
Abstract
We studied cell-specific protein expression of all the major antioxidant enzymes (AOEs) and related proteins, such as copper-zinc superoxide dismutase (CuZnSOD), manganese SOD (MnSOD), extracellular SOD (ECSOD), catalase, the heavy and light chains of γ-glutamylcysteine synthetase (γ-GCS-l and γ-GCS-h, also called glutamate cysteine ligase), the rate-limiting enzyme in glutathione synthesis, hemeoxygenase-1 (HO-1), and thioredoxin (Trx), in developing human lung, respiratory distress syndrome, and bronchopulmonary dysplasia by immunohistochemistry. Generally, after 17 weeks of gestational age, MnSOD was predominantly expressed in bronchial epithelium, alveolar epithelium, and macrophages, CuZnSOD was expressed in bronchial epithelium, ECSOD was expressed in bronchial epithelium, vascular endothelium, and the extracellular matrix, catalase was expressed in bronchial epithelium and alveolar macrophages, γ-GCS-h was expressed in bronchial epithelium and endothelium, and γ-GCS-l was expressed in bronchial epithelium. Trx was restricted to bronchial epithelium and to a lesser extent to alveolar macrophages, and HO-1 found in alveolar macrophages. Basically, the expression of these enzymes was similar in normal and diseased lung. It can be concluded that various AOEs and related proteins differ in their distribution and expression in lung before term, but generally it seems that infants are better adapted to high oxygen tension than might be expected.
I
The most important AOEs and related proteins include superoxide-scavenging superoxide dismutases (SODs), such as copper-zinc SOD (CuZnSOD, SOD1), manganese SOD (MnSOD, SOD2), and extracellular SOD (ECSOD, SOD3), and several enzymes that participate in H2O2 degradation, such as glutathione (GSH) peroxidases, catalase, and the small cysteine-containing proteins thioredoxin (Trx) and Trx peroxidases (peroxiredoxins) (Hass and Massaro 1987; Hayashibe et al. 1990; Rickett and Kelly 1990; Das et al. 1999; Nozik-Grayck et al. 2000; Kim et al. 2001). Moreover, hemeoxygenase-1 (HO-1) is induced by hyperoxia and has been claimed to protect lung cells against high oxygen tension and oxidants (Otterbein et al. 1995). Previous studies in experimental animals have indicated increased expression of these enzymes during lung morphogenesis. Some of these enzymes, such as ECSOD (Nozik-Grayck et al. 2000) and Trx (Das et al. 1999), have also been shown to be induced immediately after birth. Unfortunately, very few such investigations have been done on human lung, and the results of these studies are somewhat conflicting (Strange et al. 1988,1990; McElroy et al. 1992; Dobashi et al. 1993; Asikainen et al. 1998,2001). Therefore, we analyzed the cell-specific protein expression of the major AOEs and related proteins, CuZnSOD, MnSOD, ECSOD, catalase, the heavy and light chains of γ-glutamylcysteine synthetase (i.e., glutamate cysteine ligase; γ-GCS-l and γ-GCS-h), the rate-limiting enzyme in GSH synthesis, HO-1, and Trx in developing human lung, RDS, and BPD.
Materials and Methods
Patients and Handling of Specimens
Samples of lung tissue were retrieved from the files of the Department of Pathology, Oulu University Hospital. The study protocol was approved by the Ethical Committee of the local hospital. The study material for developing lung consisted of two cases of spontaneous abortion, five cases of stillbirth, and four cases of autopsied infants who had died within 2 days after birth; for infant lung diseases, the materials were from six patients with RDS and five patients with BPD with typical morphological findings (Rosan 1975; Hussain et al. 1998). The gestational ages of the infants with normal lungs ranged from 17 to 40 weeks, corresponding to the canalicular (weeks 17–28, four cases), saccular (weeks 29–35, four cases), and alveolar (weeks 36–40, three cases) periods. None of the cases studied involved pneumonia, cardiac abnormalities, or features of maceration. The autopsies had been performed within 2.5 days (18 cases within 1 day, three cases within 2 days, one case within 2.5 days). Clinical information was obtained from patient records. The clinical characteristics of the infants with normal developing lung are shown in Table 1, and those of infants with RDS and BPD are shown in Table 2.
Lung tissues from either right or left lung removed at autopsy were fixed in 10% formalin, dehydrated, and embedded in paraffin, and 4-μm sections were stained with hematoxylin and eosin. The whole material was reevaluated, and one representative tissue block from each case was selected for immunohistochemical studies. To identify the various lung cells, the sections were also stained with commercially available antibodies against α-smooth muscle actin (clone 1A4 from Sigma Bio-Sciences, St. Louis, MO; diluted 1:50), CD31 (clone JC/70A from Dako, Glostrup, Denmark; diluted 1:40), CD68 (clone PG-M1 from Dako; diluted 1:50), and myeloperoxidase (clone A 398 from Dako; diluted 1:100). CD31 is characterized by its strong reactivity with a formalin-resistant epitope in endothelial cells in normal tissues and in benign and malignant proliferations. CD68 stains macrophages in a wide variety of human tissues, including lung.
Clinical characteristics of patients with normal developing lung

GA, gestational age (weeks plus days).
Clinical characteristics of newborns with RDS and BPD

ICH, intracerebral hemorrhage; IVH, intraventricular hemorrhage.
Immunohistochemistry
Four-μm-thick sections were cut from a representative paraffin block. The sections were first deparaffinized in xylene and rehydrated in a descending ethanol series. To enhance immunoreactivity, the sections were incubated in 10 mM citrate buffer (pH 6.0) and boiled in a microwave oven for 2 min at 850 W and then for 8 min at 350 W. The paraffin blocks were then incubated with 2% milk powder to diminish background staining. Endogenous peroxidase activity was eliminated by incubation in 0.1% H2O2 in absolute methanol for 10 min. After incubation with the specific antibody overnight at +4C, a biotinylated secondary anti-rabbit or anti-mouse antibody was applied followed by the avidin-biotin peroxidase complex (both from Dakopatts; Glostrup, Denmark). The color was developed using aminoethylcarbazol (AEC), and the sections were lightly counterstained with hematoxylin and mounted with Eukitt (Kindler; Freiburg, Germany). Replacement of the primary antibody by PBS at pH 7.2 and rabbit or mouse primary antibody isotype control (Zymed Laboratories; San Francisco, CA) was used as a negative control.
Antibodies
Polyclonal rabbit antibodies for MnSOD (at a dilution of 1:1000), ECSOD (1:100), CuZnSOD (1:2000), and catalase (1:400) were a gift from Prof James D. Crapo (National Jewish Medical and Research Center, Denver, CO) and have been shown to be specific for those enzymes (Slot et al. 1986; Kinnula et al. 1994). The antibody against HO-1 (1:100) has been characterized (clone 23; Transduction Laboratories, Lexington, KY) and is commercially available. The affinity-purified rabbit anti-human Trx antibody (1:1000) has been characterized previously (Sahaf et al. 1997). Rabbit antibodies against γ-GCS-h (1:1000) and γ-GCS-l (1:1000) peptides were gifts from Dr T Kawanagh (University of Washington, Seattle) and are specific for γ-GCS, as shown previously (Soini et al. 2001b). In the negative controls, the primary antibodies were substituted with PBS (pH 7.2) or serum isotype control (Zymed Laboratories). The specificity of all these antibodies has been confirmed previously not only by the appropriate negative controls but also by Western blot analysis (Slot et al. 1986; Kinnula et al. 1994; Sahaf et al. 1997; Lakari et al. 2001; Soini et al. 2001a,b).
Scoring of the Immunoreactivity
The extent of the immunoreactivity for various AOEs was evaluated semiquantitatively as negative (0) or positive (+) in different types of pulmonary cells, such as epithelial cells of bronchioli and bronchi, alveolar epithelium (i.e., pneumocytes), alveolar macrophages, endothelial cells, interstitial cells, chondrocytes, mesothelium, and submesothelial connective tissue cells. Extracellular tissue of the lung parenchyme was also evaluated.
Results
CuZnSOD
Bronchial epithelial cells showed similar CuZnSOD positivity in the developing normal lung, RDS, and BPD (Tables 3, 4, and 5; Figure 1A). Alveolar macrophages were negative with the exception of BPD, which showed a weak positivity in four cases. Alveolar epithelium, endothelium, and mesothelium were also negative.
MnSOD
The immunoreactivity in bronchial epithelium was positive in the normal developing lung, RDS, and BPD (Figure 1B). Types I and II pneumocytes were positive in normal lung, RDS, and in three cases of BPD. Alveolar macrophages were negative in seven and positive in four cases of normal lung (weeks 35–40). Alveolar macrophages were positive in all cases with RDS and BPD. A positive immunoreactivity for MnSOD was observed also in smooth muscle cells of bronchi and arteries, epithelium of bronchial glands, chondrocytes, interstitial cells, endothelium of arteries and veins, mesothelial cells, and submesothelial connective tissue cells in normal lung, RDS, and BPD.
ECSOD
ECSOD was expressed in the epithelium of bronchus and bronchiolus in normal developing lung in every case studied. ECSOD was expressed extracellularly during all developmental periods, its expression being most prominent in the vessel wall (Figure 1C). A positive extracellular expression for ECSOD was observed also in the walls of bronchi from week 25 onward. It was expressed in intima, including endothelium, and media of arteries during all developmental stages, but not in veins, alveolar interstitium, submesothelial connective tissue of pleura, or mesothelial cells. In RDS, ECSOD was expressed in the epithelium of bronchi and bronchioli, chondroid cells, endothelium and media of arteries, and occasionally also in alveolar epithelium. Hyaline membranes overlying detached alveolar epithelium showed positive immunoreactivity. In BPD, the metaplastic alveolar epithelium, the epithelium of bronchi and bronchioli, alveolar interstitium, chondroid cells, and alveolar macrophages showed positive immunoreactivity in most cases. The interstitial cells showing positivity for ECSOD were often localized to the same sites as the cells expressing α-smooth muscle actin, which is known to correspond to interstitial myofibroblast-type cells.
Catalase
Catalase was negative or weakly positive in bronchial epithelium from weeks 17 to 31, whereas its expression was positive from weeks 35 to 40. Alveolar epithelium was negative from weeks 17 to 31, but a positive expression from week 35 toward term was detected. Endothelium of arteries and veins exhibited positive expression from week 31 onward. In RDS, bronchial epithelium, and occasionally alveolar epithelium and endothelium, showed positive immunoreactivity. In BPD, positive immunoreactivity was found in bronchial and alveolar epithelium (Figure 1D). Alveolar macrophages were positive during all developmental periods of normal lung and also in RDS and BPD. Pleural mesothelium and alveolar interstitial cells were negative.
Scoring of the immunohistochemical intensity of various AOEs in normal developing human lung from weeks 17–40, RDS, and BPDa
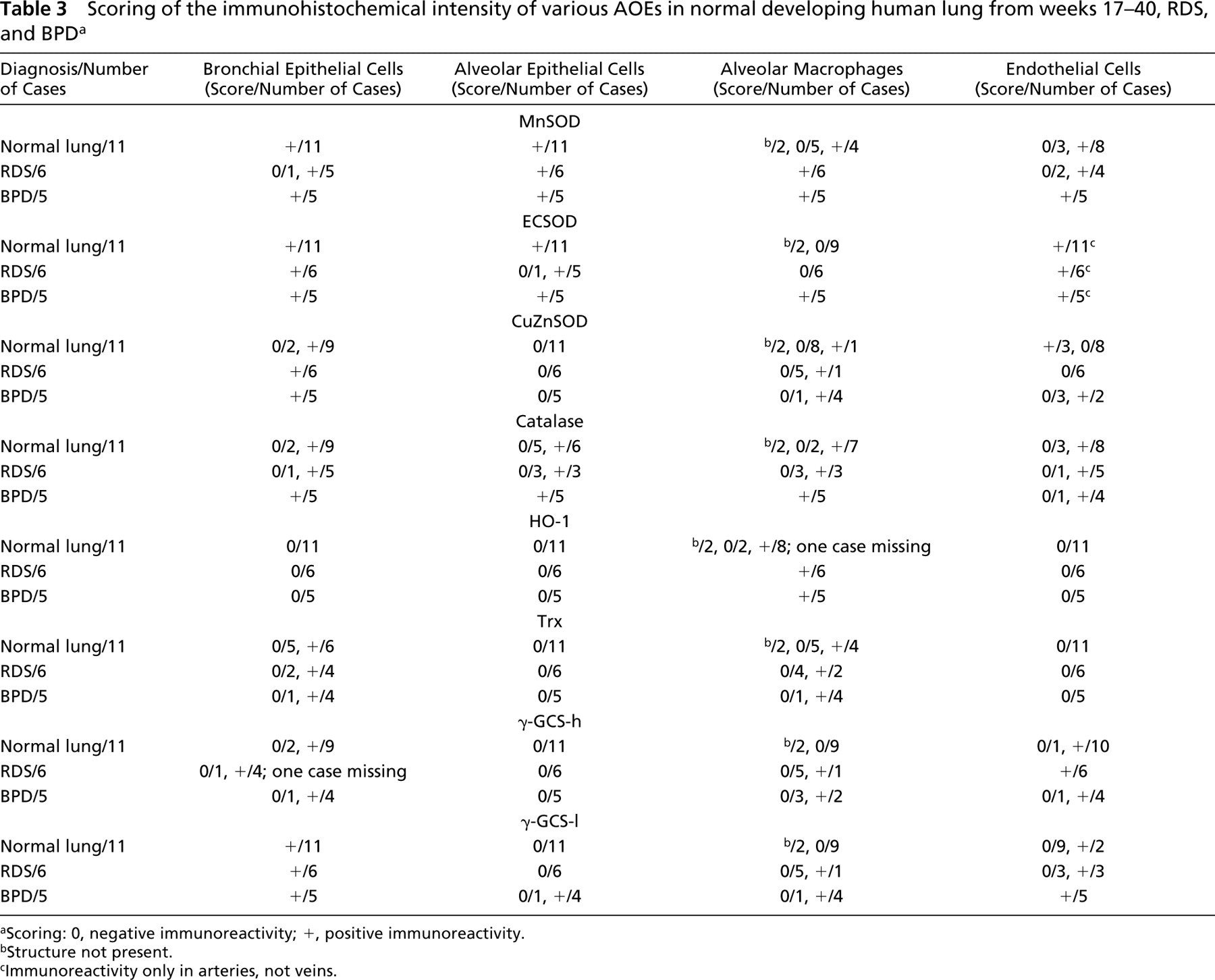
Scoring: 0, negative immunoreactivity; +, positive immunoreactivity.
Structure not present.
Immunoreactivity only in arteries, not veins.
γ-GCS-h
γ-GCS-h was positive in bronchial epithelium, endothelium of arteries and veins, and epithelium of bronchial glands during all developmental periods (Figure 1E). It could also be detected in the same sites in BPD and RDS as in the healthy lung. Chondroid cells showed occasional positive immunoreactivity. Alveolar macrophages were negative in developing normal lung, with the exception of positive expression in two cases of BPD and one case of RDS. Alveolar epithelium, interstitial fibroblasts and myofibroblasts, and mesothelial cells were invariably negative.
γ-GCS-l
γ-GCS-l was positively expressed in bronchial epithelium in every case, including developing normal lung, RDS, and BPD (Figure 1F). In developing normal lung, epithelium of bronchial glands was mainly positive, whereas endothelium, chondroid cells, macrophages, interstitial cells, and alveolar epithelium were negative. These results were similar in RDS with the exception that in two cases the alveolar macrophages were positive. In BPD, metaplastic alveolar epithelium, alveolar macrophages, endothelium, bronchial glandular epithelium, and a few chondroid cells showed positive immunoreactivity.
Trx
Trx was positive in the bronchial epithelium, alveolar macrophages, chondroid cells, and cells of bronchial gland epithelium from week 35 onward, with occasional positivity detected also in the nucleus of bronchial epithelial cells (Figure 1G). Bronchial epithelium of RDS and BPD were also positive. Alveolar epithelium, interstitial fibroblasts and myofibroblasts, and pleural mesothelium were negative.
HO-1
Alveolar macrophages of normal developing lung, RDS, and BPD showed similar HO-1 immunoreactivity (Figure 1H). No immunoreactivity was observed in any other cell type of the lung.
Summary of the expression of various AOEs and related proteins in the neonatal lung

Localization of AOEs at different stages of lung development
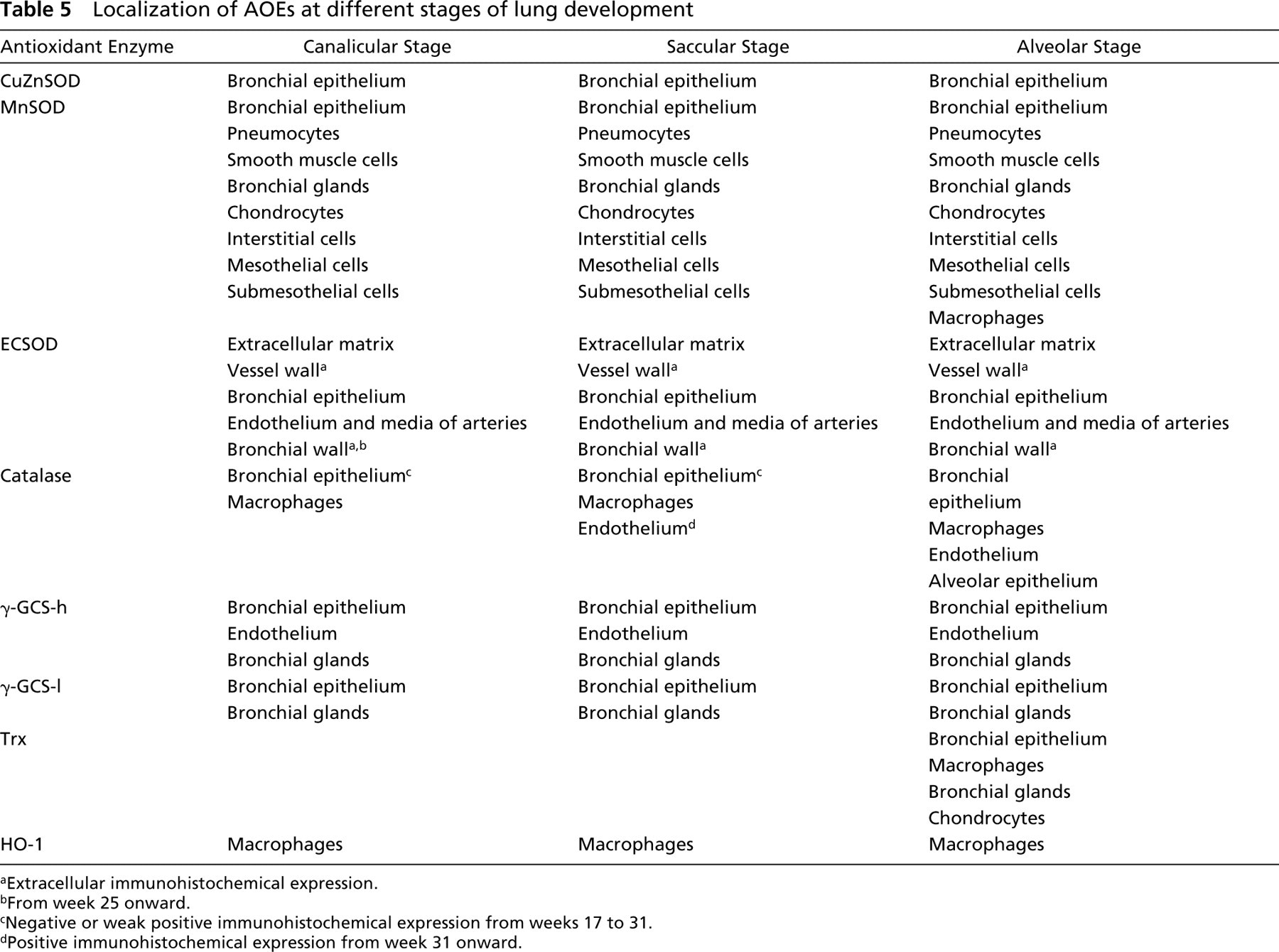
Extracellular immunohistochemical expression.
From week 25 onward.
Negative or weak positive immunohistochemical expression from weeks 17 to 31.
Positive immunohistochemical expression from week 31 onward.
Discussion
We describe here for the first time the distribution and expression of all major AOEs during normal human lung development, RDS, and BPD. This study shows that some enzymes, such as SODs, are expressed already in the fetal lung at the 17th gestational week, whereas others, such as catalase and Trx, appear during the last 4 weeks of pregnancy. These enzymes are mainly concentrated into bronchial epithelium but are also expressed to a variable degree in the developing alveolar epithelium and alveolar macrophages. The most common lung disorders of newborn infants include RDS and BPD. There was a tendency toward increased CuZnSOD, MnSOD, and γ-GCS-l immunoreactivities in alveolar macrophages and toward ECSOD in the interstitium, arteries, and metaplastic alveolar epithelium in BPD, whereas the immunoreactivities of catalase, Trx, γ-GCS-h, and HO-1 were very similar in the normal and diseased lung.
Given the fact that animal and human lungs exhibit important developmental differences, the results obtained from animal lungs and various AOEs cannot be directly extrapolated to human lungs. Because lung tissue contains multiple cell types, the assessment of total mRNA and/or enzyme activity from the total homogenate may underestimate the enzyme level in individual cells. This may be especially important in those cell types that are directly exposed to environmental high oxygen tension and therefore are critical in the protection of lung tissue against exogenous oxidants. There are individual in situ studies into the mRNA levels of some AOEs in animal and human lung (Clyde et al. 1993; Su et al. 1997; Folz et al. 1999). However, mRNA does not necessarily correlate with the actual amount of enzyme protein (Clerch et al. 1998). Immunohistochemistry with specific antibodies remains one of the best methodologies for the assessment of AOEs in various cell types of healthy and diseased lung. There are very few commercial antibodies available for the detection of AOEs in paraffin-embedded tissues, and most of them are neither specific nor sensitive (Yan et al. 1998) The antibodies used in this study have been previously tested in several laboratories and found to detect these AOEs in human lung with high specificity (Oury et al. 1996; Kinnula et al. 1998; Soini et al. 2001a,b). Human neonatal material, which is difficult to obtain, may undergo some postmortem changes in immunoreactivity. Based on our earlier study of MnSOD levels and activities in human fetal material, significant changes are unlikely to occur if the samples can be collected within the first 2 days postmortem (Asikainen et al. 1998). It has to be noted, however, that immunohistochemical expression of the protein does not necessarily correlate with the active protein in individual cells and that a negative immunohistochemical finding does not exclude the existence of miniscule amounts of the enzyme protein in the cells.
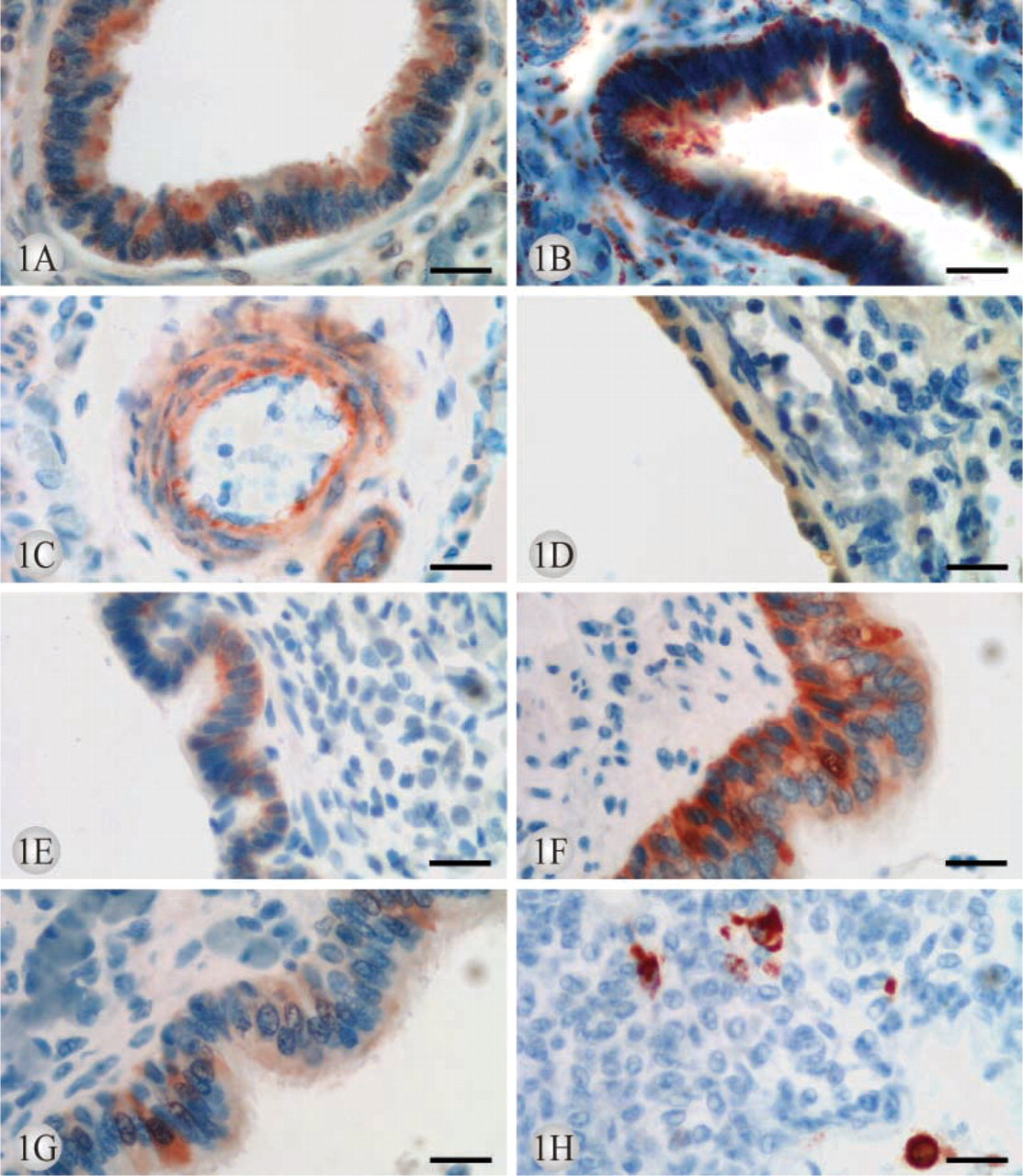
Positive intracellular immunohistochemical expression for CuZnSOD (
MnSOD is one of the most important AOEs in mammalian cells. A total deficiency of MnSOD is lethal (Li et al. 1995; Lebovitz et al. 1996; Tsan 2001), and increased levels of MnSOD protect various cells in several experimental conditions both in vitro and in vivo (St Clair et al. 1991; Wispe et al. 1992; Hirose et al. 1993). MnSOD is induced by cytokines and oxidants that may have important implications in preventing oxidant-related cell injury (Wong et al. 1989; Kinnula and Crapo 2003). In addition to MnSOD, recent knockout studies have emphasized the importance of γ-GCS and Trx for normal development (Matsui et al. 1996; Dalton et al. 2000). CuZnSOD, ECSOD, and catalase knockout animals do survive (Carlsson et al. 1995; Ho et al. 1998), and these enzymes are not inducible to such an extent as MnSOD. However, ECSOD represents the major AOE of the extracellular matrix in the lung (Oury et al. 1996), and catalase is an AOE that consumes H2O2 efficiently in human alveolar macrophages (Pietarinen et al. 1995). Very little is known about HO-1 in the lung, but this enzyme is induced by oxidant stress and probably plays an important role in protecting the lung against oxidants (Choi and Alam 1996). It is not surprising that oxidant protection of lung tissue appears to be associated with all of these proteins (Crapo and Tierney 1974; Crapo et al. 1980; White and Ghezzi 1989; Tsan et al. 1991; Kinnula et al. 1995; Choi and Alam 1996; Das et al. 1999). Simultaneous assessment of these enzymes and their distribution in the lung is the only way to obtain reliable information about the relative importance of the various antioxidant mechanisms in healthy and diseased human lung.
There are few, if any, studies on CuZnSOD during human lung development. These investigations have indicated that CuZnSOD is similar in the prematurely born human infant and adult lungs (Strange et al. 1988,1990; Dobashi et al. 1993). Our recent study suggested that CuZnSOD mRNA increases toward adulthood in human lung but also that the activity of CuZnSOD in the lung homogenates is similar in the fetal, neonatal, and adult lung (Asikainen et al. 1998). This study detected similar CuZnSOD positivity in bronchial epithelial cells after the 17th week of gestation and at term. These results are also consistent with a previous study showing that CuZnSOD is mainly localized to the bronchial epithelium of adult human lung (Lakari et al. 1998).
There are still controversies about the developmental profile of MnSOD in human lungs (Strange et al. 1990; Dobashi et al. 1993; Asikainen et al. 1998,2001). Our study with the AEC method, which is highly sensitive, confirmed the recent results with the diamino benzidine (DAB) method (Asikainen et al. 2001) on the expression of MnSOD already during the 17th gestational week in human bronchial epithelium, developing alveolar epithelium, and alveolar macrophages. In healthy adult human lung, MnSOD is weakly expressed, especially in type II pneumocytes, alveolar macrophages, and bronchiolar epithelial cells (Kinnula et al. 1994; Coursin et al. 1996; Lakari et al. 1998,2000).
There are no earlier studies on ECSOD distribution during the development of human lung. In rabbit lung, ECSOD has been found to be intracellular in both preterm and term lungs, but the extracellular distribution appears to increase toward adulthood (Nozik-Grayck et al. 2000). There are several differences between rabbit and human ECSOD, however, one of those being the lower heparin-binding capacity of rabbit ECSOD. Our results also suggest that the extracellular distribution of ECSOD increases toward adulthood in human lung, because no ECSOD reactivity was observed in the interstitium in newborns, whereas previous studies have reported its expression in the interstitium of adult human lung (Oury et al. 1996)
Catalase was expressed only during the later stages of lung development, which is in agreement with our previous study in which we found catalase was the only AOE that increased at the level of mRNA and specific activity throughout the period of human lung development (Asikainen et al. 1998). This result also indicates that catalase may play an important role against oxidant stress of human lung and that a lack of catalase may also predispose the preterm lung to oxidant-related injury. In agreement, catalase is prominently and constitutively expressed in alveolar pneumocyte II cells and alveolar macrophages of adult lung (Lakari et al. 2000), and it is mainly responsible for the H2O2 consumption in alveolar macrophages subjected to severe oxidant exposure (Pietarinen et al. 1995).
γ-GCS can be considered to be one of the major AOEs in human lung because it is the rate-limiting enzyme in GSH synthesis and GSH has been postulated to be one of the most important antioxidants of human lung (Rahman and MacNee 2000). Importantly, the expression of both subunits of γ-GCS could already be detected at the 17th gestational week, as has also been shown to be the case with GSH peroxidase, a major GSH-related H2O2-scavenging AOE in human lung (Asikainen et al. 1998). The expression of γ-GCS in the bronchial epithelium of developing lung and newborns is consistent with the expression of γ-GCS in the bronchial epithelium of adult human lung (Soini et al. 2001b; Harju et al. 2002a).
Trx was not detectable in the fetal lung at the 17th week of gestation, which suggests that the Trx-system may develop later than the GSH-mediated mechanisms in the lung and play a role especially in the transition from the fetal atmosphere to the ambient oxygen concentrations. This finding is in agreement with recent findings on baboon lung, which have found up-regulation of Trx by high oxygen tension after delivery (Das et al. 1999). The distribution of Trx in newborns was very similar to that observed in adult lung (Soini et al. 2001b; Tiitto et al. 2003).
HO-1 has been considered an efficient antioxidant in the lung. Its distribution has not been investigated in human lung development, but its expression in alveolar macrophages is consistent with recent findings that macrophages are the major cell type to express HO-1 also in adult lung (Lakari et al. 2001; Harju et al. 2002b).
RDS and BPD represent the major lung complications of newborn babies. The evaluation of AOEs in these disorders is difficult because these enzymes can be induced or inactivated by high oxygen tension and cytokines (Wong et al. 1989; Clerch et al. 1998). In addition to oxygen therapy, premature infants are treated with numerous therapeutic regimens that may also have some direct effects on nearly all of these enzymes. The baboon model of BPD has revealed that MnSOD protein expression is increased in BPD, being regulated by posttranscriptional mechanisms (Clerch et al. 1996), but there have been very few investigations on the other AOEs in neonatal lung diseases. The following conclusions can be drawn from these few investigations: the expression of MnSOD is similar in RDS, BPD, age-matched controls, and healthy newborns (Strange et al. 1990; Asikainen et al. 2001), with the exception of higher MnSOD immunoreactivity in the proliferating pneumocyte II cells of BPD (Dobashi et al. 1993) and alveolar macrophages. Our study also suggests that RDS is associated with only minor, if any, induction of AOEs but also that lung tissue has a well-developed antioxidant already several weeks before birth. However, the trend toward increased levels of several AOEs in BPD, especially in alveolar macrophages, emphasizes the importance of these cells in the defense of human lung. The expression of ECSOD in the alveolar interstitium in BPD may reflect an attempt to protect lung tissue in parenchymal lung disorders of newborns. It can be concluded that various AOEs differ in their distribution and expression before term, but in general, infants are better adapted to high oxygen tension than might be expected.
It seems that the risk of oxidant stress in RDS/BPD is increased by several mechanisms, including increased oxygen concentration in the inspired air and inflammation and consequent activation by numerous oxidant-producing pathways. The major determinant in the development of oxidant-mediated lung injury may ultimately be attributable to the relatively modest induction of the antioxidant defense occurring in these diseases. These suggestions are in full agreement with the observed attenuation of lung injury in an experimental BPD model with a SOD mimetic (Chang et al. 2003). In agreement with these proposals, our unpublished studies show that RDS lungs are associated with high neutrophil myeloperoxidase activity, one indicator of highly increased oxidant generation. Overall, even though the antioxidant defense in newborn lungs is well developed, its response to oxidants via induction and its supplementation with new potential anti-oxidants await future investigations.
Footnotes
Acknowledgements
Supported by the Finnish Anti-Tuberculosis Association Foundation, the Juselius Foundation, the Paulo Foundation, and the Research Foundation of Orion Corporation.
MD, PhD Riitta Herva is kindly acknowledged for her expert knowledge of pediatric pathology. The technical assistance of Manu Tuovinen and Hannu Wäänänen is kindly acknowledged.