
Abstract
Epidermal growth factor (EGF) in rat salivary glands is regulated by testosterone, thyroxin, and growth hormone (GH). Salivary glands of 45-day-old giant and dwarf male and female transgenic mice were examined histologically and by immunohistochemistry (IHC) for EGF. Male giants showed no significant differences from wild-type (WT) parotid and submandibular glands. However, their sublingual glands expressed EGF diffusely and strongly in granular cells within the striated ducts, where they were not found in WT mice. Submandibular gland ducts of female WT were different, having individual granular cells strongly positive for EGF and distributed sporadically along the striated duct walls. Neither female GH-antagonist dwarf mice nor GH-receptor knockout mice had any granular cells expressing EGF in any gland. Obvious presence of granular duct cells in the sublingual glands of giant male mice suggests GH-upregulated granular cell EGF expression. Furthermore, absence of granular duct cells from all glands in female GH-antagonist and GH-receptor knockout transgenic mice suggests that GH is necessary for the differentiation of the granular cell phenotype in female salivary glands.
T
The aim of this study was to determine if excess GH secretion in giant mice affects hypertrophy of the granular cell phenotype in the salivary glands. Conversely, competition of bovine growth hormone antagonist with endogenous GH in the GH-antagonist dwarf mouse would induce failure of granular cell development in striated ducts. Moreover, absence of growth hormone receptor (GHr) in GHr-deleted dwarf mice would result in absence of the granular cell phenotype.
Materials and Methods
Animals
Giant growth hormone excess mice (GHXs) overexpress a bovine growth hormone (bGH) transgene under the control of a mouse metallothionein-I (MT) promoter, MT-bGH. Background is Bb/SJL (Palmiter et al. 1982,1983).
Dwarf antagonist mice (GHAnt) overexpress a mutant bovine growth hormone GH antagonist in which a lysine is substituted for glycine 119 (bGH-G119K). Background is C57Bl/b5 (Chen et al. 1997).
Dwarf growth hormone receptor knockout mice (GHr KO) are homozygous for a gene deletion of the receptor gene created by insertion of a neo cassette into exon four. Background is C57Bl/b5 (Zhou et al. 1997).
Wild-type, control, or heterozygote littermates of these transgenic models of GH action were used for comparison with the above treatment groups (Kopchick et al. 1999).
Generation of Mice
The genotypes of littermates were determined soon after birth by PCR using DNA from tail tips. The mice were maintained in specific pathogen-free conditions and fed ad libitum on mice pellets.
All animal experimentation was carried out in accordance with National Health and Medical Research Council of Australia guidelines and was approved by the University of Queensland Animal Ethics Committee.
The study was undertaken on the following groups of mice; from giant mice litters, at least 12 animals, three male and three female WT littermates; from GH antagonist mice litters, at least 12 animals, three male and three female GHAnt mice and and three male and three female WT littermates; from Gh receptor knockout mice litters, at least 18 animals, three male and three female honozygous GHR-KO mice and three male and three female WT littermates.
Tissue Preparation
Animals were sacrificed by decapitation under CO2 asphyxiation. The heads were totally immersed in tissue fixation solution, i.e., 4% paraformaldehyde in PBS for immunohistochemistry (IHC). Fixation was at 4C for 24 hr. The submandibular, sublingual, and parotid glands were dissected free, immersed in 70% ethyl alcohol, and processed for paraffin embedding. The glands were oriented to be cut in the parasagittal plane. Five-μm sections were obtained, attached to positively charged glass slides (polylysine), and stained with hematoxylin and eosin (H and E), Mallory phosphotungstic acid hematoxylin (PTAH), and EGF IHC, as follows.
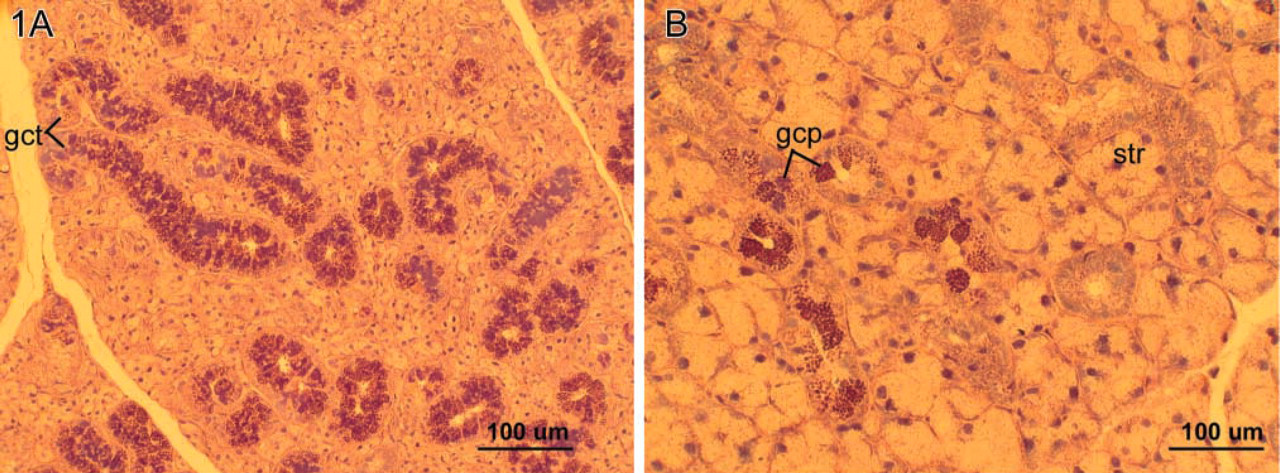
To differentiate the acinar and ductal elements of the mouse submandibular, sublingual (mucous), and parotid (serous) glands, PTAH stain was used (Lillie 1965). Briefly, after deparaffinization, sections were immersed in 95% alcohol for 5 min, followed by 0.5% sodium thiosulfate for another 5 min. Sections were then stained in phosphotungstic acid/hematoxylin for 17 hr. Finally, they were dehydrated in 95% and 100% alcohol, passed through xylene, and mounted. This stain gave excellent discrimination of striated ducts by staining the mitochondria of the basal striations blue. It also stained the granules of the cells of the granular convoluted tubules (GCTs) within the submandibular glands variously, as dark purple, blue, or red spheres or irregular blue lakes (Figure 1). Accordingly, the criteria for the presence of the granular cell phenotype in this study were intense eosinophilia in H- and E-stained sections and dark purple, blue, or red spheres in PTAH-stained sections. The relative number and distribution of this granular cell phenotype, which varies with developmental age and sex of the mouse, were studied for changes relative to GH status.
The expression of EGF in the salivary gland and specifically by the granular cell phenotype was studied by IHC employing an anti-EGF antibody (SIGMA E-2635; Sigma, St Louis, MO) diluted 1:100. Briefly, after deparaffinization, hydration, and washing in PBS, the sections were exposed to 3% hydrogen peroxidase for 15 min. Next, the sections were incubated with 1:10 swine serum (code ×0901; Sigma) for 30 min to block nonspecific protein binding. After incubation, excess serum was blotted and the sections were incubated with the primary antibody for 90 min. Specificity of EGF staining was checked on negative control sections by omission of the primary antibody. After two 3-min washes with PBS containing 0.1% Triton (SIGMA T-9284), the sections were incubated with biotinylated secondary antibody (biotinylated swine anti-mouse, anti-rabbit, and anti-goat (DAKO LSAB-kit; DAKO, Carpinteria, CA) for 15 min. The sections were then rinsed in PBS containing 0.1% Triton and incubated with streptavidin-peroxidase complex (DAKO LSAB-kit) for 15 min. The peroxidase activity was visualized with 3,3'-diaminobenzidine (DAB) solution for 3 min and counterstained with hematoxylin for 30 sec. Finally, sections were dehydrated through alcohol, cleared in xylene, and coverslipped.
Histological Evaluation
Sections stained with H and E and with PTAH were evaluated by conventional light microscopy for the following features:
The distribution of GCTs or of the granular cell phenotype in association with striated ducts of submandibular glands.
(
(
(
The presence or absence of the granular cell phenotype in either the sublingual (mucous) or the parotid (serous) glands where this phenotype is not normally present.
Sections stained for EGF by IHC were evaluated by conventional light microscopy to record the cellular expression of the stain and to give a qualitative estimate of the relative intensity of immunostaining in the duct cells. Grades assigned were –, undetectable; +, weak; 2+, strong; or 3–4 +, very strong.
All sections of each animal were examined and graded, without knowledge of treatment by one of the authors (WGY). When the results of all glands from all animals in the study were complete, the codes were broken and the observations were compared among littermates and among treatment groups.
Results
Histology
Wild-type male mouse salivary glands showed no appreciable variations among the strains of mice in the differentiation of the several duct types. Their submandibular glands had extensive differentiation of GCTs and the adjacent striated ducts were readily identified. These components stained positively for EGF (Tables 1–3). In the sublingual (mucous) and parotid (serous) glands (which lacked GCTs) the striated ducts stained consistently for EGF.
The male pattern of the GCT differs from the female pattern of GCT in the distribution of granular cell phenotypes. In all male mice, granular cells that lined the GCT were uniformly granular and contiguous. Their granules were strongly eosinophilic and stained red or blue with PTHA (Figure 1A). WT female mouse salivary glands showed no appreciable differences among the strains from the female pattern of distribution of granular duct cells. In controls, granular cells were found in discrete groups or singly along the lining of the striated ducts in the submandibular gland (Figure 1B). Whereas the cytoplasm of the striated duct cells stained diffusely for EGF, the granular cell granules stained more intensely by IHC. However, such intensely stained granular cells were not normally found in the duct lining cells of the striated ducts of the sublingual (mucous) glands or in the parotid (serous) glands. The contents of capillary blood vessels were also EGF-positive.
No histological differences were discriminated in the salivary glands between the WT and their heterozygous littermates bearing the knockout gene in both male and female animals, nor was the intensity of EGF staining appreciably different among the littermates of the same sex. Female heterozygotes exhibited the same groupings of the phenotypic granular cells positive for EGF as the WT in the submandibular glands.
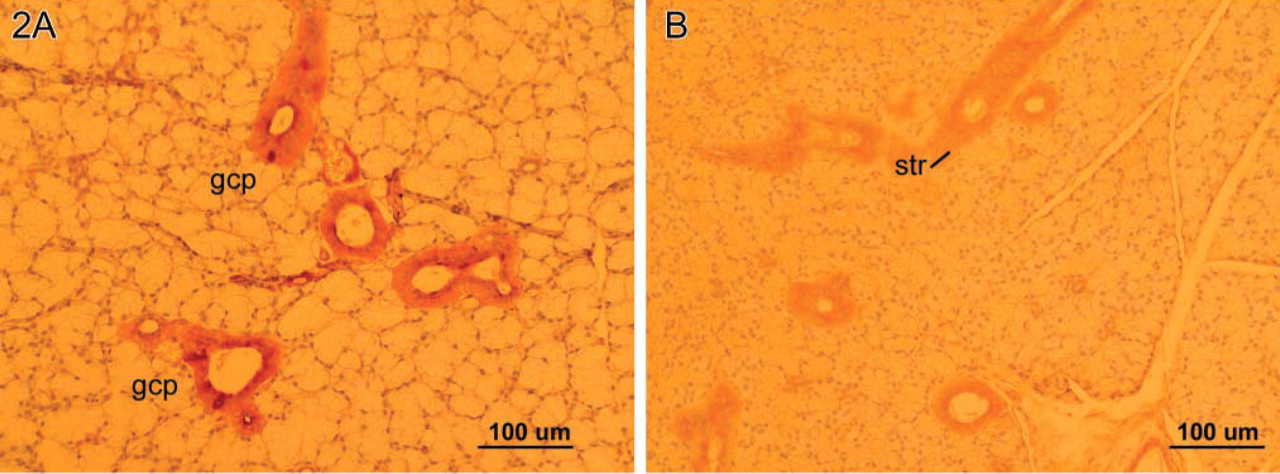
The male giant (GHXs) mice submandibular glands exhibited the male pattern of GCTs with abundant eosinophilic and blue granules with PTAH (Figure 1A). No quantifiable differences from the WT pattern were appreciated in sections stained for EGF by IHC. The sublingual (mucous) glands in male giant mice, however, were remarkable in that granular cell phenotypes were identified with H and E, PTAH, and EGF IHC (Figure 2A). This granular cell phenotype was not found in the sublingual glands of any of the wild strains nor in those of the heterozygote littermates of the KO strain (Figure 2B). Moreover, in the female giant mice this cell phenotype was absent from the striated ducts of the sublingual (mucous) glands in all animals. No instances of the granular cell phenotype were found in parotid gland striated ducts in any of the treatment groups.
Epidermal growth factor expression in GH excess mice
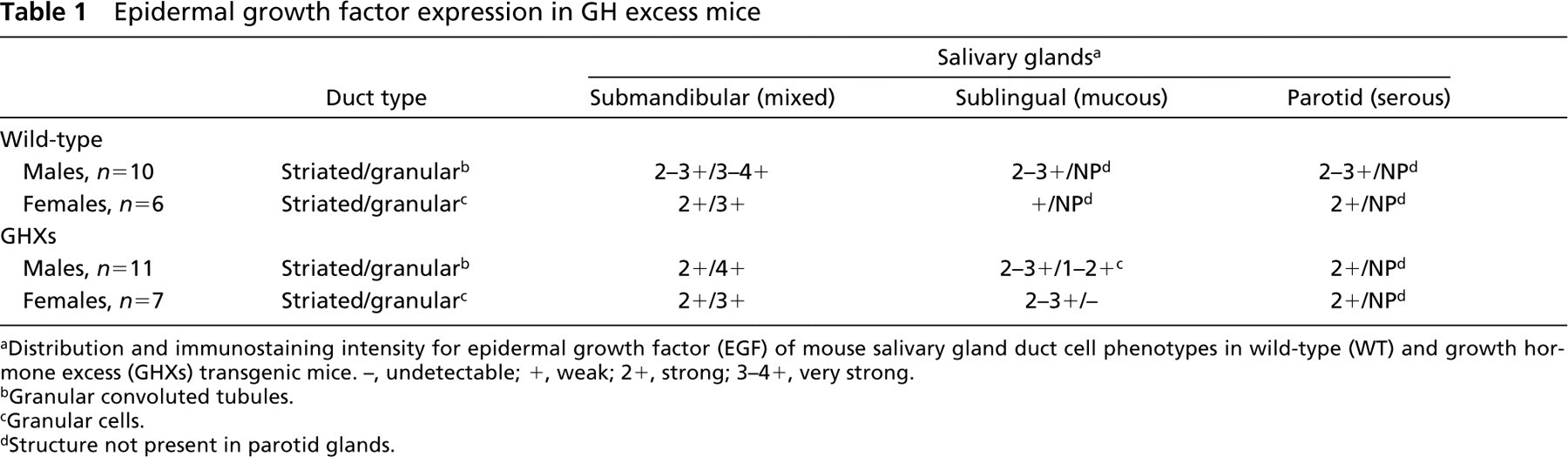
Distribution and immunostaining intensity for epidermal growth factor (EGF) of mouse salivary gland duct cell phenotypes in wild-type (WT) and growth hormone excess (GHXs) transgenic mice. –, undetectable; +, weak; 2+, strong; 3–4+, very strong.
Granular convoluted tubules.
Granular cells.
Structure not present in parotid glands.
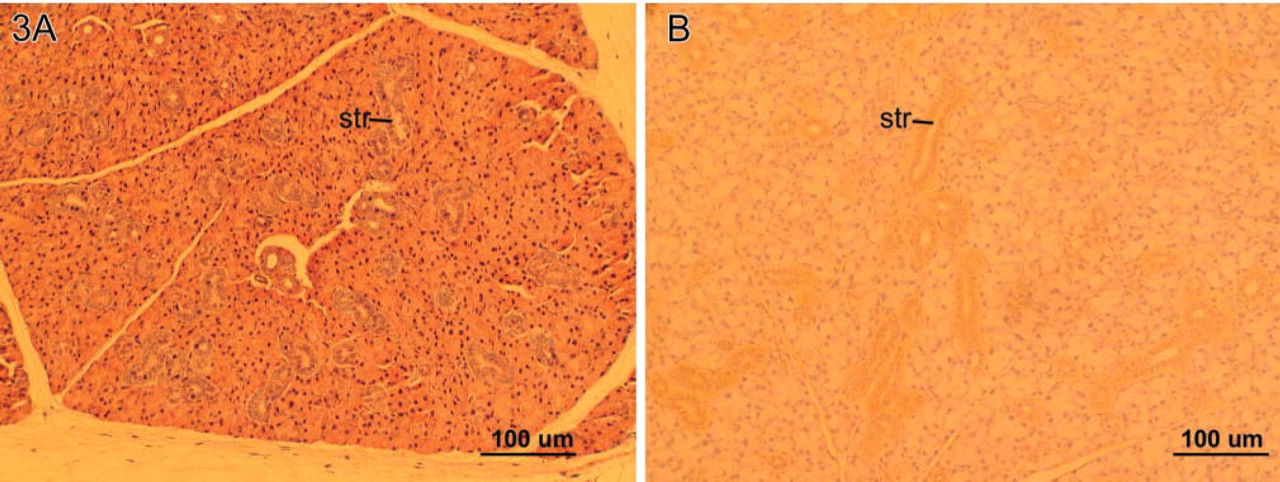
The male patterns of GCT in the salivary glands of GHAnt dwarf mice were not appreciably different from those of their WT littermates, although the glands were approximately one third of normal size. In contrast, in female dwarf GHAnt mice the granular cell phenotype was absent from the striated ducts of the submandibular glands (Figure 3A), nor were granular cells identified in their sublingual or parotid glands. EGF was only weakly expressed throughout the striated ducts in all glands of GHAnt mice (Table 3). Although the salivary glands of the male homozygous GHr KO mice were about one third the size of their WT and heterozygous littermates, the GCTs were normal and of the normal male pattern. Similarly, the granular cells were eosinophilic with H and E, had red and purple granules in PTAH, and stained strongly for EGF. Striated ducts were readily identifiable in the submandibular, sublingual, and parotid glands, and these ducts were stained diffusely for EGF. No granular cells were found in the latter two glands.
Epidermal growth factor expression in GH antagonist mice

Distribution and immunostaining for epidermal growth factor (EGF) intensity of antagonist (GHAnt) transgenic mice. Intensity as in legend to Table 1.
Granular convoluted tubules.
Granular cells.
Structure not present in parotid glands.
In contrast, in the submandibular salivary glands of the female homozygous GHr KO mice, the granular cell phenotype was absent in all of the glands examined (Figure 3B). Striated duct epithelium of all three gland types in GHr KO female mice stained uniformly weakly for EGF. No examples of the granular cell phenotype positive for EGF were found.
Epidermal growth factor expression in GH receptor knockout mice
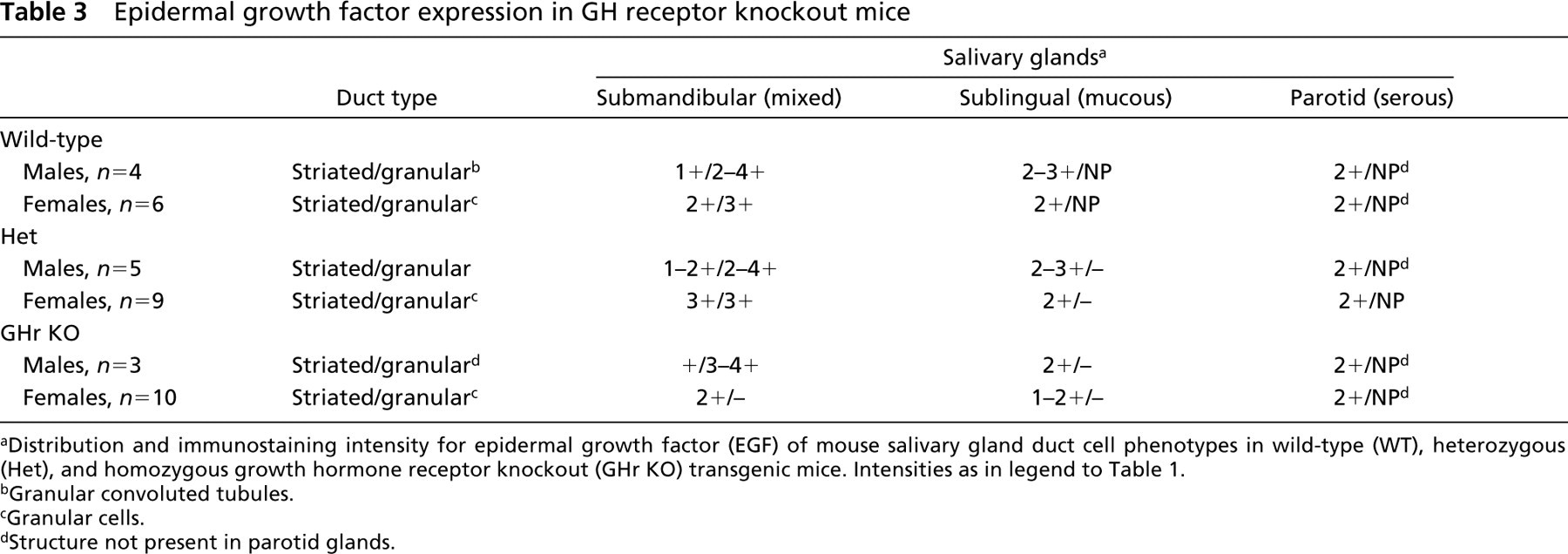
Distribution and immunostaining intensity for epidermal growth factor (EGF) of mouse salivary gland duct cell phenotypes in wild-type (WT), heterozygous (Het), and homozygous growth hormone receptor knockout (GHr KO) transgenic mice. Intensities as in legend to Table 1.
Granular convoluted tubules.
Granular cells.
Structure not present in parotid glands.
Discussion
At 45 days, mouse salivary glands have virtually completed their postnatal histodifferentiation into the highly specialized cells of acini, myoepithelium, GCTs, striated ducts, and excretory ducts (Navia 1977). The GCT, the site of EGF secretion, demonstrates a clear sexual dimorphism (Gresik 1994). In this study, the GCTs in the submandibular glands of all male WT and GHXs mice were fully developed and no morphological differences attributable to the bovine transgene were found. GCTs with similar morphology were found in the submandibular glands of the male dwarf GHAnt and GHr KO mice. However, because these glands were smaller in dwarf mice, total EGF output might be affected.
Suggesting that GH may have an effect on differentiation of the granular cell phenotype was the finding of granular cells in the striated ducts of the sublingual (mucous) glands of giant male mice. This cell phenotype was not found in sublingual glands of WT littermates or in those of the other genetically modified groups. In other studies, occasional cells of the granular cell phenotype have been described in sublingual glands, positive for EGF (Gresik and Barka 1983; Garrett and Anderson 1991; Kurabuchi and Gresik 2001). It may therefore be that excess GH produces a hypertrophy of cells of the granular cell phenotype in sublingual gland ducts. Because this was not found in any of the dwarf mice sublingual glands, it is unlikely to be a compensatory mechanism for lack of granular cell differentiation in the submandibular gland.
In this context, it would be of interest to determine the distribution of growth hormone receptor in the ducts of the three types of mouse salivary gland. If GH does not affect the salivary glands directly through its receptor on target cells, its effect may be indirect through trophic effects on the thyroid glands and possibly on the adrenal cortices or the gonads in males, all of which have strong influences on differentiation of the granular cell phenotype in rodents.
One reason why the absence of GH receptor failed to influence the differentiation of GCTs in either male GHAnt or GHr KO mice is presumably that androgens overcome the lack of growth hormone in the antagonist state, and that GH receptor is not a prerequisite for the androgen effect. Castration of male rats decreases EGF to the same level as that of intact females (Gresik and Barka 1977; Tajima et al. 2000). Hypophysectomized male rats have reduced EGF levels, to about 7% of normal (Gresik et al. 1981). This reduction is partially restored by administration of corticosterone or triiodothyronine (T3) (Gresik et al. 1981; Steidler and Reade 1982,1983). EGF levels respond to the additive effects of testosterone propionate with GH or with T3 or with both together (Hiramatsu et al. 1994; Tajima et al. 2000). These findings, together with those of the present study, suggest that the male GCTs of the submandibular gland depend for their differentiation, and hence for EGF production, on androgens. However, the granular cell phenotype may differentiate under the influence of GH in males, as shown in the sublingual (mucous) glands of male giant mice. Lack of granular cells in the female GHAnt and GHr KO dwarf mice supports the conclusion that GH is involved in differentiation of granular cells in females. Ovariectomy and/or administration of estradiol-17β to ovariectomized rats fails to affect EGF levels in female mice (Gresik 1994). Evidently it is GH that is important for the differentiation of the granular cell phenotype, for the maintenance of this phenotype, or for appearance of the phenotype in the striated ducts of female animals in the course of maturation towards adulthood and the subsequent production of EGF in the female mouse.
EGF was constitutively expressed by the striated duct lining epithelial cells in all glands in all treatments. GH appears to have its effect on the differentiation of granular cell phenotype in females. If GH controls differentiation of this phenotype and also up-regulates the expression of EGF receptor (Ekberg et al. 1989), then a link between GH and EGF in the growth and repair of the gut-associated epithelia can be postulated.
Conclusions
This study determines the effect of GH on the differentiation of the granular cell phenotype in the salivary glands concomitantly with EGF expression by these cells in three different GH transgenic mice models. The presence of granular duct cells in the sublingual glands of giant male mice, not normally present in WT mice, suggests that GH upregulates granular cell EGF expression. Furthermore, the absence of granular duct cells from all glands of female, GH antagonist, and GH receptor knockout transgenic mice suggests that GH is necessary for the differentiation of the granular cell phenotype in female mouse salivary glands.