
Abstract
In murine skin, after depilation-induced anagen, there was a differential spatial and temporal expression of pro-opiomelanocortin (POMC) mRNA, of the POMC-derived peptides β-endorphin, ACTH, β-MSH, and α-MSH, and of the prohormone convertases PC1 and PC2 in epidermal and hair follicle keratinocytes and in the cells of sebaceous units. Using a combination of in situ hybridization histochemistry and immunohistochemistry, we found cell-specific variations in the expression of POMC mRNA that were consistent with immunoreactivities for POMC-derived peptides. Cells that contained POMC peptide immunoreactivity (IR) also expressed POMC mRNA, and where the IR increased there was a parallel increase in mRNA. The levels of PC1-IR and PC2-IR also showed cell-specific variations and were present in the same cells that contained the POMC peptides. Based on the cleavage specificities of these convertases and on the spatial and temporal expression of the convertases and of ACTH, β-endorphin, β-MSH, and α-MSH, we can infer that the activities of PC1 and PC2 are responsible for the cell-specific differential processing of POMC in murine skin.
P
In the pituitary and hypothalamus, posttranslational proteolysis of POMC is performed by the prohormone convertases PC1 and PC2. Cleavage by PC1 results primarily in the production of ACTH and β-lipotropin and, to a minor extent, β-endorphin, whereas cleavage by PC2 elicits β-endorphin and N-terminally extended corticotropin containing the joining peptide and either α-MSH or desacetylated-α-MSH. Removal of the terminal dipeptide from β-endorphin-[1–31] to yield β-endorphin-[1–27] is mediated solely by PC2 (Seidah et al. 1993).
To directly study the relationship between POMC gene transcription and the production of POMC peptides after depilation-induced anagen in murine skin, we used in situ hybridization (ISH) histochemistry to identify sites of POMC mRNA expression and immunohistochemistry (IHC) to identify the cells that contained POMC peptides (β-endorphin, ACTH, β-MSH, and α-MSH), and the prohormone convertases PC1 and PC2.
All of the analyses in the present study were performed on tissue sections that were taken from the same skin samples at each time point, thus permitting direct correlations of expression and processing of the POMC precursor. The results reported here can also be directly correlated with an earlier biochemical and molecular biology study in which we had demonstrated by RT-PCR that POMC mRNA is expressed constitutively in murine skin (Slominski et al. 1996). In addition, we have also shown that POMC mRNA is upregulated during anagen development and that this is accompanied by increased production of β-endorphin and ACTH (Slominski et al. 1992, 1996, 1998; Furkert et al. 1997).
Materials and Methods
Tissue Preparation
All animal experiments were performed in compliance with guidelines of the National Institutes of Health for the use of laboratory animals. C57 BL/6 mice (female, syngenic, 6–8 weeks old) were purchased from Charles River (Kingston, NY) housed with 12-hr light/darkness periods, and fed with water and mouse chow ad libitum. Anagen was induced in anesthetized mice (30 mg/kg sodium pentobarbital) by hair depilation using a warm beeswax/rosin mixture as described previously (Slominski et al. 1992, 1996). Mice with hair follicles at telogen (Day 0) and at Days 3, 5, 6, 9, 10, and 15 after depilation were sacrificed by cervical dislocation. Skin was dissected at the level of the subcutis and representative samples were fixed in 4% buffered formalin. Telogen mice were shaved before harvesting of the skin. Tissue samples were then embedded in paraffin.
Immunohistochemistry
Sections were deparaffinized with xylene and rehydrated through a decreasing ethanol series to phosphate-buffered saline (PBS; 150 mM NaCl/10 mM sodium phosphate, pH 7.3). Endogenous peroxidase activity was blocked by incubation with 1% H2O2 in absolute methanol for 5 min. Nonspecific protein binding was blocked with a solution of 5% nonfat dry milk, 0.1% Triton X-100 in PBS (Blotto) for 5 min. Samples were then washed in PBS three times for 5 min each.
Immunoperoxidase Detection
POMC antigens were detected by the indirect avidin–biotin–peroxidase method (ABC standard; Vector Laboratories, Burlingame, CA). Color was developed with 3, 3′ diaminobenzidine. Primary antisera were diluted in blocking buffer, applied to tissue sections that were encircled with a PAP pen (Research Products International; Mount Prospect, IL), and incubated for 90 min at room temperature (RT). Rabbit anti-ACTH (1:600) and rabbit anti-β-endorphin (1:500) were gifts from Dr. R.G. Allen (Oregon Health Sciences University); rabbit anti β-MSH (1:500) was a gift of Dr. S.H. Pomerantz (University of Maryland). Optimal dilutions and incubation times had been determined in a previous study (Slominski et al. 1993). The crossreactivity of the antibodies aganist β-endorphin and ACTH with non-corresponding POMC peptides has been reported to be <1%, although they recognize the POMC precursor (Allen et al. 1978, 1980). The antibodies against β-MSH had slightly lower specificity, crossreacting with α-MSH (20%) and with ACTH (5%) (Ances and Pomerantz 1974). Rabbit anti-α MSH (1:600) was from Chemicon International (Temecula, CA). According to the manufacturer, there is no crossreactivity of this antiserum with other POMC peptides. Controls (not shown) consisted of tissue incubated with nonimmune sera or with primary antibody that had been incubated with 0.1 mmol/liter concentrations of β-endorphin, ACTH (Peninsula Laboratories; Belmont, CA), β-MSH (gift of Dr. G. Davila, Yale University), or a mixture of porcine pituitary POMC peptides (gift of Dr. J. Pawelek, Yale University). In all cases at the dilutions used, the immunoreactivty was abolished when the primary antiserum was absorbed with its respective peptide. If significant amounts of unprocessed POMC precursor had been present in the tissue sections, blocking with the specific peptide alone would not have eliminated crossreactivity. Some sections were counterstained with Gill's hematoxylin. Photographs were taken on an Olympus BX40 upright microscope interfaced with a SONY Mavigraph video printer. Photographs were digitized and the figures were assembled and labeled in Photoshop.
Immunofluorescence Detection
The prohormone convertases PC1 and PC2 were detected by indirect immunofluorescence and confocal laser scanning microscopy (CLSM). Rabbit anti-PC1-Ab888 and anti-PC2-Ab1159 (1:100) were gifts from Dr. B.A. Eipper (Johns Hopkins University School of Medicine). The secondary antibody was a goat anti-mouse Ig–Alexa488 (Molecular Probes; Eugene, OR). Specimens were analyzed on a NORAN-OZ CLSM interfaced with a Nikon Diaphot 200 inverted microscope equipped with a PlanApo ×60, 1.4 NA oil-immersion objective lens. The specimens were first examined by conventional widefield fluorescence microscopy to assess the specificity of the immunostaining procedure and to identify samples with the highest fluorescence intensity. The instrument settings on the CLSM for brightness, contrast, laser power, and slit size were optimized for the brightest sample to ensure that the CLSM was set for its full dynamic range and that no images were saturated. These settings were used for all subsequent image collection for a particular immunostaining experiment. This permits relative quantitation of the cellular content of these antigens for comparative purposes, and any differences can be readily discerned directly from the images.
In Situ Hybridization Histochemical Localization
Skin sections from telogen, and at 3, 5, and 10 days post anagen induction were subjected to ISH. POMC mRNA was localized in skin and pituitary sections according to procedures outlined in Simmons et al. (1989) modified for the use of digoxigenin (DIG)-labeled cRNA using principles described in Panoskaltsis–Mortari and Bucy (1995). Sense and antisense probes were synthesized from a 700-bp plasmid containing the entire POMC exon 3, a gift from Dr. J. Roberts (Mt. Sinai Medical Center, New York) (Whitfeld et al. 1982) using a Pω Riboprobe Gemini Transcription System (Promega; Madison, WI) and Boehringer–Mannheim DIG-labeling reagents (Boehringer–Mannheim; Indianapolis, IN).
Tissue sections (5 μm) mounted on Superfrost/Plus microscope slides (Fisher Scientific; Pittsburgh, PA) were deparaffinized in xylene and rehydrated in an ethanol series to DEPC (diethypyrocarbonate)-treated H2O. Sections were transferred to 0.1 M Tris-HCl, 50 mM EDTA, pH 8.0, pre-warmed to 37C. Proteinase K from a stock solution in the same buffer was added to a final concentration of 0.001% and incubated for 30 min at 37C. Sections were then washed briefly (∼1 min) in DEPC-treated H2O, then in freshly prepared 0.1 M triethanolamine buffer, pH 8.0, and acetylated in 0.25% acetic anhydride in 0.1 M triethanolamine for 10 min. Slides were then rinsed in 2 × SSC (sodium citrate saline). Hybridization was carried out overnight with sense or antisense cRNA probe (0.3 ng/μl/kb of sequence length). Thirty μ
Immunologic Detection of DIG-labeled Probes
Hybridized DIG-labeled probes were demonstrated either with anti-DIG-labeled alkaline phosphatase (DIG-AP) or with an indirect immunogold system.
DIG-AP Detection of Probes. After washing in TBS (150 mM NaCl in 100 mM Tris-HCl, pH 7.5) for 1 min and incubation in Boehringer–Mannheim blocking solution for 30 min, sections were incubated with a mouse monoclonal anti-digoxigenin antibody (Boehringer–Mannheim) diluted 1:500 in TBS for 1 hr at RT in a humidified chamber. Sections were washed in TBS for 5 min, then in 100 mM Tris-HCl, pH 9.5, 100 mM NaCl, 50 mM MgCl2 for 10 min. Substrate solution for color detection consisted of 100 mM Tris-HCl, pH 9.5, 100 mM NaCl, 50 mM MgCl2, 1 mM levamisole, plus 5 μl NBT/ml and 3.5 μ
Indirect Immunogold Detection of DIG-labeled Probes. Sections were first blocked in Blotto for 5 min and then incubated overnight at 4C with sheep anti-DIG antibody (Boehringer–Mannheim) diluted 1:50 in PBS, 2% nonfat dry milk. Sections were washed three times for 5 min in PBS, incubated with biotinylated rabbit anti-sheep Ig diluted 1:200 for 1 hr at RT, followed by three washes in PBS of 5 min each. Sections were then incubated with 10-nm gold-conjugated goat anti-biotin antibody (EMS Sciences; Worthington, PA) diluted 1:200 in PBS, 1% BSA, 0.01% Tween-20, 0.01% Triton X-100 for 1 hr at RT. Sections were washed in PBS three times for 5 min each and incubated in 1% glutaraldehyde in PBS for 20 min at RT. Sections were then rinsed in PBS once for 5 min followed by washing in dH2O, five times for 1 min each, to remove salts. Sections were then subjected to immunogold–silver staining (IGSS) to amplify the immunogold reaction according to the method described in DeBault and Wang (1994). After IGSS staining, sections were dehydrated and coverslip-mounted with Permount.
Results
POMC mRNA Is Differentially Expressed by Separate Skin Cells at Different Times After Anagen Induction
We had shown by RT-PCR that POMC mRNA is expressed constitutively in murine skin and is upregulated during anagen (Slominski et al. 1996). POMC message was present at telogen, albeit at a low level compared to the steady-state levels observed on Days 3, 5, 10, 15, and 19 after anagen induction. Using those results, we chose telogen and Days 1, 3, 5, and 10 after anagen induction as representative time points in the hair cycle to reveal the cellular sites for POMC mRNA expression using ISH histochemistry and IHC.
ISH histochemistry for POMC mRNA on murine pituitary using an alkaline phosphatase-based immunodetection method of DIG-labeled cRNAs resulted in intense staining in the intermediate lobe, light staining in the anterior lobe, and negative staining of the posterior lobe. This was the expected pattern of staining for the positive control (Figure 1). With this procedure, POMC message was readily detected in the skin at Day 5 after induction of anagen and thereafter. However, no staining was detected with this technique at telogen or Day 1, and only weak staining was detected at Day 3 (data not shown). We therefore used a more sensitive IGSS method to detect the anticipated lower levels of mRNA in these cells while still retaining the advantage of high spatial resolution. The reaction product of the IGSS procedure is deposited as a very fine precipitate in the cytoplasm of the cells, closely resembling the silver grains in an autoradiograph. The density of the precipitate can be related directly to the amount of mRNA present in a cell.
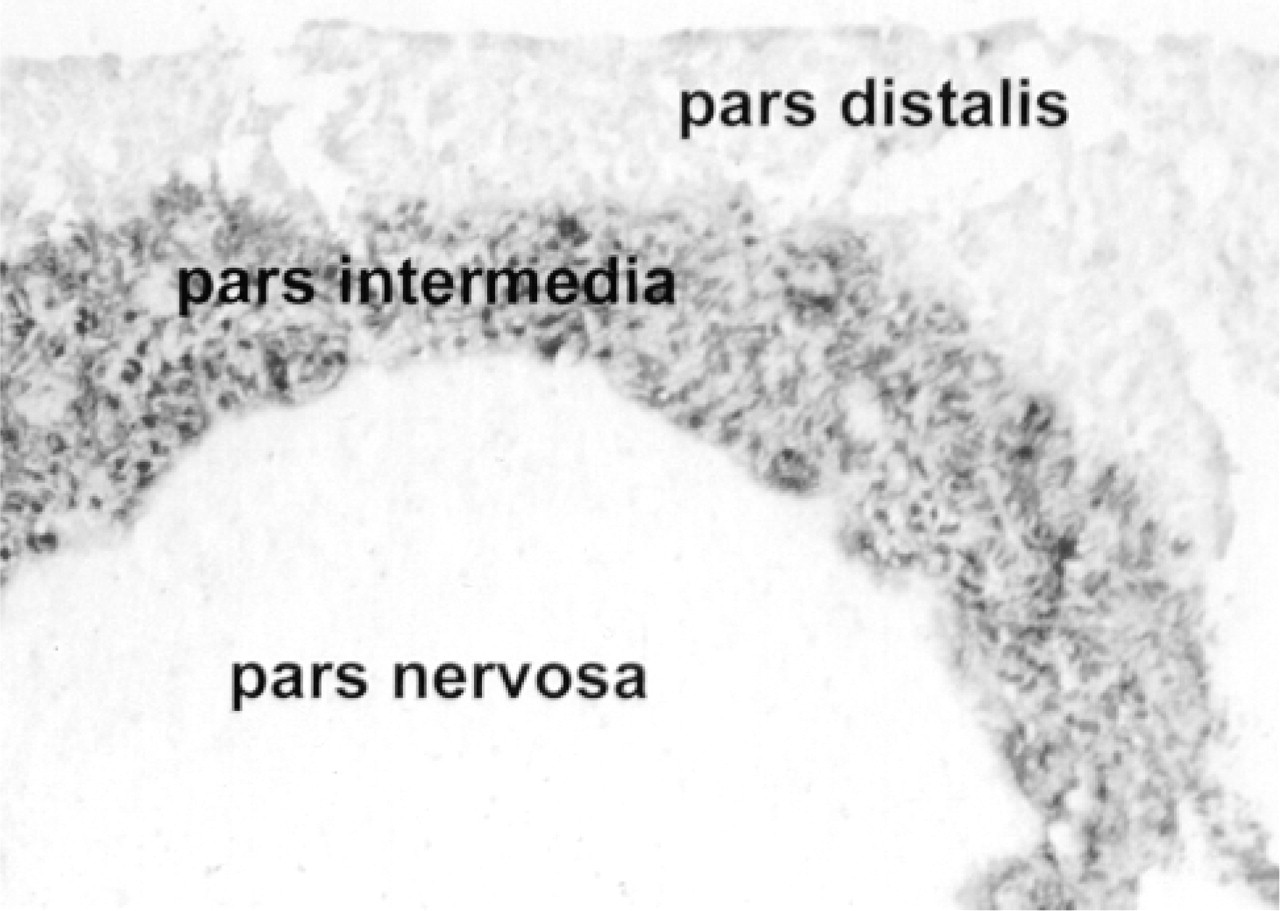
In situ hybridization for POMC mRNA on paraffin sections of mouse pituitary gland demonstrated with anti-digoxigenin labeled with alkaline phosphatase. This is the positive control for specificity of the DIG-labeled antisense probe. The intermediate lobe displays very dense hybridization signal. The anterior lobe is lightly reactive and the posterior lobe is unreactive.
At telogen, POMC mRNA levels were below the level of detection by ISH despite the use of silver enhancement. However, by Day 1 after induction, POMC mRNA was readily detectable in sebaceous units and in scattered keratinocytes in the epidermis. The staining in sebaceous units was very intense at Day 3 and remained so for the rest of the hair growth cycle (Figure 2A and 2C). The number of POMC mRNA-positive epidermal keratinocytes continued to increase throughout anagen, with all epidermal keratinocytes containing signal by Day 10 after induction. The increase in hybridization signal occurred in aggregates of cells in patches in the epidermis, until all were positive (Figure 2A and 2C). Message for POMC was detectable by Day 3 after anagen induction in keratinocytes in the hair follicle, in the region immediately adjacent to the sebaceous gland. Hybridization signal was very sparse in both inner and outer root sheath cells in the follicle below the sebaceous unit at Day 5 (Figure 2B) and increased steadily in cells of both sheaths as the cycle progressed (illustrated for Day 10 in Figure 2D). POMC mRNA was not detected in cells of the hair follicle matrix (Figure 3).
POMC Peptides Displayed a Differential Cellular Localization and Temporal Expression During the Hair Cycle
For IHC, we used antibodies to ACTH, β-endorphin, β-MSH, and α-MSH to demonstrate that the entire POMC precursor is translated. All four of these POMC peptides were present in murine skin. Immunolocalization of ACTH, β-endorphin, and β-MSH is illustrated in Figure 4.
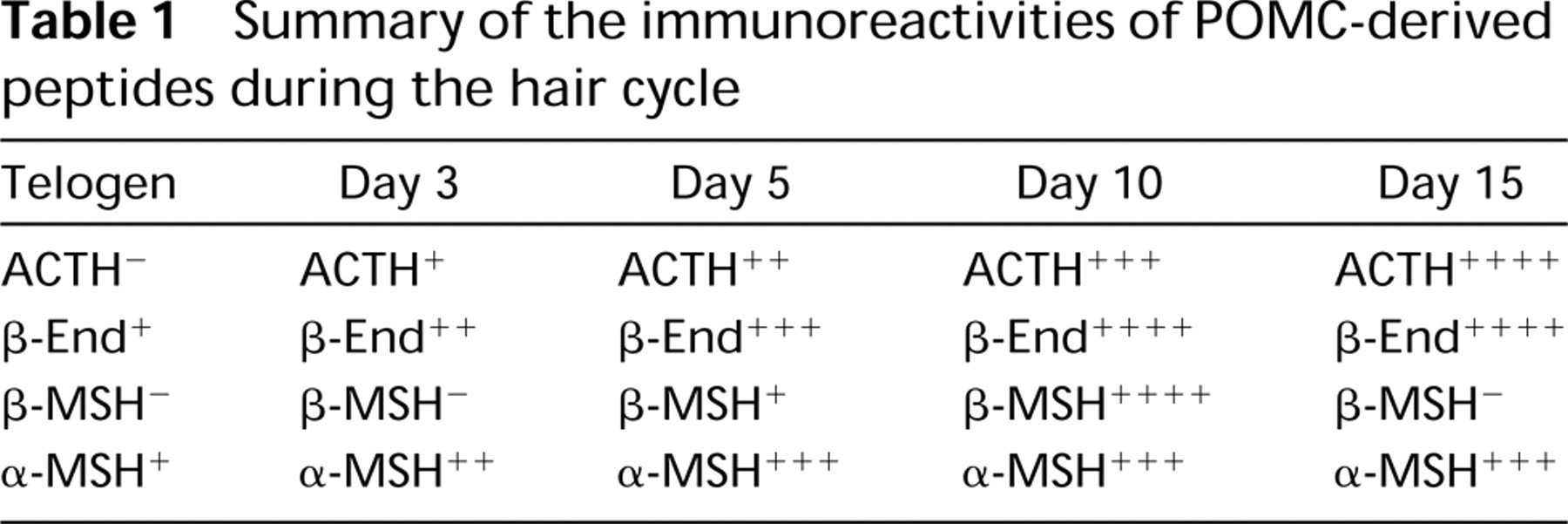
IHC for the POMC products ACTH, β-endorphin, β-MSH, and α-MSH showed a differential cellular distribution and temporal expression during induction of the hair cycle (Table 1). ACTH immunoreactivity was present in epidermal and follicle keratinocytes. Immunoreactivity was absent at telogen, with immunopositivity appearing at anagen Day 3 and increasing to Day 15. In the later part of the cycle, epidermal keratinocytes showed a decrease in ACTH-IR but follicular cell-IR remained constant. In addition to the obvious cellular immunoreactivity for ACTH, there was strong immunoreactivity at the epidermal–dermal junction (Figure 4B and 4C) This staining was apparent at Day 3 and increased in intensity throughout the hair cycle. None of the other peptides displayed the latter immunoreactive pattern.
β-MSH immunoreactivity was present in epidermal and follicle keratinocytes and in sebocytes. β-MSH-IR was absent at telogen. By Day 3, epidermal keratinocytes, some sebocytes, and sebaceous gland duct cells were immunopositive. By Day 5, outer root sheath cells were also positive. The intensity of staining increased dramatically through Day 10 after induction and returned to telogen levels by Day 15.
In contrast to ACTH and β-MSH, β-endorphin staining was limited to sebocytes. It was present at telogen and increased significantly in staining intensity throughout anagen.
At telogen, α-MSH immunoreactivity was absent in keratinocytes and was weakly present in sebocytes. Immunoreactivity was present at Day 3 in epidermal and follicle keratinocytes and increased through Day 6, after which it remained strong through Day 15. Staining of sebocytes did not change and remained at telogen level throughout the hair cycle.
PC1 and PC2 Displayed a Differential Cellular Localization and Temporal Expression During the Hair Cycle
As with POMC mRNA and POMC-derived peptides, PC1 and PC2 also displayed a differential temporal and spatial pattern of expression. PC1 and PC2 were present at very low levels in epidermal keratinocytes in telogen; PC1-IR was greater than PC2-IR (Figure 5). Immunoreactivity for both convertases was increased in anagen VI (Day 15), with PC1- showing the greater elevation. In sebaceous units in telogen, PC1-IR was present at low levels in sebocytes and in adjacent small cells, whereas PC2-IR was at or below detectable levels. In anagen VI, IR for both convertases increased, with PC1-IR again showing the greater increase. Both sebocytes and adjacent small cells showed the increase in PC1-IR, whereas only small cells showed the increase in PC2-IR (Figure 6).
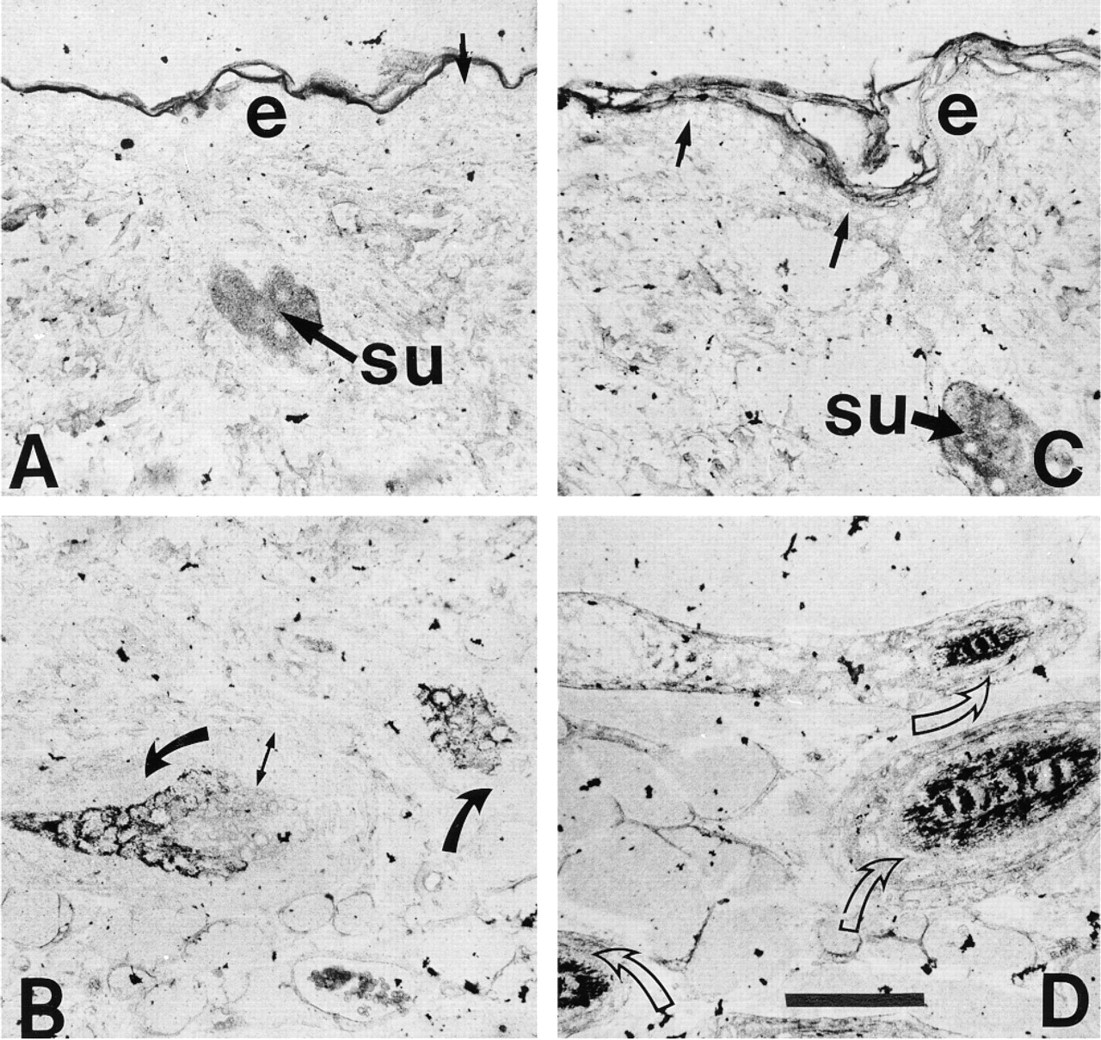
ISH for POMC mRNA in mouse skin on paraffin sections representing Day 5 (
Discussion
In this report we have described the spatial and temporal expression of POMC mRNA, the POMC-derived peptides ACTH, β-endorphin, β-MSH, and α-MSH, and the prohormone converting enzymes PC1 and PC2 during depilation-induced anagen in the murine hair cycle. In anagen, during regeneration of the hair follicle, the expression of POMC mRNA exhibited tissue-specific variations that were consistent with POMC-derived peptide immunoreactivities. Cells that contained POMC peptide IR also expressed POMC mRNA, and where the IR increased there was a parallel increase in mRNA. The levels of PC1 and PC2-IR also showed cell-specific variations and were found in the same cells that expressed POMC peptides. When the cleavage specificities of these convertases are examined (Benjannet et al. 1991; Thomas et al. 1991; Marcinkiewicz et al. 1993) and compared with the spatial and temporal expression of ACTH, β-endorphin, β-MSH, and α-MSH, it can be inferred that PC1 and PC2 activities are responsible for the cell-specific differential processing of POMC in murine skin.
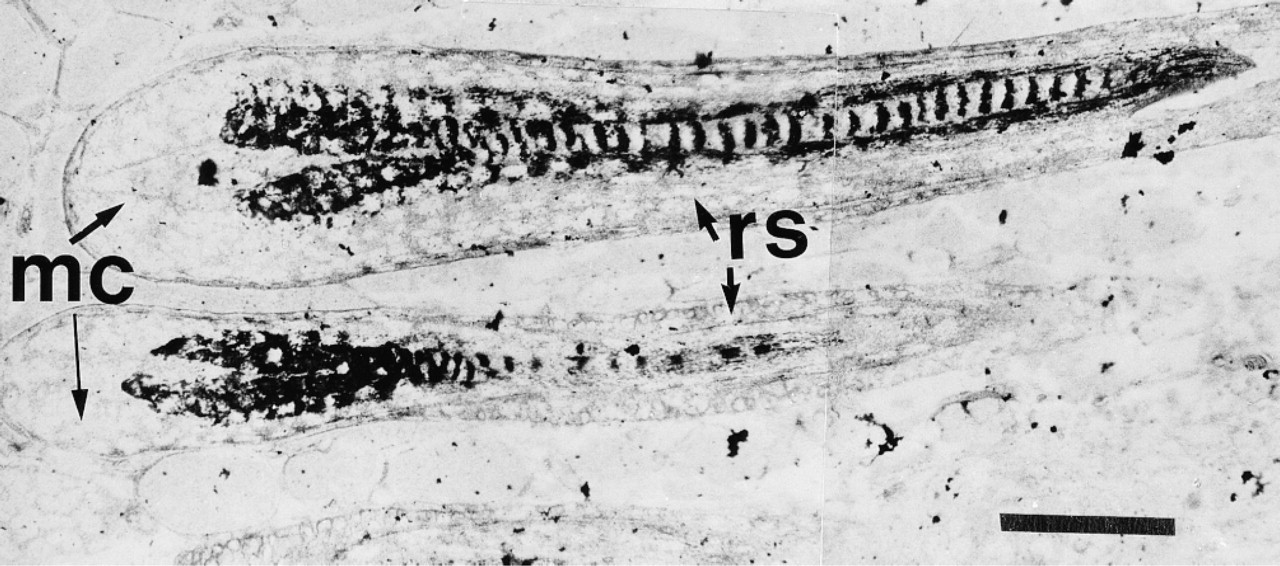
ISH for POMC mRNA in hair follicles on Day 10 after anagen induction with the indirect immunogold system. In this montage that illustrates hair follicles in longitudinal section, keratinocytes in the inner and outer root sheaths (rs) are densely labeled. Note the absence of hybridization in the matrix cells (mc). Bar = 50 μm.
We had previously shown by RT-PCR that POMC mRNA was constitutively expressed throughout the murine hair cycle, with significant upregulation during the growing phase of the cycle (Slominski et al. 1992, 1996). However, by that technique total mRNA was monitored and therefore gene expression in the entire skin, but it did not address cell-specific mRNA expression. The results of the present combined IHC and ISH study refine those earlier observations and identify such cell-specific expression. The increased resolution provided by an immunocytochemically based detection procedure for ISH enabled us to follow the steady increase in POMC message in the different skin tissue compartments. At telogen, POMC mRNA levels were below the level of detection by ISH despite the use of an indirect immunocytochemical detection system and silver enhancement. However, by Day 1 after anagen induction, POMC mRNA was detectable in sebaceous units and remained so for the rest of the hair growth cycle. POMC mRNA levels in keratinocytes in the hair follicle, in the region immediately adjacent to and below the sebaceous gland, were detectable by Day 3 after anagen induction and increased steadily in both inner and outer root sheath cells and in sebaceous gland duct cells. However, even very late in anagen, POMC mRNA was not detected in the matrix of the hair bulb. Epidermal keratinocytes exhibited a slow but steady increase in POMC mRNA levels throughout anagen, starting on Day 3. The lack of detection of POMC mRNA at telogen was similar to the results of previous studies using molecular biology methods, in which the POMC message had been readily detectable in skin in anagen using a Northern blotting technique (Slominski et al. 1992), but it had been necessary to use 26–30 cycles of RT-PCR to detect POMC message in telogen skin (Slominski et al. 1996; Ermak and Slominski 1997). Immunocytochemically based ISH detection procedures can be very sensitive. However, exceedingly low levels of message may still require an autoradiographic technique. Nevertheless, in this study it was possible to follow the differential expression of POMC message in skin throughout anagen.
In other studies, POMC gene transcripts have been detected by Northern blotting analysis in isolated cell lines derived from skin: in cultured human keratinocytes (Slominski et al. 1992; Farooqui et al. 1993; Wintzen and Gilchrest 1996), in the A431 squamous carcinoma cell line, and in stimulated and unstimulated normal keratinocytes (Wintzen et al. 1996). More recently, POMC expression was demonstrated in normal human skin using ISH (Can et al. 1998). However, the state of the hair cycle in these skin samples was not known. The data from the present study and from all of these other reports are in agreement, however, that skin cells can differentially express the POMC gene.
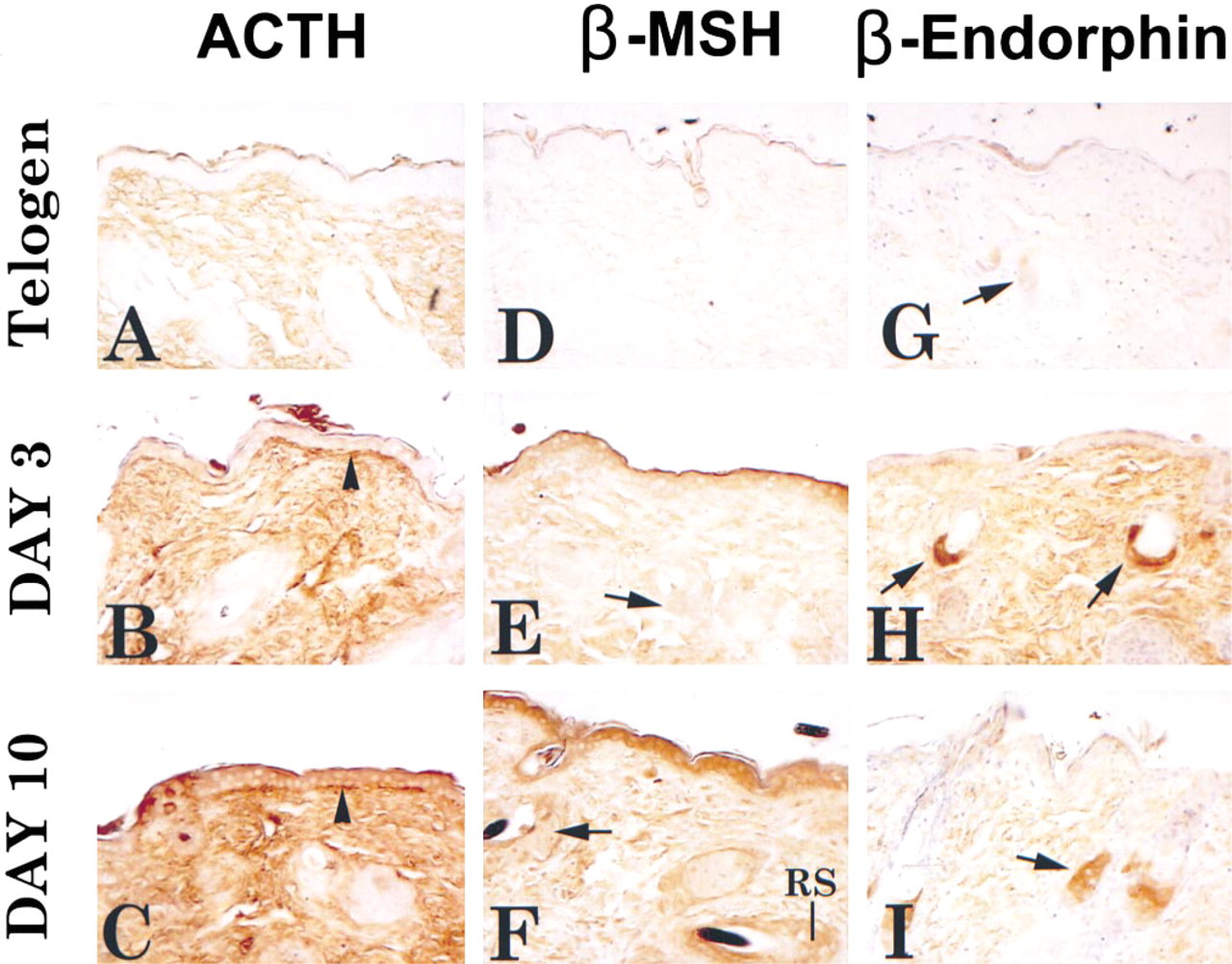
Immunoperoxidase localization of POMC-derived peptides in murine skin at telogen and at Days 3 and 10 after anagen induction. Positive immunoreactivity for ACTH, β-MSH, and β-endorphin demonstrates that the entire POMC precursor is translated and processed. Arrowheads in
Commensurate with the presence of POMC message in these skin compartments was immunoreactivity for one or more POMC-derived neuropeptides (summarized in Table 1). ACTH-IR and β-MSH-IR were both present in epidermal and follicle keratinocytes. However, the appearance of β-MSH-IR preceded that of ACTH-IR in epidermal keratinocytes and decreased to telogen levels sooner than ACTH in follicular keratinocytes. β-Endorphin-IR was not detected in these cells. Immunoreactivities for β-endor-phin, β-MSH, and α-MSH were present in sebaceous units, but ACTH-IR was not. β-Endorphin-IR was restricted to sebaceous units throughout the hair cycle and is consistent with previous reports on β-endorphin-IR in skin (Slominski et al. 1992).
As a first approximation in the identification of the processing enzymes that may be responsible for this differential expression of POMC peptides, our immunocytochemical data show that both PC1-IR and PC2-IR were present in keratinocytes at telogen and that both had increased by mid-anagen. PC1-IR was greater than PC2-IR and increased to a greater extent. In sebaceous units, on the other hand, PC1-IR was detected at low levels in sebocytes and adjacent small cells, whereas PC2-IR was at or below detectable levels. Both sebocytes and small cells showed an increase in PC1-IR, but only the small cells showed an increase in PC2-IR. In analogy with the pituitary, in cells in the skin, the differential expression of PC1 and PC2 may play a role in posttranslational POMC processing seen there (Day et al. 1992; Marcinkiewicz et al. 1993; Zhou et al. 1993).
Summary of the immunoreactivities of POMC-derived peptides during the hair cycle
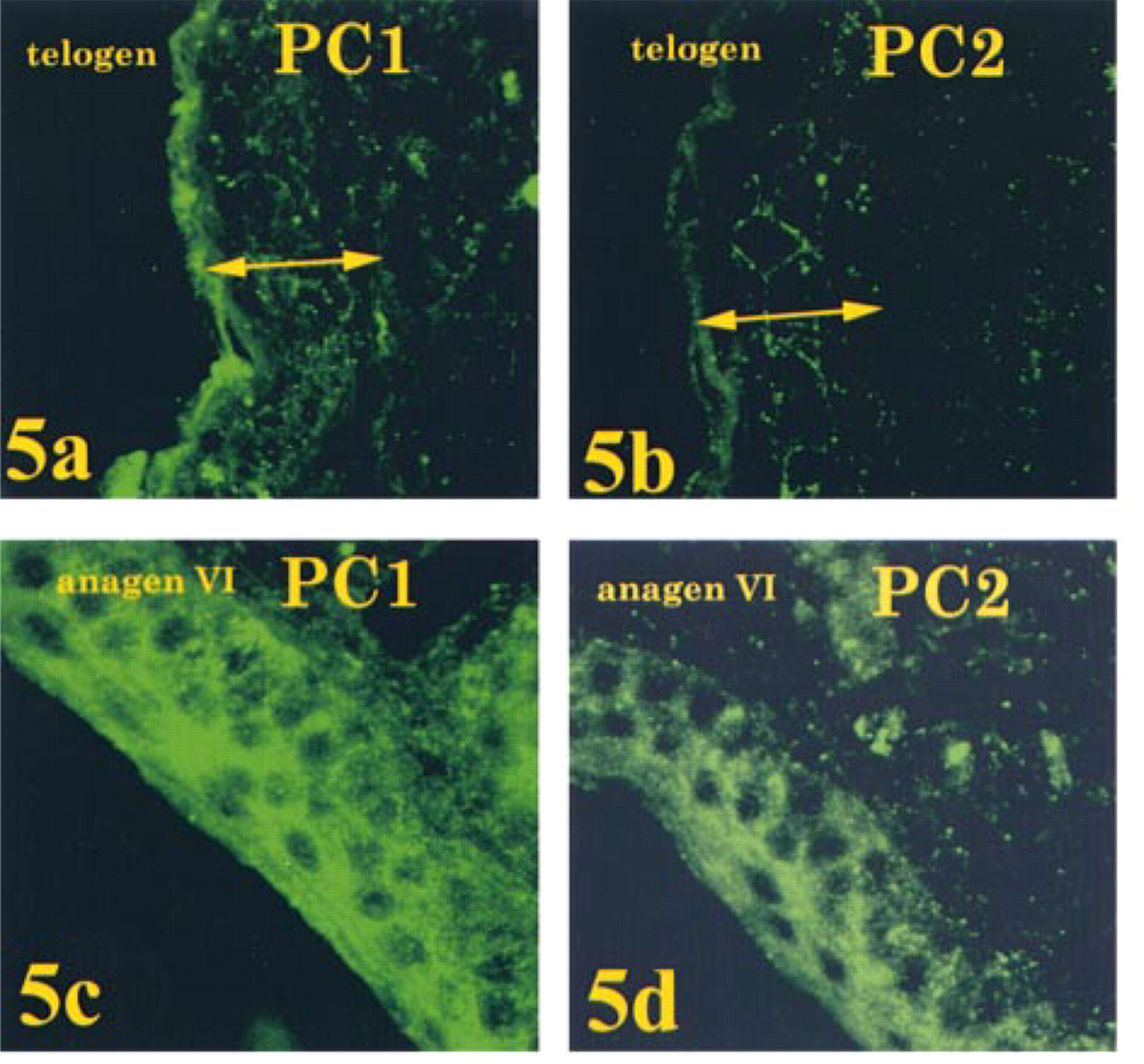
Immunofluorescence localization of PC1 and PC2 in epidermis of telogen skin and anagen VI (Day 15) skin. Maximal intensity projection images of a stack of 15 confocal images (0.3 μm apart). PC1-IR and PC2-IR were present at very low levels in epidermal keratinocytes in telogen; PC1-IR was greater than PC2-IR (
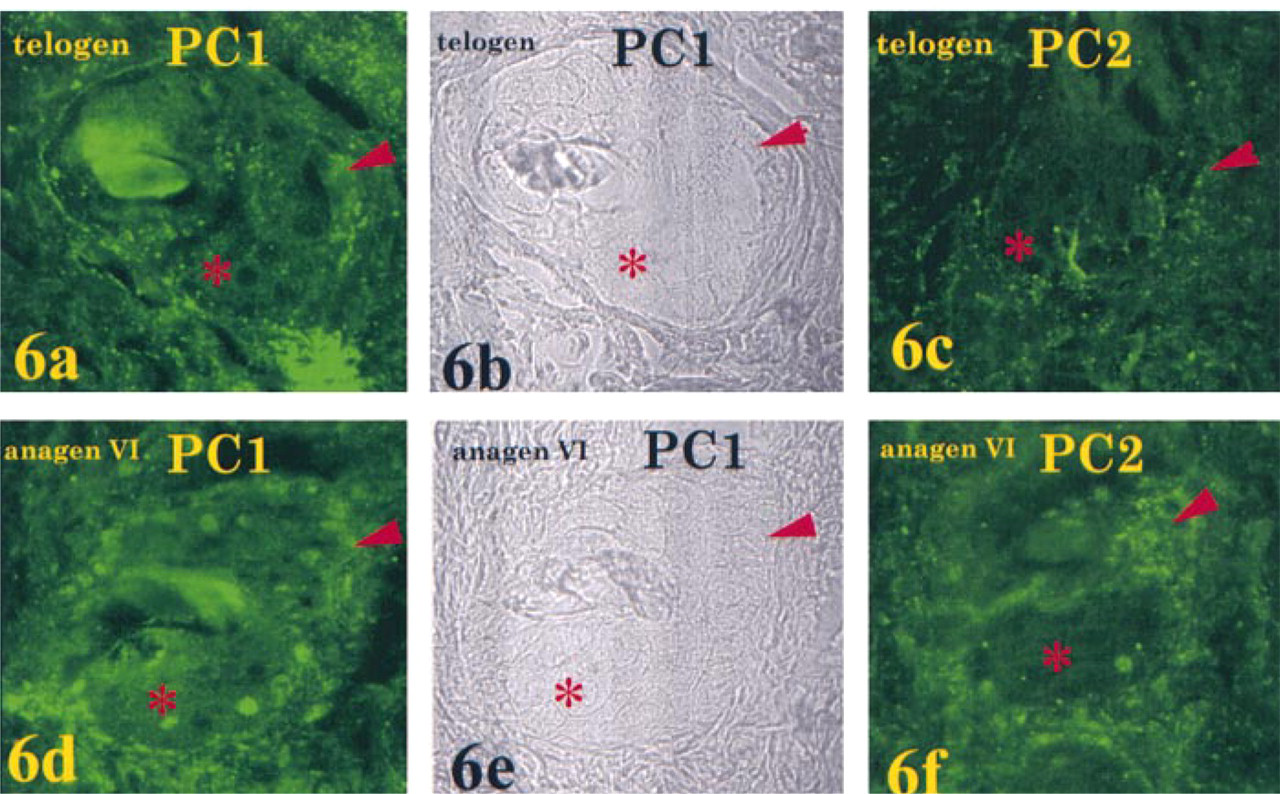
Immunofluorescence localization of PC1 and PC2 in sebaceous units of telogen skin and anagen VI (Day 15) skin. Maximal intensity projection images of a stack of 15–20 confocal images (0.3 μm apart). Upper panels: Telogen. PC1-IR was present at low levels in sebocytes (asterisk) and small cells in the periphery of the sebaceous unit (arrowhead) (
The activity of PC1, alone or in combination with furin, another prohormone convertase that is ubiquitously expressed, could account for the processing of POMC to ACTH, β-endorphin, and β-MSH observed in skin, because (a) PC1 was present in all skin cells at all times examined and (b) PC1-IR increased with progression of anagen. Although the presence of furin has not been demonstrated in skin, it appears to be ubiquitously expressed in all of the cells and tissues examined (Hatsuzawa et al. 1990; Hook et al. 1994). The production of β-endorphin is more efficient with PC2, and PC2 is required for production of α-MSH (Marcinkiewicz et al. 1993). In the present study, β-endorphin-IR, α-MSH-IR, and both PC1-IR and PC2-IR increased in sebaceous units with progression of anagen. Therefore, the increase in PC2 could account for the increase in β-endorphin-IR. Similarly, the dramatic increase in β-MSH-IR, which peaked at mid-anagen and fell back to telogen levels by late anagen, could also have resulted from this increase in PC2.
The prohormone convertases PC1 and PC2 are considered by many to be restricted in their expression to neuroendocrine cells (Seidah et al. 1990, 1991; Smeekens et al. 1991; Muller et al. 1997). However, the present study and that of Wakamatsu et al. (1997) have demonstrated a wider distribution of PC1 and PC2 that is consistent with the demonstration of several peripheral sites for the production of POMC and the many peptides derived from it. In neuroendocrine cells, these PCs function within a regulated secretory pathway, i.e., they are synthesized on the rough endoplasmic reticulum (ER), are trafficked through the Golgi complex, and are sorted into secretory granules along with their substrate in the trans-Golgi Network (TGN). PC1 is activated in the ER and can cleave POMC in the Golgi compartment, and ACTH and β-lipotropin have been shown to be produced before the entry of POMC into secretory granules (Zhou and Mains 1994). Activation of PC2 requires Ca++ and an acidic pH as is found in the TGN and secretory granules (Muller et al. 1997). PC1 and PC2 also appear capable of appropriate cleavage activity and specificity within the constitutive secretory pathway (Smeekens et al. 1992).
Do keratinocytes possess a regulated secretory pathway? The upward migration of keratinocytes in the epidermis from the stratum basal through the stratum spinosum and stratum granulosum to the stratum corneum is accompanied by differentiation of the keratinocyte into a secretory granular cell. These cells are responsible for the biogenesis of lamellar bodies (LBs) or lamellar granules, the contents of which, when secreted, are responsible for the formation of the epidermal permeability barrier (Madison et al. 1998). These LBs contain lipids and lysosomal enzymes and, like other secretory granules, bud from the TGN, and on fusion with the plasma membrane release their contents by exocytosis (Elias et al. 1998). Cells in both the spinosum and granulosum contain LBs, where they can comprise up to 10% of the cytosolic volume (Menon et al. 1992; Elias et al. 1998). The lamellar body has been called a “dual-function” organelle (i.e., part secretory and part lysosomal) because of its secretory function and its lysosomal enzyme content (Madison et al. 1998). This organelle may, in fact, be an example of the “sorting by retention” hypothesis in the regulated secretory pathway (reviewed in Arvan and Castle 1998), in which regulated secretory proteins are sorted by retention in a post-Golgi immature secretory granule (ISG). According to this hypothesis, non-secretory granule proteins or remnants of processing of regulated secretory proteins are removed by constitutive-like vesicle budding from the ISG, resulting in the formation of mature secretory granules. In particular, lysosomal enzymes and mannose-6-phosphate receptors have been found to follow this pathway, being sorted away from insulin in pancreatic β-cells (Kuliawat et al. 1997). Perhaps, in this light, the keratinocyte can be construed to possess a regulated secretory pathway. Others have suggested that the keratinocyte should be considered a specialized secretory cell (Madison et al. 1998). Whether or not POMC-derived peptides travel within the LB secretory pathway or in a similar but parallel pathway remains to be investigated, and studies are under way in our laboratory to address this issue.
In summary, we have demonstrated hair cycle-associated changes in spatial and temporal expression of the convertases PC1 and PC2 and of ACTH, α-MSH, β-MSH, and β-endorphin peptides in murine skin, and we suggest that they are the result of both POMC mRNA translation and cleavage specificities of PC1 and PC2 enzymes.
Footnotes
Acknowledgements
Acknowledgments
Supported in part by grants from the NSF: IBN 9405242 and IBN 9604354 to AS and JEM, and from the NIH: S10RR12894 to JEM.
We thank Drs B.A. Eipper (Johns Hopkins University School of Medicine), R.G. Allen (Oregon Health Sciences Center), and S.H. Pomerantz (University of Maryland) for gifts of antisera, Drs G. Davila and J. Pawelek (Yale University) for gifts of POMC peptides, and Dr J.L. Roberts (Mount Sinai Medical Center) for the mouse POMC cDNA. We acknowledge the Imaging Core Facility at the Albany Medical College (J.E. Mazurkiewicz, director) for use of the confocal microscope. We also thank Dr G. Ermak (University of Southern California) for expert technical assistance and discussion in aspects of the molecular biology.