical CAIs (using 4-μm sections) for the four most reactive apoptosis markers.
The time zero values for all four of the immunohistochemical markers quantitated were very low, varying from 0% to 3.9%. The immunohistochemical CAIs obtained after tissues were incubated for 3 hr in media with 1.0 mM NaDOC were generally lower than the strictly morphologic CAIs for such stressed tissues and were much above the background values obtained at time zero (Table 1). The values were close to the same whether tissues were fixed in ethanol or in formalin (Table 1). However, the CAIs for c-CK18 came, on average, closest to the strictly morphologic CAIs. The strictly morphologic CAIs averaged 95% and the c-CK18 CAIs averaged 79%. For c-cas-3, c-lam-A, and γ-H2AX, the average CAI values were 59%, 51%, and 54%, respectively. Figures 2C-2F show images indicating the presence of c-CK18, c-cas-3, c-lam-A, and γ-H2AX, respectively, in typical 4-μm paraffin-embedded samples of deoxycholate-stressed colonic tissue.
The c-CK18 CAI of individuals without colonic neoplasia varied from 86% to 100%, while the CAI of individuals with colonic neoplasia varied, in this study, from 33% to 78%.
In one of the patients (AdCa 3 in Table 1), the majority of apoptotic cells seen were non-goblet cells. Such non-goblet cells undergoing apoptosis were also observed in colonic mucosal tissue from the other two patients with adenocarcinoma (see Figure 2B, where one apoptotic non-goblet cell is shown), as well as in colonic mucosa of patients without colonic neoplasia but at lower frequencies.
For c-cas-3, c-lam-A, and γ-H2AX, the CAI values were variable, falling between 26% and 100% for individuals without colonic neoplasia and between 6% and 65% for individuals with a colonic adenocarcinoma (Table 1).
For each of the antibodies specific for c-CK18, c-cas-3, c-lam-A, and γHAX, only morphologically identifiable apoptotic cells showed a positive reaction, verifying that these antibodies are specific for apoptosis. Each of these antibodies reacted positively with both goblet cells and non-goblet cells which were apoptotic by morphology in the 4-μm paraffin-embedded sections. Figures 2C-2F show crypt basal regions immunohistochemically stained for c-CK18, c-cas-3, c-lam-A, and γH2AX, respectively. The c-CK18 antibody reacted positively with nearly all observed apoptotic cells. However, each of the antibodies against c-cas-3, c-lam-A, and γH2AX failed to react positively with a significant fraction of cells that were clearly identified as apoptotic using strictly morphologic criteria. To clearly identify apoptotic cells by strictly morphologic criteria, it was necessary to focus up and down through the planes of focus in the 4-μm immunohistochemically stained sections. When examined microscopically, it is clear that in deoxycholate-treated colonic tissue, antibodies against c-cas-3, c-lam-A, and γH2AX are less useful for detection of apoptosis than either the c-CK18 antibody or morphology alone.
Discussion
CK18 is an acidic intermediate filament protein that provides structural support for the cell and is found in most simple (glandular) epithelia (Bosch et al. 1988; Caulin et al. 1997; Chu and Weiss 2002). Antibody M30 detects a 10-residue epitope of CK18 (amino acid positions 387-396) on the liberated C terminus as a result of cleavage at the caspase cleavage site DALD-S (Leers et al. 1999). This CK18 neo-epitope is exposed during early apoptosis (Leers et al. 1999; Carr 2000). However, c-CK18 may only be a good marker of apoptosis in the specific cell types in which CK18 occurs. The M30 antibody appears promising for distinguishing patients at high vs. low risk for colon cancer, inasmuch as c-CK18 staining was significantly higher in the non-neoplastic colonic mucosa (CAI of 92.0%) than in the mucosa of those with an adenocarcinoma (CAI 60.7%).
The antibody to c-cas-3 identified apoptotic cells at a frequency lower than the antibody to c-CK18. It is possible that the epitope for c-cas-3 is affected more adversely than the epitope of c-CK18 by the alcohol and heat treatments used. This explanation is likely because we (and others) have shown that caspase-3 is cleaved in deoxycholate-treated cells of colonic origin (Schlottman et al. 2000; Washo-Stultz et al. 2002). On the other hand, while caspase-3 is a major protease involved in the execution phase of apoptosis (Slee et al. 2001), c-cas-3 may not be activated in some ex vivo deoxycholate-treated cells from the colonic mucosa in which CK18 is cleaved, or it may initially be present at lower levels than CK18. Activated c-cas-3 cleaves many substrates, including CK18 [although CK18 is also cleaved by caspases-6 and −7 (Caulin et al. 1997), as well as by the DEDD-procaspase-3 complex (Lee et al. 2002)]. Cleaved caspase-3 antibody detects the activated form of caspase-3 by specifically recognizing the large fragment (17-20 kD) that results from cleavage after Asp175. The antibody to c-cas-3 has been reported to be useful for detecting apoptotic cells in archival paraffin sections (Gown and Willingham 2002). In the present study, we evaluated the specificity and usefulness of the c-cas-3 antibody for distinguishing the normal mucosa of patients without colonic neoplasia from that of patients with colon cancer. The mean CAI in the mucosa of the four individuals without neoplasia [67.8% (±16.0% SEM)] was higher than the mean for the mucosa of the three individuals with adenocarcinoma [47.7% (±15.7% SEM)], but the difference was not statistically significant.
Lamin A is an intermediate filament protein and a major component of the nuclear lamina (Moir and Spann 2001). The nuclear lamins polymerize to form the nuclear lamina, a fibrous structure on the inner face of the nuclear membrane. The lamins also form structures within the nucleoplasm that help maintain the shape of the nucleus and participate in various nuclear processes (Moir and Spann 2001). C-lam-A antibody detects the large (40-45 kD) fragment of lamin A after c-cas-6 cleaves the protein at Asp230. The mean CAI for c-lam-A in the mucosa of the individuals without neoplasia was 63.5% (±8.7% SEM), higher than the mean for the mucosa of the individuals with adenocarcinoma [35.0% (±15.3% SEM)], but the difference was, again, not significant.
H2AX is a member of the H2A histone family that becomes rapidly phosphorylated (γH2AX) by ATM at Ser139 in response to DNA double-strand breaks (Rogakou et al. 2000; Burma et al. 2001). Extensive phosphorylation of H2AX appears to be an early chromatin modification following initiation of DNA fragmentation (Rogakou et al. 2000; Burma et al. 2001). The antibody used here recognizes γH2AX phosphorylated at Ser139 in the carboxy terminus. The mean CAI for γH2AX-positive cells in the mucosa of the four individuals without neoplasia was 68.8% (±15.2% SEM), higher than the mean for the mucosa of individuals with adenocarcinoma [34.0% (±12.8% SEM)], but the difference, again, was not significant.
In conclusion, strictly morphologic assessment was the most useful method for detecting colonic crypt apoptotic cells in deoxycholate-stressed tissue. c-CK18 was specific for apoptosis and nearly as useful as morphologic measurements when tissue from patients without neoplasia was evaluated but showed a lower level of identifying apoptosis in patients with a colonic neoplasia. This is interesting as a possible biomarker in stressed tissue because the cancer cases that were prospectively obtained for this apoptosis marker study were “normal” using our standard apoptosis goblet cell AI score (Bernstein et al. 1999). Reduced c-CK18 reactivity may prove useful as an intermediate biomarker for cancer risk; however, a larger study is necessary to validate this point. Antibodies to c-cas-3, c-lam-A, and γH2AX were specific for identification of apoptotic cells of the colonic epithelium but were less useful than morphologic assessment or c-CK18 for identifying apoptotic cells. The specific antibodies to detect c-PARP and AIF appear to be inefficient for assessing apoptosis in alcohol-fixed, paraffin-embedded colonic epithelium.
Footnotes
Acknowledgements
This work was supported by National Institutes of Health Grant P01 CA-72008, Gastrointestinal Specialized Program of Research Excellence Grant IP50 CA-95060, AD-CRC Grants 10016 and 6002, and VA Merit Review Grant No. 0114.