
Abstract
Our objective in this study was to localize the corticotropin-releasing factor 2 receptor (CRF2R) in rodent and human skeletal muscle. We found CRF2R protein to be abundant in neural tissues in skeletal muscle, including large nerve fibers and bundles, neural tissue associated with mechanoreceptors, muscle spindles, and the Golgi tendon organ. CRF2R protein was also abundant in blood vessels in skeletal muscle. CRF2R protein was also observed, although with less abundance, in the endo/perimysial regions in skeletal muscle. The localization of the CRF2R to blood vessels is consistent with the CRF2R-mediated vascular phenomena observed previously, but the observation of CRF2R in neural tissue in skeletal muscle is a novel finding with an unknown function. (
T
Recently we have discovered that activation of the CRF2R modulates skeletal muscle mass under physiological and pathological conditions (Hinkle et al. 2003). We have also demonstrated that the CRF2R protein expressed in skeletal muscle is active and positively coupled to Gαs (Hinkle et al. 2003). Therefore, the CRF2Rs expressed in skeletal muscle are biologically active and appear to have a role in the maintenance of skeletal muscle mass. Although CRF2R protein localization has been demonstrated in several tissues, the location of the CRF2R in skeletal muscle has not been investigated. We have undertaken experiments designed to localize CRF2R expression in skeletal muscle.
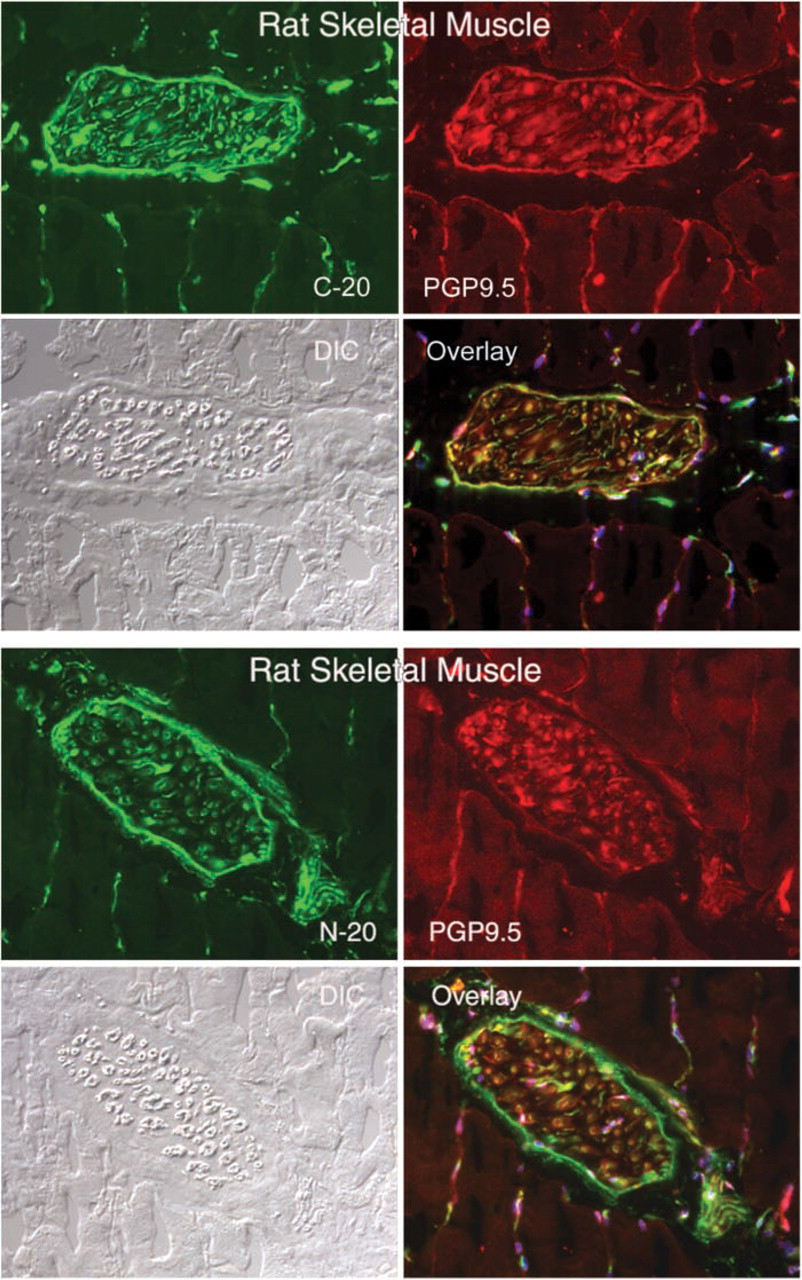
Immunofluorescence micrographs showing the co-localization of CRFR antibodies C-20 (CRFR-specific) and N-20 (CRF2R-specific) with the neural marker PGP9.5 in large neural structures of rat skeletal muscle. Magnification ×20.
Materials and Methods
Six-week-old male and female Sprague-Dawley rats (Charles River; Raleigh, NC) and C57Bl6 mice (Charles River) were housed singly and acclimatized to the conditions of the facility for ∼1 week before use. Mice and rats had access to lab chow and water ad libitum. Animals were subjected to standard conditions of humidity, temperature, and a 12-hr light cycle. All studies described in this report were conducted in compliance with the US Animal Welfare Act and rules and regulations of the State of Ohio Departments of Health, and were approved by the local IUCAC. Rat and mouse lower leg muscles (tibialis anterior and medial gastrocnemius muscles) were obtained by removing the indicated muscles and immediately placing the muscles in optimal cutting temperature (OCT) compound (Sakura Finetek; Torrance, CA). Tissues in OCT compound were frozen in liquid nitrogencooled isopentane and held at −70C. Human vastus lateralis muscle biopsies from healthy young adults, frozen in OCT, were obtained from Dr. Mark Tarnopolsky (McMaster University; Hamilton, Ontario, Canada). Human muscle biopsies were obtained under the approval from the McMaster University Ethics Committee. Two anti-CRF2R antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, CA) and were used throughout this study. The CRFR (C-20, sc-1757) antibody reacts with both CRFR1 and CRFR2, while the CRFR2 (N-20, sc-1826) antibody is specific for CRFR2. Anti-CRFR1 and -CRFR2 (RDI-CRFR1CabG, RDI-RTCRFR2NabG) antibodies (Research Diagnostics; Flanders, NJ) were used to validate results obtained with the Santa Cruz products. The specificity of each CRFR antibody was established by running preadsorption controls. Each antibody was preincubated for 4 hr with a 5000-fold molar excess of its corresponding peptide (sc-1757P for C20 and sc-1826P for N20) or an irrelevant peptide. The antigen-antibody mixture was centrifuged at 100,000 × g for 30 min with the supernatant used in place of primary antibody in the normal immunocytochemistry protocol. Preadsorbed samples were processed in parallel with normal CRFR antiserum to optimize comparative staining.
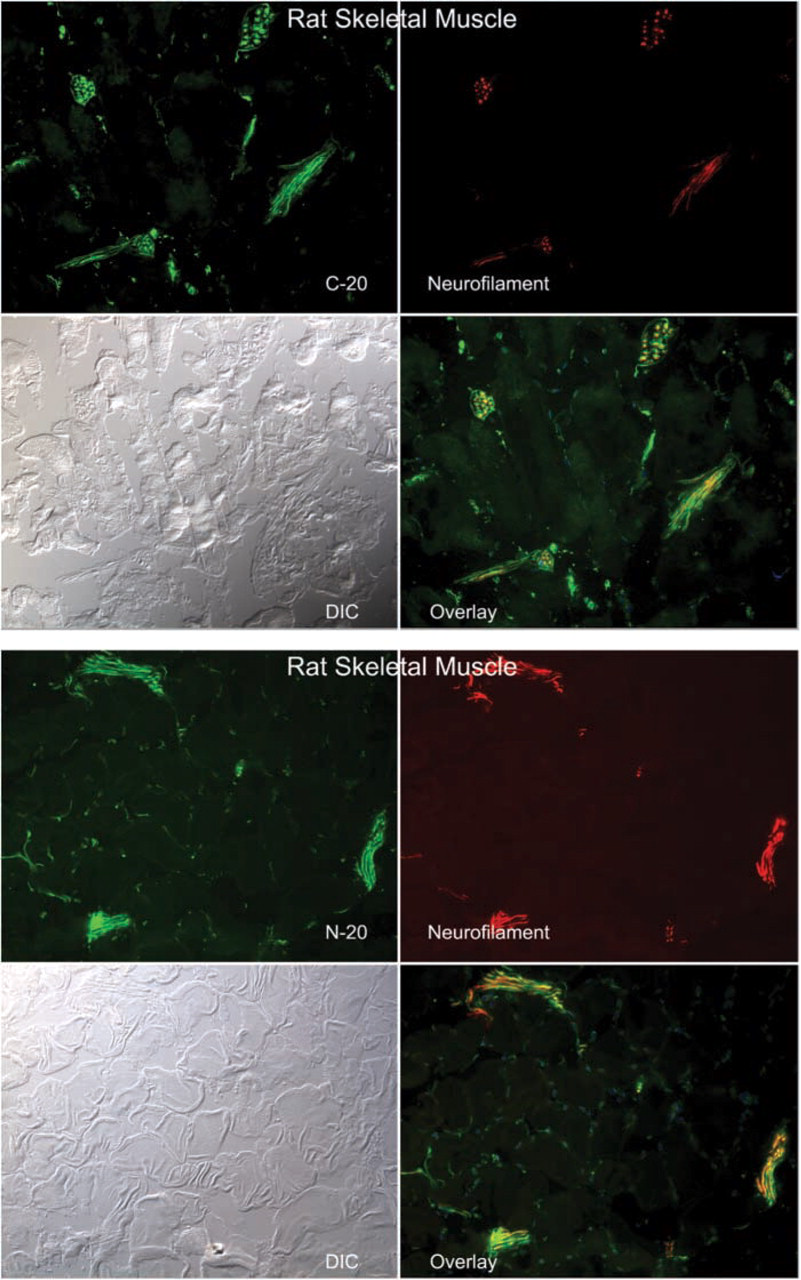
Immunofluorescence micrographs showing the co-localization of CRFR antibodies C-20 and N-20 with the neural marker neurofilament in large neural structures of rat skeletal muscle. Magnification ×20.
Immunocytochemical analysis was performed on 7-μm frozen sections by indirect labeling, as follows. Cryostat sections were attached, the slides were warmed to room temperature, sections were encircled with a PAP pen, and were hydrated in PBS. Sections were permeabilized in PBS/0.1% Triton X-100 for 30 min, aspirated, and covered with blocking buffer of 10% normal donkey serum (NDS) in BSA buffer [2% bovine serum albumin (BSA)/0.1 M phosphate buffer, pH 7.4] for 1 hr at room temperature (RT). Primary antibodies were diluted in BSA buffer, and sections were incubated directly, after aspiration of blocking solution, for 1 hr at RT. Slides were washed three times for 10 min in PBS, then incubated with fluorescein-conjugated donkey secondary antibodies (Jackson Immunoresearch Labs; West Grove, PA) at 5 μg/ml in BSA buffer with 0.5 μM diamidinophenylindole dihydrochloride (DAPI; Molecular Probes, Eugene, OR) for 1 hr at RT. Sections were washed twice for 10 min in PBS with a final wash in 0.1 M Tris buffer, pH 8.5. Sections were coverslipped under PPD glycerol (10% 1 M Tris, 90% glycerol, pH 8.5, containing 1 mg/ml p-phenylene-diamine), sealed with nailpolish, and stored at −20C. Tissue sections were examined with a Nikon Microphot FXA (Melville, NY). Images were captured with a Spot II digital camera (Diagnostic Instruments; Sterling Heights, MI) and processed using Metamorph 4.6 imaging software (Universal Imaging; Downingtown, PA).
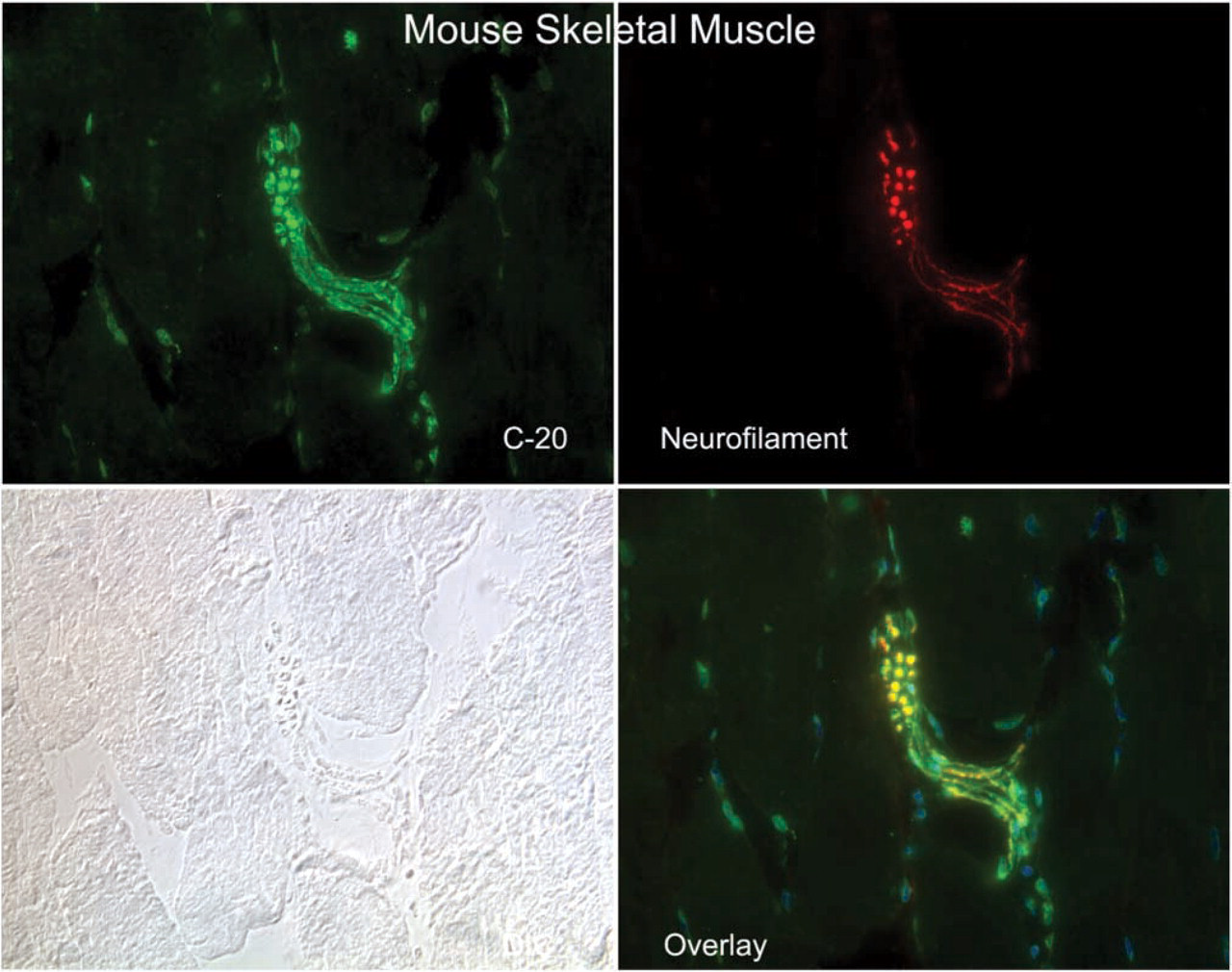
Immunofluorescence micrographs showing the co-localization of the CRFR antibody C-20 with the neuronal marker neurofilament in a large neural structure of mouse skeletal muscle. Magnification ×40.
Co-localization analyses were performed with the following landmark antibodies and filter sets appropriate for fluorescein and Cy-3: α-smooth muscle actin (A2547; Sigma Chemical, St Louis, MO); β-tubulin (T4026; Sigma Chemical; neurofilament (34–1000; Zymed Laboratories, San Francisco, CA); PGP9.5 (UC RA 95101; Accurate Chemical and Scientific, Westbury, NY); nestin (rat-401; Developmental Studies Hybridoma Bank, Iowa City, IA); and rhodamine-labeled α-bungarotoxin (B13421; Molecular Probes).
Results
CRF2R localization in rat, mouse and human skeletal muscle was performed using two antibodies, one that was CRFR-specific with equivalent affinity for both CRF1R and CRF2R (C20) and a second that was specific for CRF2R only. Staining was considered positive if both antibodies stained the same structures. Both of these antibodies were crossreactive with mouse, rat, and human CRF2R, allowing cross-species comparison. In addition, we attempted to localize the CRF2R to specific substructures in skeletal muscle using antibodies specific for proteins found in neurons (neurofilament and PGP9.5) (Krikler et al. 1992; Nahirney and Ovalle 1993; Blumer et al. 2001; Kawabuchi et al. 2001), proteins found on blood vessel (α-smooth muscle actin) (Nora et al. 1998), and proteins found at the myotendinous junction (tubulin and nestin) (Ralphs et al. 1991; Yarnitzky et al. 1997; Carlsson et al. 1999; Vaittinen et al. 1999). In addition, we used rhodamine-conjugated α-bungarotoxin (Molecular Probes) to co-localize CRF2R staining to the neuromuscular junction. With these tools, we observed CRF2R protein staining in large nerve bundles and intrafusal fibers of muscle, where it co-localized with neuronal markers PGP9.5 and neurofilament protein staining (Figures 1–3). At neuromuscular junctions, CRF2R staining co-localized with bungarotoxin (Figure 4). Rhodamine-conjugated α-bungarotoxin was used to delineate the postsynaptic membrane of the neuromuscular junction, where it binds tightly to the acetylcholine receptors (Blumer et al. 2001; Kawabuchi et al. 2001).
CRF2R protein staining was also observed at the myotendinous junction, where it is co-expressed with nestin and tubulin (Figure 5). CRF2R protein staining was enriched on blood vessels; CRF2R staining co-localized with smooth muscle actin staining (Figure 6). Finally, CRF2R staining was found in the endo- and perimysium surrounding rat, mouse, and human muscle fibers (Figure 7). These connective tissue regions contain small arterioles, capillaries, venules, fibroblasts, neural processes, and satellite cells. CRF2R staining in all the human skeletal muscle samples was similar (five males and five females). Because of sampling limitations for human skeletal muscle, we were unable to evaluate CRF2R staining at myotendinous junction, neuronal, and vascular regions (samples were needle biopsies taken from the belly of the vastus lateralis muscle). Finally, C20 and N20 antibody specificity was evaluated by blocking antibody binding with peptide immunogen compared with staining in which the antibody was blocked with an irrelevant peptide. The blocking data were used in conjunction with our control data to establish staining and antibody specificity (Figure 8).
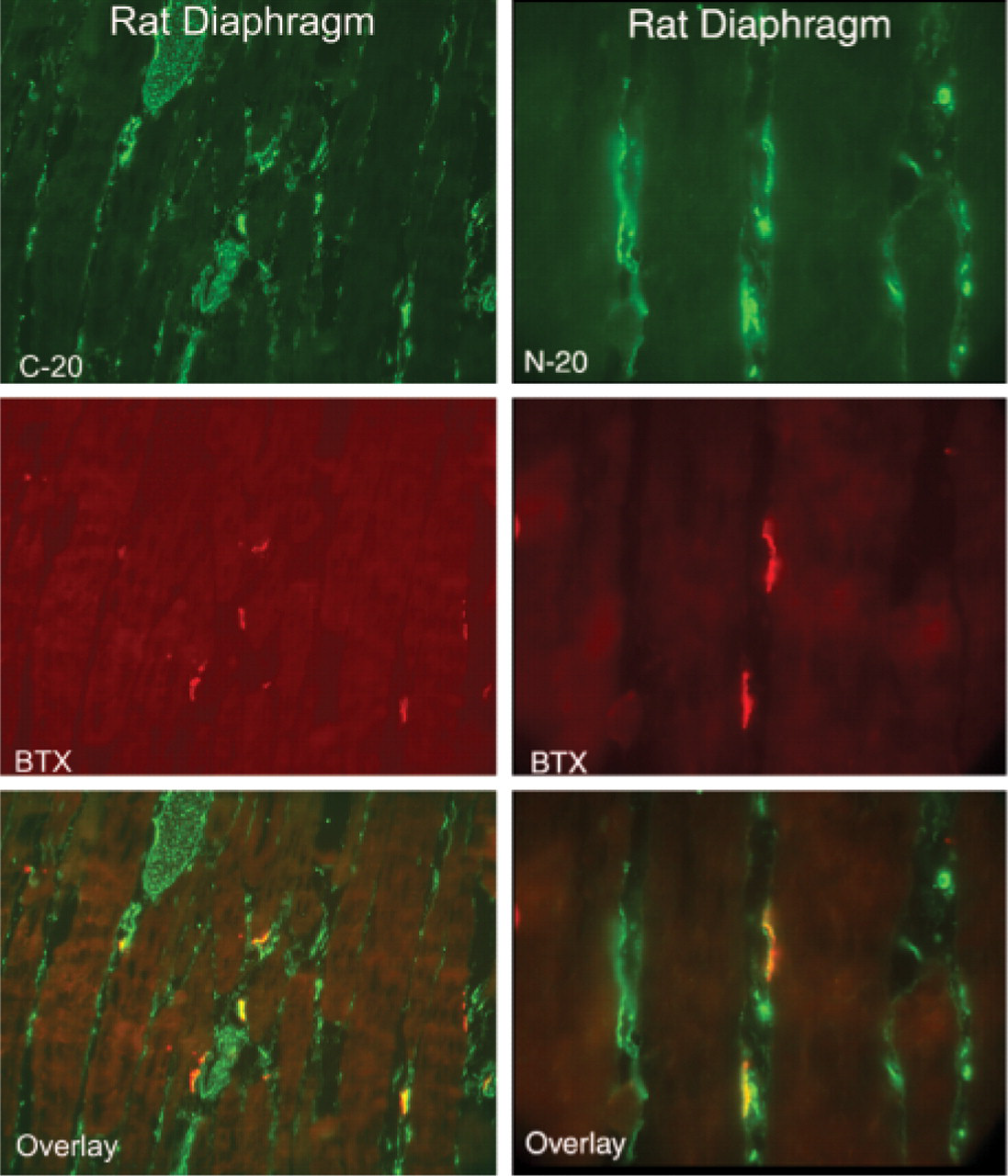
Longitudinal sections of rat diaphragm muscle showing co-localization of CRFR antibodies C-20 and N-20 with α-bungarotoxin (BTX). Magnification ×20.
Discussion
In this study we showed that the CRF2R in skeletal muscle localized to neural structures, blood vessels, myotendinous junctions, and endomysial/perimysial spaces. Interestingly, expression of the CRF2R in skeletal muscle myocytes was not observed. This raises the possibility that the effects of CRF2R activation on skeletal muscle mass may be an indirect effect, potentially secondary to activation of the CRF2R on either nerves, blood vessels, myotendinous junction, or endo/perimysium. Current understanding of CRF2R effects on skeletal muscle does not provide conclusive evidence about which of these sites of CRF2R expression is critical for the skeletal muscle mass effects of CRF2R activation. However, it can be inferred from the data presented here that the effects of CRF2R activation on skeletal muscle mass are an indirect myocyte effect. One possibility that is consistent with known CRFR biology in other tissues is that activation of the CRF2R results in the release of trophic factors from nerve endings, factors that directly increase skeletal muscle mass. In particular, it is known that members of the CRH family of ligands function as neurohumoral factors (Garzon et al. 1985; McAlexander and Undem 1997; Heymann-Monnikes et al. 1991; Tamaoki et al. 1998; McCarthy et al. 1999; Grill et al. 2000; Wang et al. 2000; Kihara et al. 2001; Dautzenberg and Hauger 2002; Miampamba et al. 2002). It has been observed that peripheral CRFR agonistic peptides activate CRFR on neurons in the colon, bronchi, stomach, airway smooth muscle, ileum, and duodenum (Garzon et al. 1985; Heymann-Monnikes et al. 1991; McAlexander and Undem 1997; Tamaoki et al. 1998; McCarthy et al. 1999; Grill et al. 2000; Wang et al. 2000; Kihara et al. 2001; Dautzenberg and Hauger 2002; Miampamba et al. 2002). The effect of CRFR activation in these tissues is directly mediated either by stimulation of the CRFR on the nerves that innervate that tissue or by potentiating the action of other neurohumoral factors. In addition, it is known that neural release of trophic factors is an important mechanism for the maintenance of skeletal muscle mass. Therefore, the indirect release of these factors via CRF2R activation is a viable mechanism of CRF2R action for mediating myocyte mass (Bray and Harris 1975; Markelonis et al. 1980; Davis 1983; Helgren et al. 1994; Fernandez et al. 1999; Marques and Neto 1997. Alternatively, activation of the CRF2R expressed at the myotendinous junction may provide a feedback loop connected with muscle use, resulting in increased muscle mass through a neuronally mediated mechanism, possibly through the release of trophic factors. Localization of the CRF2R at the myotendinous junction and co-localizing with nestin indicates that the CRF2R may be part of the Golgi tendon organ, a nerve rich portion of the tendon involved in mechanosensation (Swett and Schoultz 1975). In addition, CRF2R associated with free nerve endings found throughout skeletal muscle and with muscle spindles may also regulate the release of trophic factors associated with increased muscle use because both of these neural structures are involved in mechanosensation (Stacey 1969; Mense and Meyer 1985; Hunt 1990; Nahirney and Ovalle 1993).
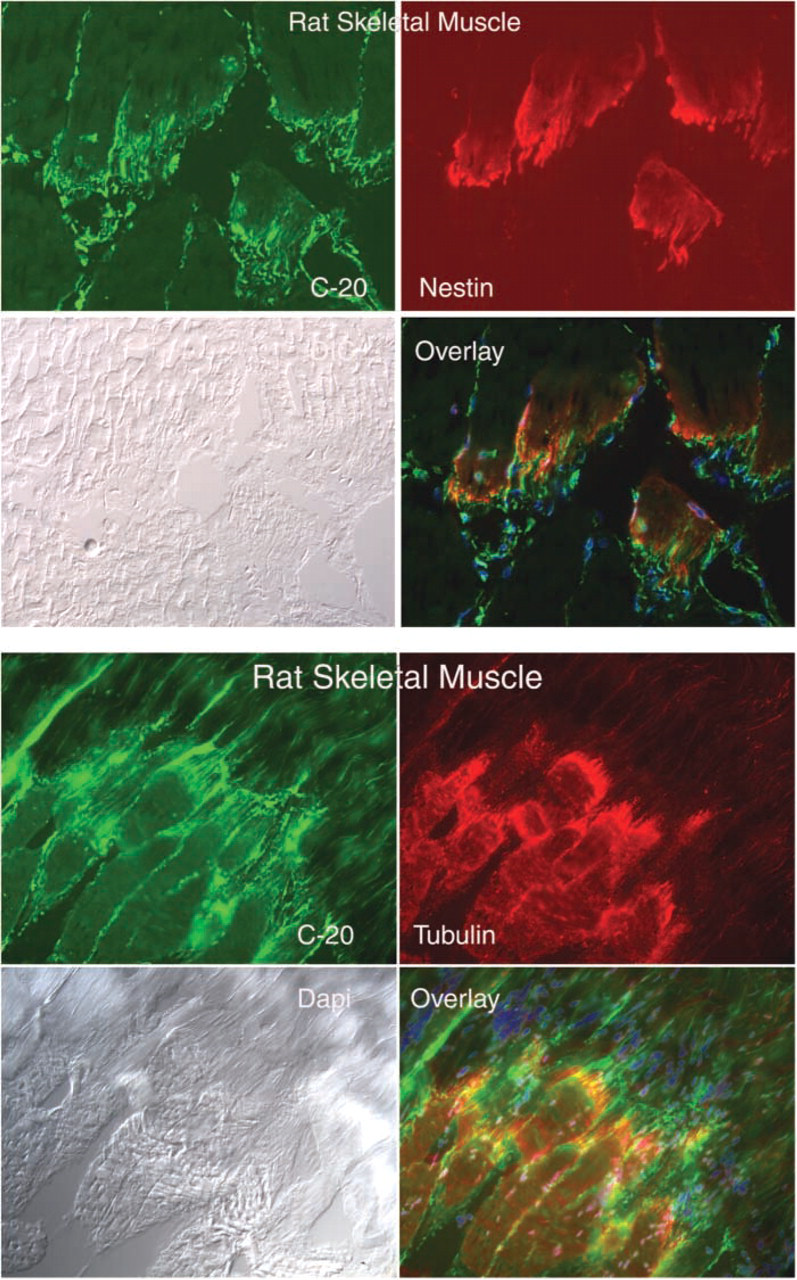
Longitudinal sections through the transition region between rat skeletal muscle fibers and their associated tendon (myotendinous junction), showing co-localization of CRFR antibody C-20 with nestin and tubulin. Magnification ×20.
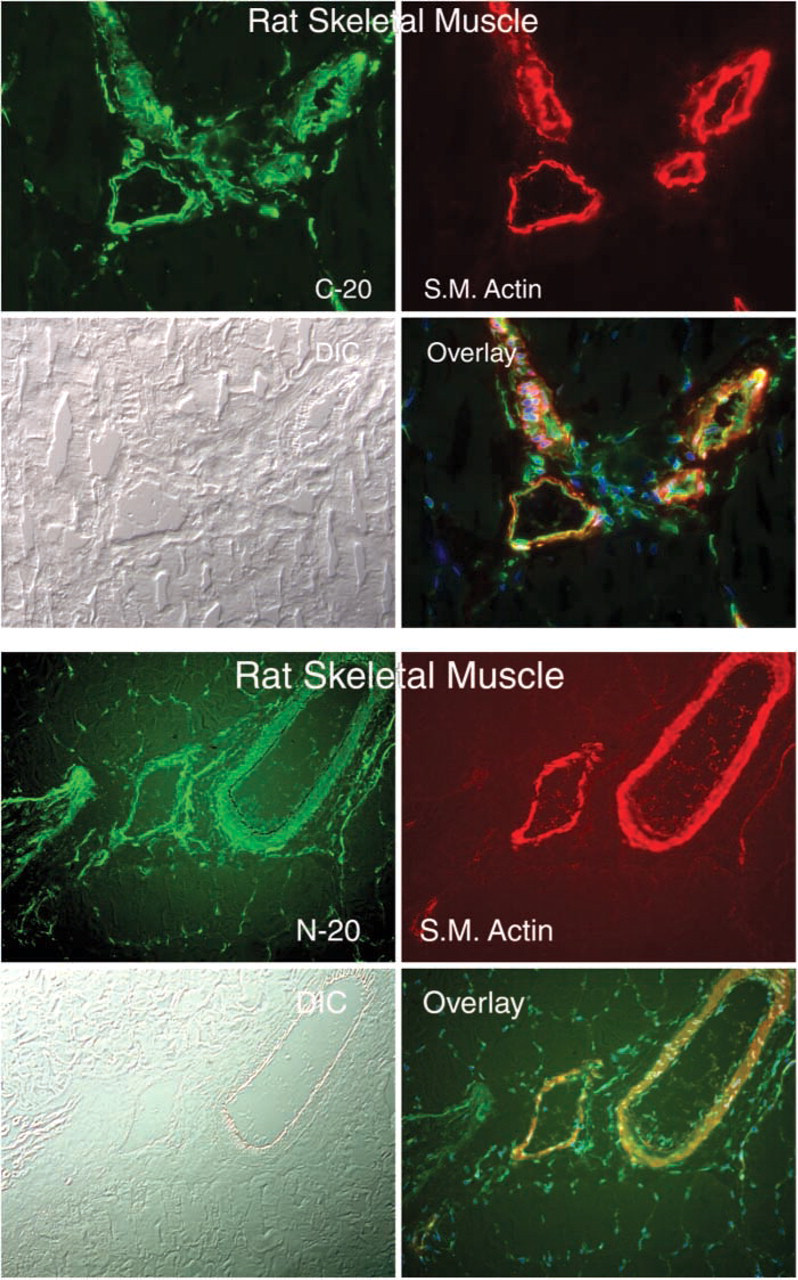
Immunofluorescence micrographs showing the co-localization of CRFR antibodies C-20 and N-20 with smooth muscle actin (S.M. Actin) in blood vessels of rat skeletal muscle. Magnification ×20.
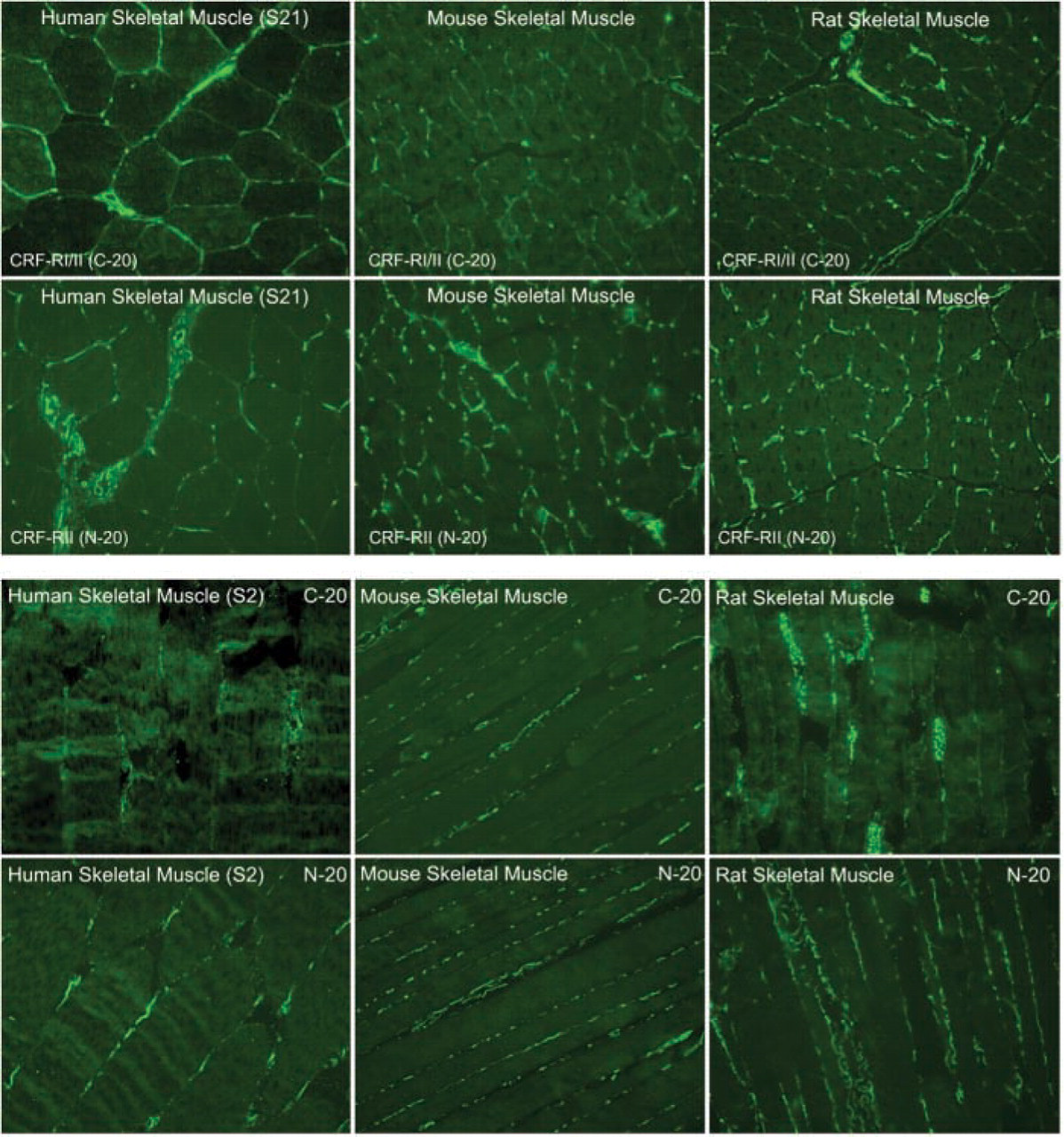
Immunofluorescence micrographs of cross- and longitudinal sections through human (S21 and S2), mouse and rat skeletal muscle demonstrating localization of the CRFR antibodies with C-20 and N-20. Magnification ×20.
Do CRF2Rs present on blood vessels or in the endo/perimysium function to modulate skeletal muscle mass? CRF2Rs present on blood vessels are known to have an important role in the control of blood pressure and vasodilatation (Coste et al. 2000). It is unclear how vasodilatation would function to regulate skeletal muscle mass other than to release fluid content in muscle. We have previously demonstrated that the increase in muscle mass observed after CRF2R activation is mediated by increased myocyte mass and by myocyte cross-sectional and myocyte force production (Hinkle et al. 2003). Therefore, CRF2R activation does more than increase fluid content in muscle. It is possible that vasodilatation leads to increased exposure of myocytes to serum anabolic growth factors, such as IGF-1, a known skeletal muscle anabolic hormone. CRF2R present in the endo/perimysium may have a role in regulating myocyte mass and contractile apparatus protein levels by the production, by fibroblasts or other connective tissue cells present in the endo/perimysium, of anabolic growth factors. This hypothesis does have some support from previously reported experimental evidence demonstrating that activation of CRFR in fibroblast modulates prostaglandin synthesis (Fleisher-Berkovich and Danon 1995). The results we report here are not sufficient to validate any of these possible mechanisms. However, the data we present are important because they provide a set of observations that provide insights necessary for the design of future experiments to evaluate the validity of these potential mechanisms of CRF2R action.
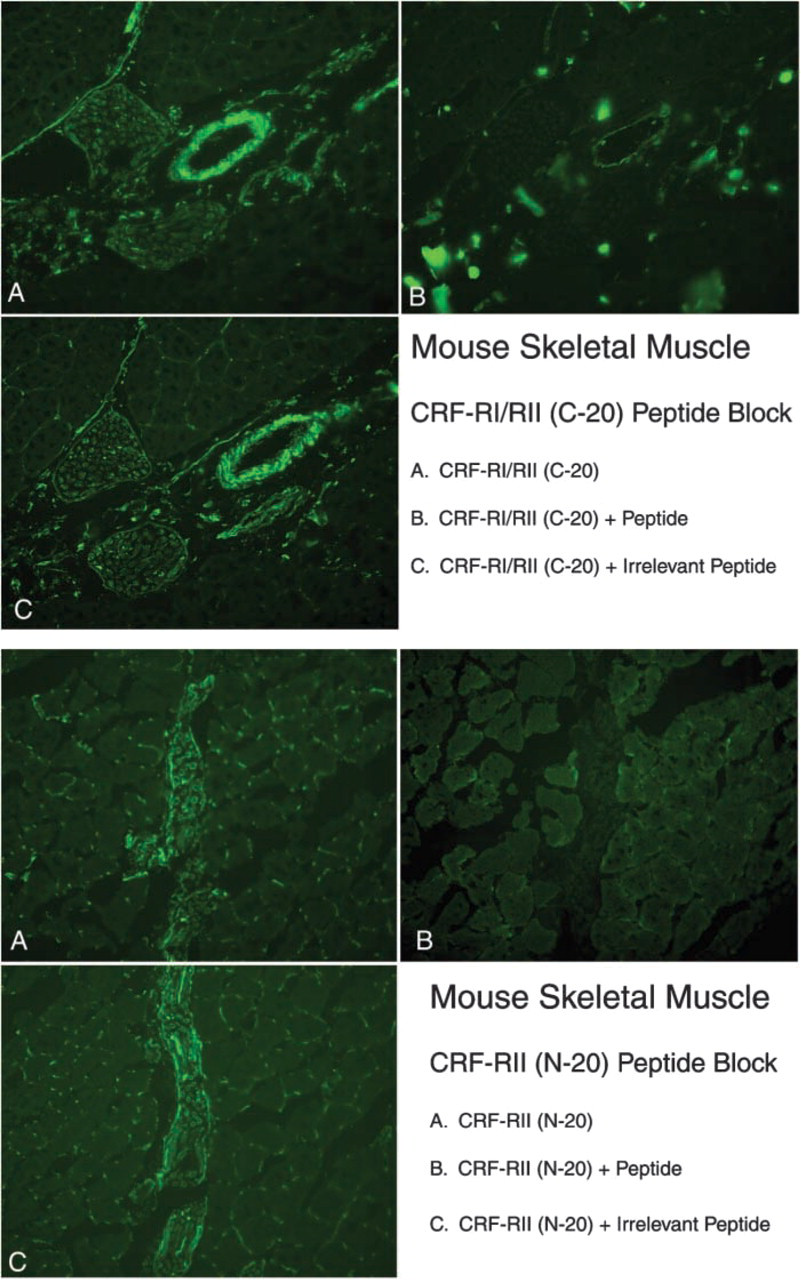
CRFR antibodies C-20 and N-20 staining specificity in mouse skeletal muscle. Immunoreactivity was blocked with the peptide immunogen but was not blocked with an irrelevant peptide. Magnification ×20.
In conclusion, CRF2R protein has been localized to neural structures, myotendinous junction, blood vessels, and endo/perimysial space in skeletal muscle. CRF2R expression was not observed on myocytes, even though treatment of skeletal muscle in vivo resulted in increased myocyte mass. The mechanism underlying the effects of CRF2R activation on regulating skeletal muscle mass is presently unknown.